
微咸水灌溉下冬小麦光合作用与光响应曲线模拟
2021-12-07 05:36庞桂斌张立志潘维艳徐征和徐俊增
农业机械学报 2021年11期
庞桂斌 张立志 丛 鑫 潘维艳 徐征和,4 徐俊增
(1.济南大学水利与环境学院, 济南 250022; 2.山东省地下水数值模拟与污染控制工程技术研究中心, 济南 250022;3.山东省水利科学研究院, 济南 250014; 4.山东省生态固碳与捕集利用工程技术研究中心, 济南 250022;5.河海大学农业科学与工程学院, 南京 210098)
0 引言
开发利用微咸水资源,对于缓解淡水资源短缺、扩大农业水源、抗旱增产有着极其重要的作用。微咸水用于灌溉,在提供作物生长所需水分的同时,也给土壤带入大量盐分[1-2]。目前的研究主要关注微咸水灌溉方法及其水土环境效应、作物影响等方面,包括灌溉技术[3-5]、灌溉制度[6-7]、灌溉水质[8]、土壤水盐运移[9-11]、土壤理化性质[12-13]、地下水环境[14]、作物生长和产量品质[15-17]等,积累了较多的经验,而微咸水灌溉对不同作物生长影响的内在机理及作物遭受盐分胁迫后的调节机制仍需进一步研究。微咸水灌溉导致土壤含盐量增加,势必会影响土壤溶液的渗透势,从而会改变土壤水分的有效性,可能会引发生理干旱,同时土壤-植物根系-植物叶片水势梯度的变化,会诱导叶片细胞膨压的改变,导致气孔导度、蒸腾作用和光合特性等生理指标的调节与适应,从而对光合产物积累及其在根、茎、叶和籽粒间的分配产生显著影响[18-20],因此微咸水灌溉下的作物生理响应规律及其适应机制有待进一步深入研究。
光合作用是作物生长发育的基础和生产力的决定性因素,是作物育种、栽培以及作物应对环境胁迫领域研究的重要指标。为了定量研究作物光合速率对光照强度的响应特征,国内外学者通过建立光响应模型,例如直角双曲线模型、非直角双曲线模型、指数模型、直角双曲线修正模型等[21-22],模拟得出反映作物光合生理生态过程的基础参数,学者们还对不同水肥调控、逆境胁迫下的多种作物进行光响应曲线模型的比较研究[23],但针对冬小麦在微咸水灌溉条件下的光响应特征相关研究报道相对较少。以往的研究表明,修正的直角双曲线模型克服了其他几类模型无法直接获得植物的最大净光合速率和饱和光照强度等指标的缺陷,可以直接求得最大净光合速率、光补偿点和饱和光照强度等参数,而且其参数拟合效果较好[24]。本文以黄河三角洲地区的主要灌溉粮食作物冬小麦作为研究对象,选择直角双曲线修正模型[25],研究微咸水灌溉条件下抽穗期和开花期冬小麦旗叶的光合作用及光响应特征,揭示微咸水灌溉下冬小麦的光合生理响应规律与调节机制,为该类地区制定较为合理的微咸水灌溉策略提供科学依据。
1 材料与方法
1.1 试验区概况
试验区位于山东省滨州市沾化区下洼镇张王二村(图1a),地处韩墩引黄灌区的梢段,北纬37°34′,东经117°45′,属于暖温带季风气候区,大陆性气候特征明显,四季差别显著。年平均日照时数为2 690.3 h,年平均气温12℃,年平均降水量575.5 mm,年内降水量季节分配不均匀,年均蒸降比为3.22,易造成地下水上升,土壤返盐,形成土壤盐碱化。试验区地下水位埋深在2~3 m,浅层地下水矿化度为5~10 g/L。试验区0~40 cm土壤平均含盐量为2.11 g/kg,按照土壤盐渍化分布等级[26],属于中度盐碱地。根据土壤粒径分析,0~20 cm为壤土,20~40 cm为砂质壤土,40~60 cm为砂质壤土,60~80 cm为壤质砂土,80~100 cm为壤土,土壤理化性质见表1。2015—2016年小麦生育期内降雨总量为139.1 mm,2016—2017年小麦生育期内降雨总量为117.5 mm,如图2所示。
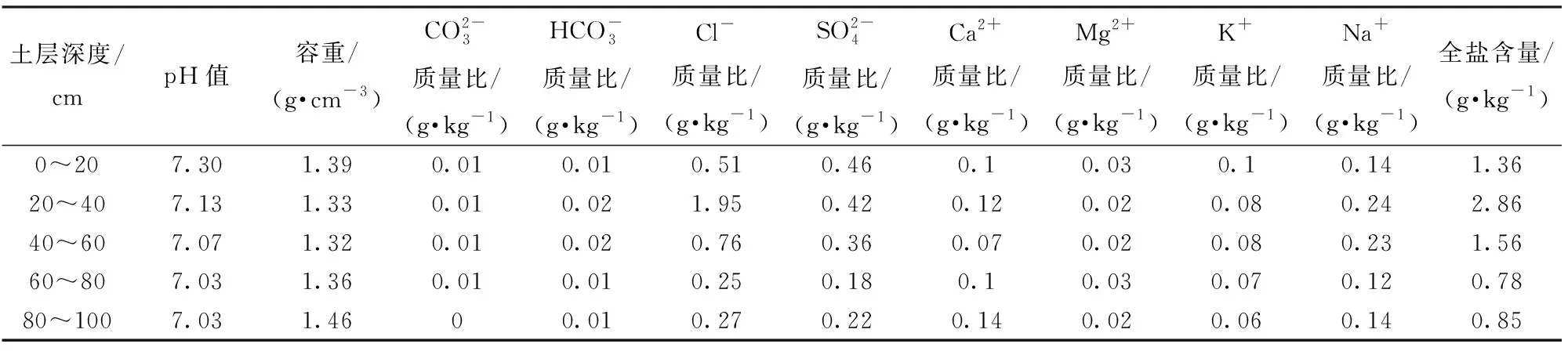
表1 试验区土壤理化性质Tab.1 Soil physical and chemical properties of experimental area
1.2 试验设计
设置 2 种灌溉处理(微咸水和淡水处理(对照处理)),每个处理设3次重复,共6个试验小区,小区面积为18 m2(6 m×3 m)(图1b)。为避免侧渗干扰,小区之间设置0.5 m隔离带和1.5 m深塑料薄膜。供试作物为冬小麦,2015—2016年生长季播种日期为2015年10月8日,次年6月13日收获,生育期为249 d,2016—2017年生长季播种日期为2016年10月8日,次年6月12日收获,生育期为248 d。其他措施与常规大田生产相同,并保持所有处理一致。
微咸水灌溉所用水源来自试验区浅层地下水,经咸水淡化装置淡化处理后的矿化度低至0.20~0.23 g/L,直接作为淡水水源。将处理后的淡水和抽取的地下咸水储存在蓄水池进行充分混合,将矿化度调至3 g/L,作为微咸水灌溉水源。根据以往研究发现,冬小麦幼苗期对盐分比较敏感[27],因此在灌溉试验设计时返青-拔节期间使用淡水。中后期采用了拔节-抽穗期和抽穗-灌浆期灌溉微咸水的组合方案,2016年分别于3月11日、4月21日和5月15日进行灌溉,2017年分别于3月15日、4月23日和5月12日进行灌溉。具体灌水方案见表2。

表2 冬小麦微咸水灌溉方案Tab.2 Brackish water irrigation scheme of winter wheat mm
1.3 观测内容与方法
土壤含盐量:采用对角线(三点法)取样,在返青后各个生育期和灌溉后采用取土钻分别采集0~20 cm、20~40 cm、40~60 cm、60~80 cm、80~100 cm层土壤,参考《土壤农业化学分析方法》,采用水土质量比为5∶1的土壤浸提液滴定法测定各盐分离子含量,并计算出相应的土壤全盐含量。
光合特性:采用英国ADC公司生产的LCpro-SD型便携式光合作用仪,测量冬小麦叶片净光合速率(Pn,μmol/(m2·s))、蒸腾速率(Tr,mmol/(m2·s))、气孔导度(Gs,mol/(m2·s))、胞间CO2浓度(Ci,μmol/mol),大气CO2浓度(Ca,μmol/mol)等生理指标。在冬小麦抽穗期、开花期、灌浆期和乳熟期选择典型晴好天气,沿小区对角线选择3片旗叶,于08:00—16:00每隔2 h进行测定。
光响应特征曲线:在拔节期微咸水灌溉后的抽穗期和开花期,选择典型晴好天气,在观测日的10:00—12:00期间,采用LCpro-SD便携式光合作用仪,测定冬小麦旗叶的光响应特征。测定时利用人工光源自动控制光合有效辐射通量密度,分别设定为2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、200、100、50、0 μmol/(m2·s)(共13个梯度),每个光照强度梯度下测定时间为120 s。光响应特征曲线模拟采用直角双曲线修正模型,模型表达式为
(1)
式中Pn(I)——净光合速率函数
α、β、γ——系数
I——光合有效辐射,μmol/(m2·s)
Rd——植物的暗呼吸速率,μmol/(m2·s)
模型特征参数的计算方法参照文献[24]。
气孔限制值[28](Ls,%)计算公式为
Ls=(Ca-Ci)/Ca×100%
(2)
1.4 数据分析
统计分析和显著性检验采用IBM SPSS Statistics 22.0完成,非线性模型模拟参数求解采用1stopt 1.5专业版完成,图表绘制采用Microsoft Excel 2013完成。
2 结果与分析
2.1 微咸水灌溉对土壤盐分的影响
返青期两个处理采用淡水灌溉,土壤盐分经水分淋洗向下运移,盐分下降明显。至拔节期灌溉前,土壤含盐量变化较小,返盐不明显。拔节期灌溉后,淡水处理土壤含盐量进一步下降,微咸水处理由于盐分进入土壤,土壤表层含盐量增加。由图3可知,在2016、2017年0~20 cm土壤含盐量分别增加0.8、0.49 g/kg;20~40 cm土壤含盐量分别增加0.92、0.05 g/kg。至冬小麦灌浆期灌溉前,气温升高,土壤水分蒸发加强,各处理均出现不同程度的返盐。灌浆期灌溉后,两年的淡水处理土壤浅层(0~40 cm)土壤全盐含量分别下降至2.25、2.28 g/kg,微咸水灌溉处理土壤浅层(0~40 cm)全盐含量分别上升了1.68、2.67 g/kg。从整个生育期来看,与淡水处理相比,2016、2017年微咸水灌溉导致浅层(0~40 cm)土壤含盐量显著增加了37.8%和64.3%,而深层(40~60 cm)土壤含盐量分别增加了9.1%和减少了8.3%,变化不显著(p>0.05)。此外,该区域自4月下旬开始,降雨逐渐增多,可以对盐分进行淋洗,降低土壤含盐量,减轻微咸水灌溉对作物生长的影响以及下茬轮作种植的影响。
2.2 微咸水灌溉对冬小麦蒸腾速率的影响
选取拔节期灌溉后典型日08:00—16:00期间5个时刻测定的冬小麦叶片蒸腾速率,分析微咸水灌溉对冬小麦叶片蒸腾速率日变化的影响(图4)。
由图4可以看出,冬小麦叶片蒸腾速率日变化呈现出“单峰型”(2017年5月12日)和“双峰型”(2016年5月1日、2016年5月10日、2017年5月1日)的变化规律,“单峰型”在12:00左右达到峰值,由于当天午间为多云天气,气孔蒸腾在午间仍然保持在较高水平。 “双峰型”在10:00左右达到第1个峰值,在12:00左右有所下降,到14:00左右达到第2个峰值,这是由于在光照较为强烈的午间,作物为了控制自身失水过多关闭了部分气孔,导致蒸腾速率的下降。
不同灌溉处理的差异明显,微咸水灌溉处理冬小麦叶片蒸腾速率日均值均低于淡水处理,2016年和2017年抽穗期分别降低了19.1%和31.4%,处理间差异显著(p<0.05),而2016年和2017年开花期分别降低了11.6%和11.0%,处理间差异并不显著。由2.1节可知,拔节期微咸水处理灌后0~40 cm表层土壤含盐量较灌溉前增加了73.3%(图3),微咸水灌溉使得土壤含盐量上升,土壤溶液的渗透势随之增长,降低了土壤水分的有效性,土壤-根系-叶片的水势梯度减小,引起部分气孔关闭,冬小麦叶片的蒸腾速率下降。但是随着时间推移,微咸水灌溉对作物蒸腾速率的负面影响在降低,这与作物自身对盐分的适应调节机制及土壤水分等因素有关。
2.3 微咸水灌溉对冬小麦净光合速率的影响
分析冬小麦叶片净光合速率的日变化规律(图5)可以看出,日变化趋势不尽相同,有的呈现“单峰型”曲线变化,有的呈现“双峰型”曲线变化,还有的呈现“下降型”变化趋势,这是由于光合速率受作物自身、土壤水盐以及气候环境等因素的综合影响所致。但是可以看出,一天当中净光合速率的最高值出现在午前,而作物午间受到强光照射胁迫时出现了“光合午休”的现象,这与前人的研究结果一致[29]。
比较两种灌溉处理的净光合速率的日变化差异可以看出,在2016年的抽穗期和开花期,12:00之前,微咸水灌溉处理的净光合速率较淡水处理降低了9.7%和7.7%,在12:00之后,微咸水灌溉处理的净光合速率出现反弹,而微咸水灌溉处理净光合速率日均值仅降低了0.8%和0.9%。2017年抽穗期和开花期,两种灌溉处理也表现出相似的变化规律,只是微咸水灌溉处理高于淡水处理的时间提前到10:00,计算净光合速率日均值可以发现,微咸水灌溉处理较淡水处理增加了8.3%和3.5%,处理间差异并不显著(p>0.05),这也表明微咸水灌溉并没有显著降低冬小麦抽穗开花期叶片净光合速率,反而提高了叶片午后的光合效率,可见盐分处理的作物在高温和强光下具有更高的光合速率[19]。
2.4 微咸水灌溉对冬小麦气孔限制值的影响
冬小麦叶片光合作用主要受气孔因素和非气孔因素影响,在不同的条件下占主导地位的因素不同[30]。分析不同灌溉处理下的冬小麦气孔限制值变化规律(图6)可知,抽穗期(2016年5月1日、2017年5月1日)和开花期(2016年5月10日、2017年5月12日),微咸水灌溉处理叶片气孔限制值在午前部分时段(2016年5月1日、2017年5月12日)出现了低于淡水处理的现象,有部分时段(2016年5月10日、2017年5月1日)出现了高于对照的现象,而午后均表现为高于淡水处理的趋势,不同灌溉处理之间的对比差异与光合速率变化规律类似。这表明了微咸水灌溉处理午前叶片净光合速率的下降受到气孔因素和非气孔因素的共同影响,而午后净光合速率的增加均伴随着较高的气孔限制值,可能是由于适度的盐分胁迫反而促进了根系下扎,延缓了叶片衰老,改善了叶肉细胞光合活性和叶片的光合能力,提高了胞间CO2的利用,使胞间CO2浓度降低,从而减少了非气孔因素对光合作用的限制。
2.5 微咸水灌溉下冬小麦光响应曲线模拟
由表3可以看出,光响应曲线模拟的相关系数达到0.997~0.999,说明了直角双曲线修正模型可以较好地模拟冬小麦叶片净光合速率对于光合有效辐射的响应关系。
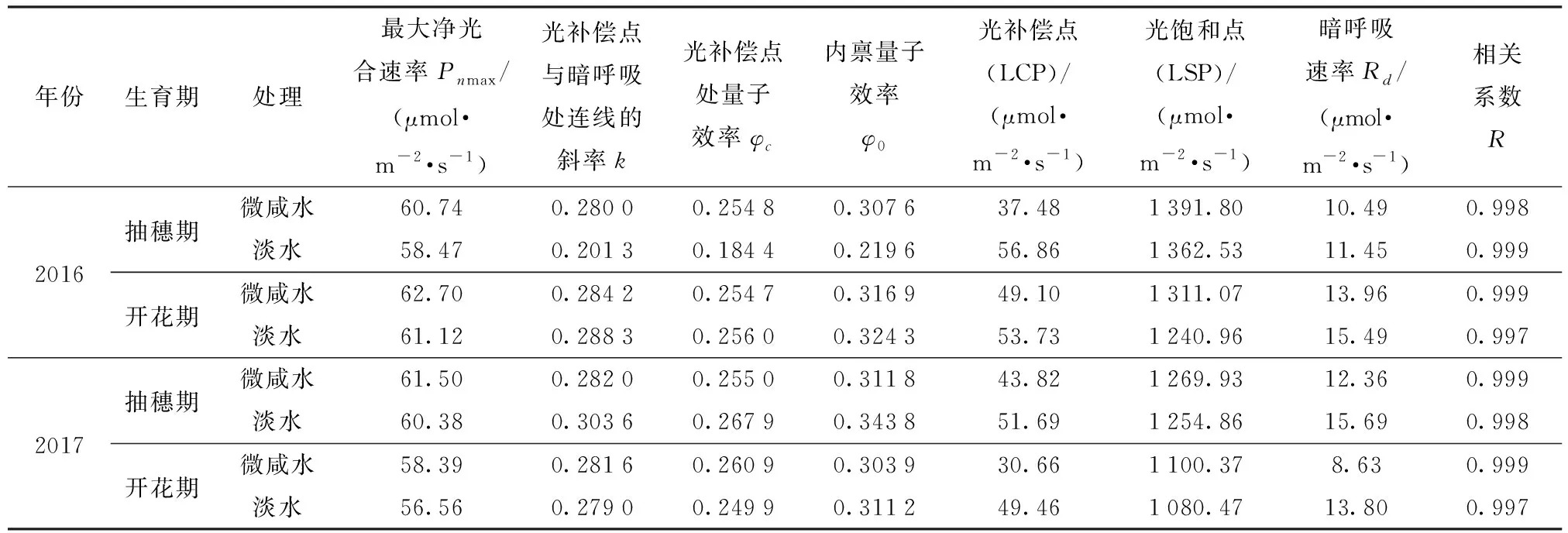
表3 不同灌溉处理冬小麦抽穗开花期光响应曲线特征参数Tab.3 Winter wheat leaf photosynthetic light response characteristics parameters at heading and flowering stages under different irrigation treatments
与淡水处理相比,2016年和2017年微咸水处理冬小麦抽穗期最大净光合速率Pnmax增加了2.27、1.12 μmol/(m2·s),光饱和点(LSP)增加了29.27、15.07 μmol/(m2·s),光补偿点(LCP)降低了19.38、7.87 μmol/(m2·s);冬小麦开花期Pnmax增加了1.58、1.83 μmol/(m2·s),LSP增加了70.11、19.9 μmol/(m2·s),LCP降低了4.63、18.8 μmol/(m2·s)。说明微咸水灌溉改善了冬小麦的光合作用能力,增强了冬小麦对于强光和高温的适应能力,同时利用弱光的能力也有所提升。从不同灌溉处理内禀量子效率φ0和光补偿点处量子效率φc的对比也可以发现,抽穗期微咸水处理均高于淡水处理,而到开花期两个处理几乎一致,可见微咸水灌溉并未抑制冬小麦把光能转化为净能量的能力和利用光能的潜能,部分生育阶段甚至优于对照。同时,与淡水处理相比,2016年和2017年微咸水处理冬小麦抽穗期的暗呼吸速率Rd分别降低了0.96、3.33 μmol/(m2·s),微咸水处理冬小麦开花期Rd分别降低了1.53、5.17 μmol/(m2·s),较低的暗呼吸速率可以保证较快的干物质累积,一定程度促发了冬小麦利用光能的潜力。
光响应曲线反映植物光照强度与光合速率间的变化规律,是评价植物光合能力的重要手段[31]。如图7所示,在低光照强度(0~200 μmol/(m2·s))下,无论是淡水处理还是微咸水处理,冬小麦叶片净光合速率均随着光照强度的增加而近乎直线上升,而且微咸水处理的上升速率略快于淡水处理;在中光照强度(200~800 μmol/(m2·s))下,不同处理的净光合速率的增加变缓;在高光照强度(>800 μmol/(m2·s))下,冬小麦叶片净光合速率达到最大值,然后随着光照强度的增加净光合速率趋于稳定,甚至略有下降的趋势。但是在不同的光照梯度下,无论是抽穗期还是开花期,微咸水处理的净光合速率均呈现略高于淡水处理的现象,这也进一步验证了2.3节中自然光照条件下,微咸水灌溉并未明显抑制冬小麦净光合速率的研究结果。
3 讨论
一般认为,灌溉水中的盐分会对作物产生毒害,如降低膨压、减小细胞扩张速度、破坏叶绿体等,从而导致生长速度和光合作用降低,最终对干物质积累和产量产生不良影响[32]。张余良等[33]研究了微咸水灌溉对小麦生理特性及产量的影响,在灌水量900 m3/hm2条件下,与淡水相比,矿化度3.5 g/L微咸水灌溉时,蒸腾速率和气孔导度分别下降了18.8%和22.5%,光合速率却没有降低,而灌入矿化度4.5 g/L微咸水时,光合速率下降了19.7%。史海滨等[19]对不同盐分处理对向日葵光合速率的影响研究表明,随着灌溉水盐分含量的增加,根系吸收水分的能力降低,叶片的蒸腾速率随之下降,同时12:00之前受到气孔限制的光合速率降低,而12:00后的光合速率和气孔限制值高于低盐处理。微咸水灌溉后土壤含盐量增加,土壤溶液的渗透势随之增长,降低了土壤水分的有效性,土壤-根系-叶片的水势梯度减小,引起部分气孔关闭,导致了叶片气孔导度和蒸腾速率下降[33]。本研究在微咸水灌溉冬小麦的试验中也发现了类似的规律。作物光合速率的变化受到气孔因素和非气孔因素的影响,本研究表明,微咸水灌溉降低了冬小麦12:00之前光合速率,抽穗期和开花期光合速率的下降主要是受到气孔因素和非气孔因素的共同影响,12:00之后微咸水处理冬小麦光合速率出现高于对照的情况,同时也伴随着较高的气孔限制值,主要是由于非气孔因素的改善所致,具体的非气孔因素还有待进一步研究。
郭丽等[31]通过长期的微咸水灌溉试验发现,矿化度控制在1.8 g/L咸淡混灌和3.6 g/L咸水与淡水轮灌处理并未导致产量及光合作用下降,此外,采用非直角双曲线方程对光响应曲线进行拟合得出,最大净光合速率、表观光量子效率、光饱和点、光补偿点和暗呼吸效率均没有显著差异。本研究表明,无论是抽穗期还是开花期,在人工光源不同的光照强度梯度下,微咸水处理的净光合速率略高于淡水处理,同时,通过直角双曲线修正模型拟合可得,光补偿点和暗呼吸速率略有降低,但是最大净光合速率、表观光量子效率和光饱和点却呈现出超过淡水处理的现象,这与郭丽等[31]的研究结果相类似但有所差异,说明3 g/L微咸水灌溉改善了抽穗开花期冬小麦的光合作用能力,增强了冬小麦对于强光和高温的适应能力,同时利用弱光的能力也有所提升,表现出更高的光合效率。这可能是由于试验田初始含盐量并不高,短时间内微咸水带入土壤的盐分含量较少,不足以对作物生长和光合作用造成明显的影响。一般利用微咸水灌溉的前1~3年,可能出现比淡水灌溉更有利于作物生长的现象,随着微咸水灌溉年限延长,土壤盐分逐步增加,对作物生长影响的负面作用逐步显现[34]。此外,采用不同光响应曲线模型进行特征参数拟合,也可能会表现出不一样的变化规律,相关的研究结论需进一步的试验验证。
4 结论
(1)微咸水灌溉将盐分带入土壤,导致土壤浅层(0~40 cm)的含盐量增加,但对深层(60~100 cm)土壤含盐量的影响较小,对作物生长产生一定的影响。抽穗期和开花期,淡水处理(矿化度0 g/L)和微咸水处理(矿化度3 g/L)冬小麦叶片蒸腾速率日变化呈现出“单峰型”和“双峰型”的变化规律,微咸水灌溉后土壤含盐量增加,降低了土壤水分的有效性,引起部分气孔关闭,导致了叶片蒸腾速率下降;冬小麦光合速率及气孔限制值日变化规律表明,与淡水处理相比,微咸水灌溉降低了冬小麦午前光合速率,光合速率的下降主要是受到气孔因素和非气孔因素的共同影响,午后微咸水处理冬小麦光合速率的增加却伴随着较高的气孔限制值,主要是由于非气孔因素的改善所致。
(2)在人工光源不同的光照强度梯度下,无论抽穗期还是开花期,微咸水处理冬小麦叶片净光合速率均超过了淡水处理,引入直角双曲线修正模型对光合特征参数进行拟合得出,2016年和2017年微咸水处理冬小麦抽穗期Pnmax增加2.27、1.12 μmol/(m2·s),LSP增加29.27、15.07 μmol/(m2·s),LCP降低了19.38、7.87 μmol/(m2·s),Rd降低了0.96、3.33 μmol/(m2·s);2016年和2017年冬小麦开花期Pnmax增加了1.58、1.83 μmol/(m2·s),LSP增加了70.11、19.9 μmol/(m2·s),LCP降低了4.63、18.8 μmol/(m2·s),Rd降低了1.53、5.17 μmol/(m2·s)。说明微咸水处理较淡水处理表现出更高的光合效率,不但增强了抽穗开花期冬小麦对于强光和高温的适应能力,同时也提升了冬小麦利用弱光的能力,一定程度促进了冬小麦利用光能的潜力。
猜你喜欢
中国农学通报(2022年29期)2022-11-25
农业工程学报(2022年4期)2022-04-24
农业与技术(2020年12期)2020-07-04
山东工业技术(2018年13期)2018-08-20
现代农业科技(2018年22期)2018-01-15
新课程·下旬(2017年7期)2017-08-14
安徽农业科学(2017年3期)2017-08-13
绿色科技(2017年7期)2017-05-12
江苏农业科学(2016年12期)2017-04-05
红领巾·成长(2016年1期)2016-09-10
