
营养代偿效应对育肥猪胴体品质、肉品质、抗氧化能力及肌内脂肪代谢的影响
2021-12-05 07:19穆淑琴李千军
家畜生态学报 2021年11期
闫 峻,刘 哲,罗 强,李 宁,穆淑琴,李千军*
(1.天津市农业科学院 畜牧兽医研究所,天津 300381;2.天津大学 化工学院,天津 300354;3.天津农学院 动物科学与动物医学学院,天津 300384;4.西北农林科技大学 动物科技学院,陕西 杨凌 712100)
营养代偿效应是指动物经过一段时间的营养限制后,去除限制因素后动物加速生长的一种现象,限饲则是进行营养限制的普遍手段[1]。在营养代偿效应的研究中表明,代偿生长可以影响牛肌肉的嫩度,在代偿生长过程中牛脂肪酸代谢也受到调节,从而影响牛肌内脂肪组成[2]。研究表明,强度适宜的限饲可以改善动物体的健康状态,通过抗氧化酶系统增加动物的抗氧化能力,激活机体对能源物质的利用,改变能量代谢的方式[3];但高强度的限饲,则会导致动物产生饥饿应激,机体内消耗大量脂肪产生不饱和脂肪酸,不饱和脂肪酸在代谢过程中又生成氧自由基,使动物产生氧化应激,影响恢复期动物代偿生长的能力和肉品质[4]。目前关于育肥猪代偿效应的研究较少,本试验旨在利用代偿生长原理,初步探究限饲和补偿对育肥后期猪胴体品质、肉品质、抗氧化能力以及肌内脂肪代谢的影响,为生猪育肥后期优质猪肉的生产提供新的策略。
1 材料与方法
1.1 试验设计与饲养管理
试验于2019年11月5日至2019年11月21日在天津市武清区下伍旗天津畜牧研究所基地进行。选取30头体重为(115±1.5)kg、日采食量相近(3.4±0.2)kg、健康状态良好的“杜×长×大”三元杂交猪作为试验动物。试验采用单因素完全随机设计,将30头猪随机分成三组,每组10头猪,每头猪单栏饲养。预饲期3 d,试验期为14 d。对照组:自由采食;限饲组(FAST):每日提供2.5 kg饲粮,限饲14 d;补饲组(FAST+FED):每日提供2.5 kg饲粮,限饲7 d,之后7 d恢复自由采食。限饲组每天分3次于07:00(1 kg)、13:00(0.5 kg)和20:00(1 kg)饲喂。试验期间生猪自由饮水,按照猪场常规的要求进行饲养管理。
1.2 基础饲粮
本试验饲粮结合育肥猪营养需要特点,参照美国NRC(2012)猪营养需要中消化能(DE)和粗蛋白质(CP)的推荐量。各种营养水平均按照饲料原料实际测得的数据计算。基础饲粮组成及营养水平见表1。

表1 基础饲粮组成及营养水平(风干基础)
1.3 样品采集
试验期结束前一天空腹12 h,每组选择6头屠宰,宰前使用高压电击致昏后放血、屠宰,取背最长肌中段,除去脂肪和筋腱,灭菌手术刀切出肌肉,放入无菌的冻存管,做好标记立刻转移至液氮保存,分离背最长肌完整中段约10 cm放入4 ℃保温箱,测定肌肉感官品质。
1.4 测定指标
1.4.1 胴体品质、肉品质测定 胴体品质参照郑梓等[5]的方法进行。肉品质测定参照NY/T 821-2004行标猪肌肉品质测定技术规范测定。将新鲜肌肉样品切0.5 cm小块后,真空冷冻干燥机冻干12 h,低温粉碎机粉碎备用。使用全自动脂肪分析仪(ankom XT10),抽提脂肪,计算结果。计算公式为:肌内脂肪含量(%)=(肌内脂肪重/样本重)×100。甘油三酯使用试剂盒测定,试剂盒购自北京普利莱基因技术有限公司,具体步骤按照说明书操作检测。肌肉样本称取0.1 g,加入0.9 mL生理盐水,匀浆,离心提取上清液,东芝120全自动生化仪检测胆固醇含量,以每 mg蛋白浓度对胆固醇含量进行校正。蛋白浓度使用BCA蛋白检测试剂盒进行测定,试剂盒购自北京普利莱基因技术有限公司,下同。
1.4.2 氧化应激指标的测定 总抗氧化能力(T-AOC)、总超氧化物歧化酶(T-SOD)、谷胱甘肽过氧化物酶(GSH-Px)、过氧化氢酶(CAT)、丙二醛(MDA)均采用南京建成生物工程研究所的试剂盒测定。肌肉前处理方法同胆固醇测定,以每 mg蛋白浓度对含量进行校正。
1.4.3 脂肪酶活性测定 激素敏感性脂肪酶(HSL)、脂肪酸合成酶(FAS)均采用南京建成生物工程研究所的ELISA试剂盒测定。肌肉前处理方法同胆固醇测定,以每 mg蛋白浓度对含量进行校正。
1.4.4 Western blotting 取-80 ℃冻存肌肉样,加入预冷RIPA裂解缓冲液(碧云天),全自动组织研磨仪(JX-FSTPRP-24)匀浆裂解,12000×g 离心10 min后取上清液即为总蛋白溶液。30 μg蛋白上样,电泳,转膜,封闭,孵育一抗和耦联二抗。具体步骤参照闫峻[6]等的方法进行。甘油醛-3-磷酸脱氢酶(GAPDH)作为内参蛋白分析蛋白条带灰度值。过氧化物酶体增殖物激活受体 γ(PPARγ)、脂肪酸结合蛋白-4(FABP-4)、HSL、解偶联蛋白-1(UCP-1)等抗体均购自Abcom。
1.4.4 数据统计及分析 本试验数据采用 Excel 2013 软件进行整理,使用SPSS 20.0 中单因素方差分析(one-way ANOVA)和Duncan氏多重比较方法分析数据显著性,使用平均值±标准误(SE)表示统计结果。P<0.05 表示差异显著。
2 结果与分析
2.1 营养代偿效应对胴体性状和肉品质的影响
由表2可见,与对照组相比,FAST组和FAST+FED组中肉色黄度b*显著降低(P<0.05),背膘厚度、眼肌面积、肌内脂肪、甘油三酯、胆固醇、pH24 h、亮度L*、大理石纹评分等有降低趋势(P>0.05),pH45 min、红度a*、失水率、剪切力等有升高趋势(P>0.05)。与FAST组相比,FAST+FED组背膘厚度、眼肌面积、肌内脂肪、甘油三酯、pH45 min、亮度L*、大理石纹评分有提高趋势,分别提高了4.9%、0.4%、17.7%、12.1%、1.2%、7.7%、11.8%,胆固醇、红度a*、黄度b*、失水率、剪切力有下降趋势,分别下降了3.9%、0.9%、12.9%、8.5%、0.9%。
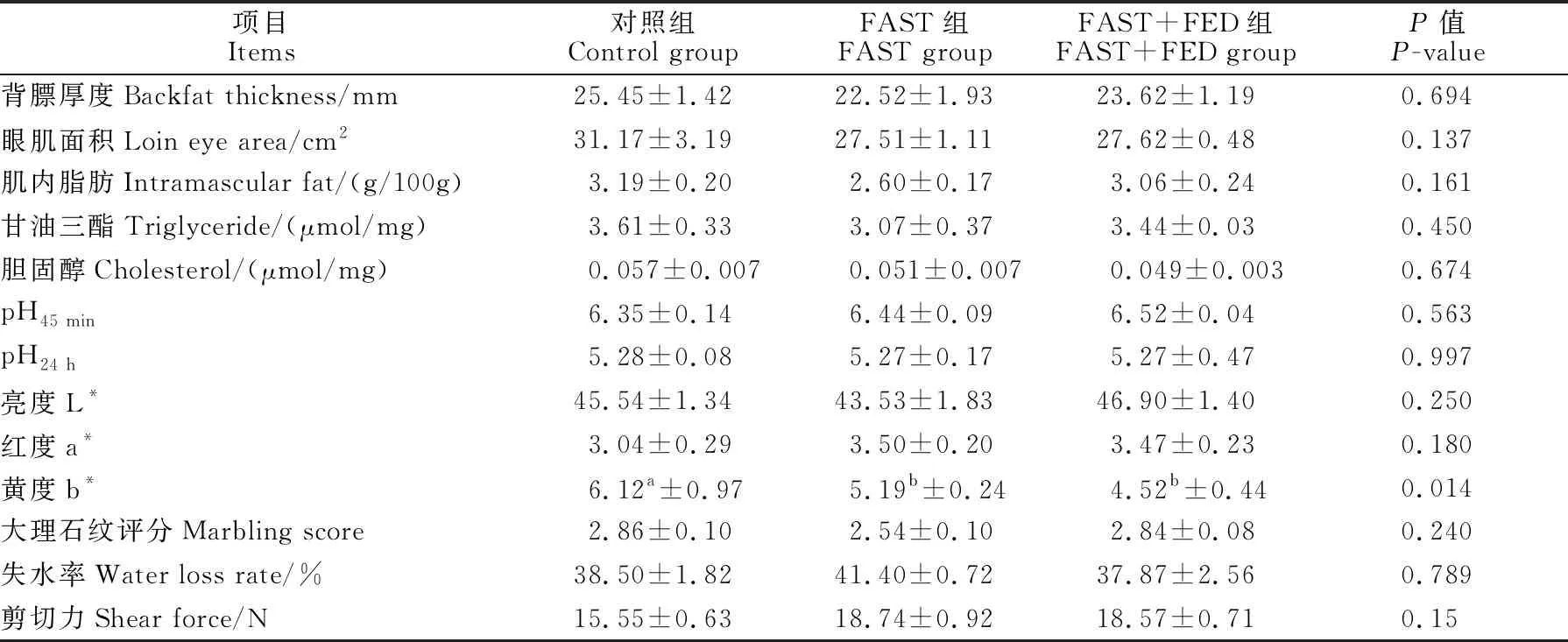
表2 营养代偿效应对育肥猪胴体性状和肉品质的影响
2.2 营养代偿效应对肌肉抗氧化能力的影响
由表3可见,与对照组相比,FAST组育肥猪背最长肌中谷胱甘肽过氧化物酶(GSH-Px)活性以及总抗氧化能力(T-AOC)、丙二醛(MDA)含量显著降低(P<0.05),过氧化氢酶(CAT)活性差异不显著(P>0.05);FAST+FED组中总超氧化物歧化酶(T-SOD)活性、总抗氧化能力、GSH-Px活性、CAT活性差异不显著(P>0.05),与FAST组相比显著提高了31.3%,46.2%和39.7%(P<0.05)。
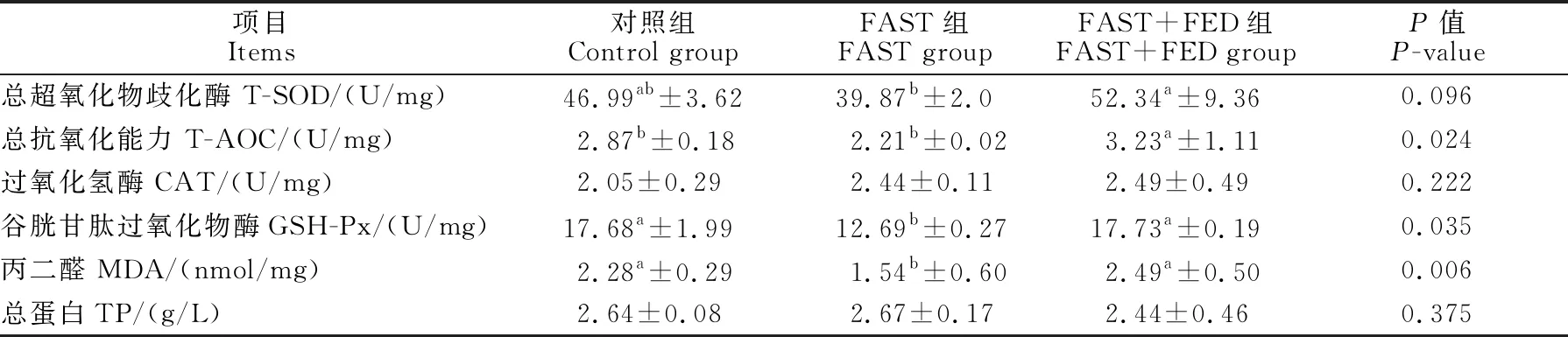
表3 营养代偿效应对育肥猪肌肉组织抗氧化能力的影响
2.3 营养代偿效应对肌肉脂代谢相关蛋白表达的影响
由图1可见,与对照组相比,FAST组中PPARγ蛋白表达量显著下调(P<0.05),FABP-4蛋白表达量无显著差异(P<0.05),HSL、UCP-1蛋白表达量显著上调(P<0.05);FAST+FED组中PPARγ蛋白表达量显著下调(P<0.05),FABP-4、HSL、UCP-1蛋白表达量无显著差异(P>0.05);与FAST组相比,FAST+FED组PPARγ蛋白表达量显著上调(P<0.05),HSL、UCP-1蛋白表达量显著下调(P<0.05)。
2.4 营养代偿效应对肌肉组织脂代谢酶活性的影响
为了探究营养补偿效应对育肥猪肌肉中脂代谢酶活性的影响,检测了育肥猪背最长肌肌肉中的相关脂代谢酶HSL和FAS的活性,结果见表4。与对照组相比,FAST组中FAS活性都显著降低(P<0.05),HSL活性均显著升高(P<0.05); FAST+FED组HSL活性差异不显著(P>0.05),FAS活性显著高于对照组(P<0.05)。
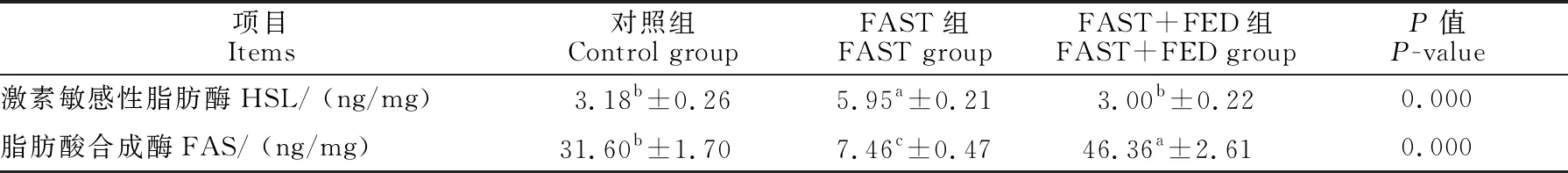
表4 营养代偿效应对育肥猪肌肉组织脂代谢酶活性的影响
3 讨 论
3.1 营养代偿效应对胴体性状和肉品质性状的影响
背膘厚与眼肌面积是评价育肥猪胴体性状的重要指标。研究表明,限饲主要影响脂肪的沉积,适量限饲对胴体品质影响不显著[7]。限饲能够调节猪背脂的沉积使背膘厚度有降低的趋势[8],补偿生长可以促进皮下脂肪的沉积[9]。本试验中,进行限饲和补饲对育肥猪背膘厚与眼肌面积无显著性影响,但限饲使背膘厚度和眼肌面积呈现下降趋势,补饲减弱了背膘厚和眼肌面积的下降幅度。说明在本试验条件下补饲对猪胴体品质有一定的代偿效应。脂肪是影响肉品质和营养的主要物质,是评价肉品质的重要指标[9]。肌内脂肪主要由甘油三脂、磷脂(膜脂)、胆固醇等构成。甘油三酯在肌纤维细胞质中的脂滴中储存,穿插在纤维束间,是肌内脂肪含量增加的主要因素[10]。胆固醇是动物细胞膜的重要组成部分,其在肌肉组织中含量普遍较低[11]。而Heyer等[9]的研究结果表明,在猪生长期(30~70 kg)时限饲,育肥期(70 kg~110 kg)时恢复自由采食,会显著降低110 kg体重时的肌内脂肪含量,而显著增加皮下的脂肪组织。本试验中,各组间的肌内脂肪、甘油三酯、胆固醇的含量无显著性差异,但限饲组中肌内脂肪、甘油三脂、胆固醇含量比对照组下降,补饲使肌内脂肪和甘油三脂含量比限饲组提高但仍低于对照组水平。这提示我们在猪不同阶段和进行不同时间的限饲对育肥猪脂质沉积的影响效果会有所不同。
肉品质是消费者最关注的性状特征之一。Heyer等[9]的研究表明,代偿生长几乎不影响育肥猪110 kg时的肉质。本试验条件下,限饲与补饲显著降低了肉的黄度b*值,但对育肥猪背最长肌肉品质其他感官性状影响不显著。但有些研究者认为营养限制的条件下会导致机体生理代谢过程的减慢,从而影响肉品质[11]。兰儒冰等[12]和王震等[13]的研究显示限饲可使内蒙羔羊和藏羊背最长肌pH接近7,失水率上升,补偿结束后恢复至正常水平。本试验中,限饲使育肥猪背最长肌剪切力、pH45 min、失水率有提高的趋势,补饲使育肥猪背最长肌剪切力和失水率降低,pH45 min升高,与前人结果[14]基本一致。整体来看,育肥期限饲有降低育肥猪的胴体和肉品质性状的趋势,先限饲再补饲产生的营养代偿效应可以降低因限饲造成的育肥猪胴体和肉品质性状下降的风险。
3.2 营养代偿效应对背最长肌抗氧化能力的影响
本试验条件下,限饲显著降低了育肥猪背最长肌中总抗氧化能力、GSH-Px活性以及MDA含量。说明限饲状态下,育肥猪肌肉组织中抗氧化水平处于较低的水平,补饲后育肥猪背最长肌中T-SOD、GSH-Px活性和总抗氧化能力比限饲状态下显著升高,表明了营养代偿效应可以增强育肥猪肌肉的抗氧化能力,这与苏艳莉等[15]研究结果一致。
3.3 营养代偿效应对肌内脂代谢蛋白表达的影响
饥饿状态下,脂肪作为机体重要的能源物质被消耗分解。适宜的饥饿(短期饥饿和限制性饮食等)会激活机体对能源物质的利用,改变能量代谢的方式[3]。Costa等[2]研究表明生长途径影响牛胸肌肉中脂代谢过程,在限饲期间脂肪酸β-氧化相关的基因表达增加,而在补偿后脂肪酸合成相关的基因表达占主导。限饲降低了肌内脂肪和甘油三脂含量,补饲产生代偿效应促进了肌内脂肪和甘油三脂含量的增加。PPARγ是哺乳动物调控脂肪细胞分化和脂类代谢的关键转录调控因子[16],本试验中限饲组中脂肪分化相关酶PPARγ表达量较对照组显著降低,说明在限饲过程中脂合成过程被抑制,补饲组中PPARγ含量较限饲组显著上调,说明营养补偿效应通过上调PPARγ表达促进了肌内脂肪的沉积。HSL和UCP-1是调控脂肪分解和产热的关键酶,在机体能量供应不足时,HSL可以催化甘油三酯的分解释放脂肪酸,UCP-1是分布于线粒体内膜上的产热蛋白,在能量供应不足时可以促进机体产热,调节能量代谢[17-18]。本试验中限饲组中HSL和UCP-1表达量显著提高,说明在限饲过程中脂解作用增强,肌内脂肪沉积减弱,补饲组中HSL和UCP-1表达量较限饲组显著降低,说明营养补偿效应通过抑制脂解作用表达促进肌内脂肪的沉积。为了进一步验证该结论,脂代谢相关酶的酶活结果显示HSL酶活在限饲状态下显著提高,补饲降低了HSL的活性,与钱宝英等[17]研究结果一致。FAS是脂肪酸合成的关键酶[19],限饲状态下FAS活性被显著降低,补饲组显著提高了FAS活性,且FAS活性显著高于对照组。证实了代偿效应可以通过上调脂合成,抑制脂解来促进肌内脂肪的沉积。
4 结 论
本试验条件下,育肥期适度限饲对猪胴体及肉品质无显著性影响。育肥期限饲显著降低了育肥猪背最长肌抗氧化能力,营养代偿效应可以通过提高抗氧化酶活性恢复育肥猪抗氧化能力。育肥期限饲下调脂合成过程,上调脂解过程,减少了肌内脂肪的沉积;补饲产生的营养代偿效应可以通过下调脂解过程,上调脂合成过程促进肌内脂肪的沉积。
猜你喜欢
销售与市场(营销版)(2022年3期)2022-11-13
能源研究与信息(2022年3期)2022-10-29
肉类工业(2022年8期)2022-09-14
临床肝胆病杂志(2022年8期)2022-09-07
养猪(2022年1期)2022-02-17
疯狂英语·新阅版(2021年8期)2021-09-10
时代英语·高二(2021年4期)2021-07-29
银行家(2020年10期)2020-11-06
保健与生活(2019年13期)2019-08-27
时尚北京(2016年12期)2017-01-16
