
山地隔离和空间距离对高黎贡山灰腹地莺(Tesia cyaniventer)鸣唱特征的影响
2021-11-26 10:21:50冯莹莹李兴权
生态学报 2021年21期
冯莹莹, 梁 丹, 李兴权, 罗 旭,*
1 西南林业大学生物多样性保护学院/云南省高校极小种群野生动物保育重点实验室, 昆明 650224 2 江西省林业科学院野生动植物保护研究所, 南昌 330032 3 普林斯顿大学公共与国际事务学院, Princeton 08540 NJ, USA
鸣唱在鸟类繁殖中具有重要作用[1],同时也存在广泛的空间变异,从而限制种群间交流、促进种群分化,因此对于依赖鸣声进行配偶选择的鸟类,鸣唱变异被认为是鸟类物种分化的最初步骤[2- 3]。多种因素可导致不同鸟类种群产生鸣声变异,包括地理因素、生境、性选择、奠基者效应、鸣声漂变和遗传差异等[4- 5]。地理因素中,空间距离对鸣唱的影响有较多的研究,如森林斑块间大山雀(Parusmajor)鸣唱曲目的共享研究,表明距离越远的不同种群间鸣唱曲目的共享越少[6]。金冠带鹀(Zonotrichiaatricapilla)的鸣唱变异也与空间距离有关[7]。环形种暗绿柳莺(Phylloscopustrochiloides),环绕青藏高原连续分布,高原两侧的种群从南至北鸣唱差异逐渐增大,导致分布区两端的种群鸣声差异巨大,即使在中亚交汇也无法交流[3,8]。以及欧亚大陆的广布种大山雀的major和minor亚种在鸣声上的分化,并且在西伯利亚阿穆尔河谷中部出现生殖隔离[9- 10],因此minor亚种被提升为种。以上研究表明的较为一致的规律是,鸣唱差异随着空间距离的增加而加大。地理上的另一个因素是隔离,比如高山、海峡等,因为不可逾越,导致被阻隔的临近种群无法交流从而形成鸣声特征上的“间断”。比如画眉(Garrulaxcanorus),指名亚种(G.c.canarus)鸣唱更为复杂、音节型更多、组合形式也更多,台湾亚种(G.c.taewanus)鸣唱的简化推测与隔离分化和奠基者效应有关,此后的分子遗传研究建议将台湾亚种提升为独立的种[11- 12];印度西南部分布的白腹短翅鸫(Brachypteryxmajor),也因受到地理隔离的影响,两个种群的鸣声、遗传和表型均出现分化,因此被分为两个不同的种[13]。地理隔离的存在,使得鸟类鸣唱的地理变异更加复杂,它和空间距离可能会共同影响鸟类鸣唱变异,比如泰国北部的鹊鸲(Copsychussaularis),鸣唱特征就受到两者的共同作用[14]。
在山地系统中,复杂地形对鸟类的鸣声有较大影响。地理阻隔和空间距离的作用往往同时存在,比如山脊对一些较低海拔分布的鸟类是难以跨越的地理阻隔,较大的空间距离也可能在一定程度限制种群间的基因交流[15]。然而,山地的地理阻隔和空间距离如何影响鸟类的鸣唱特征,尤其是对森林鸟类鸣声特征的影响研究仍然十分缺乏。对于运动能力较弱的地栖性森林鸟类,我们推测鸟类的鸣唱特征呈现出“隔离-距离”共同作用的格局,在微地理尺度上隔离的作用会更大。
为验证这一假设,选取灰腹地莺(Tesiacyaniventer)为研究对象。灰腹地莺为雀形目、树莺科(Cettiidae)、地莺属(Tesia)的小型地栖性留鸟(体长约9.5 cm)。在国内,主要分布于西南山地的中、高海拔湿性常绿阔叶林中,一般多单个或成对活动于林下潮湿山谷、或溪流附近,行踪较为隐蔽[16]。灰腹地莺的领域性较强,繁殖期鸣声十分洪亮而易于识别,是研究鸟类鸣声的良好对象。研究地点选择在高黎贡山,它是怒江(Salween River)和伊洛瓦底江(Irrawarddy River)的分水岭,南北走向,山体巨大,主峰嘎娃嘎普海拔5128 m,山脊平均海拔超过3100 m[17]。在高黎贡山,灰腹地莺分布海拔为2000—2800 m,在海拔超过3000 m的山脊无灰腹地莺记录[18- 19],因此山脊极有可能成为灰腹地莺难以跨越的障碍;而高黎贡山南北绵延数百公里的跨度也为鸣声地理差异提供了空间上的可能。基于前述假设,本文通过对高黎贡山南北段的灰腹地莺东西坡的鸣唱比较,研究山体隔离和空间距离对该物种鸣声的影响,旨在验证2个问题:(1)分布在高黎贡山山体同一侧的种群,只要距离足够远,种群间会产生鸣唱特征的变化;(2)分布于山体两侧的种群,鸣唱差异会大于同侧分布的种群。
1 研究方法
1.1 野外录音采样
根据灰腹地莺的海拔分布及栖息生境,我们在高黎贡山选择4个地点录制灰腹地莺的鸣声(图1A)。南段采样点在云南省怒江傈僳族自治州泸水市片马垭口(海拔3150 m)的东西坡两侧(SE为南段东坡,SW南段西坡)、野外录音沿S316省道行进,集中在海拔2000—2800 m之间,录音时间为2014年4—5月、2018年4—5月。北段采样点位于怒江州贡山县的独龙江垭口(海拔3172 m)的东西两侧(NE为北段东坡,NW北段西坡),野外采样沿S212省道行进,集中在海拔2000—2600 m之间,录音时间为2016年5—6月、2018年5—6月。南、北两个点相距约200 km,北段灰腹地莺繁殖晚于南段,因此野外录音采集时间也较晚。野外观察表明,灰腹地莺的领域较小,录制时保持200 m以上的间隔,以确保不对同一只个体重复录音。在同一个地点至少间隔2年采集鸣声,以尽可能排除假重复。
录音设备采用便携式数字录音机Sony PCM—D100(日本),或便携式数字录音机Zoom H4n(日本),配以外接强指向性话筒Rode NTG—3(澳大利亚)。采样精度为16 bit,采样频率为44.1 kHz,文件保存为.WAV格式[20]。录音时间集中在晨昏,灰腹地莺因声音洪亮而易于发现,声音录制尽可能地长,直至它停止鸣叫或飞走,以获得足够的分析样本。
1.2 鸣唱参数的测量
利用Raven Pro 1.5鸣声分析软件生成语图,参数设置:Window Type: Hamming; Window size:1024 samples; Overlap:50%; hop size:512 samples, DFT size: 1024 sample, Grid spacing:43.1 Hz。此外,用Raven Pro 1.5去除1500 Hz以下和6000 HZ以上的背景噪音,以获得清晰的语图。在Raven 1.5中筛选录音样本中,去除语图不清晰或句子数不足10的个体。通过筛选,共对58个个体的鸣声进行分析。
参照其他鸟类的鸣唱特征的相关研究[11,21]对本文所涉及的鸣声术语进行如下定义。音节指一个或几个固定组合在一起的音素总是以相同的顺序一起出现,代表语义的最小单位。音节型指音节的结构特征,主要包括音素组成和排列顺序。鸣唱型为鸣唱句子的结构特征,主要包括音节组成和排列顺序,也称句式、句法。起始音素意为句首出现的单一的、持续时间较短的音素,结构相对固定,音量较弱。句子表示有音素或音节的连续段落,句子间有明显的时间间隔。曲目则代表每个个体或种群的所有鸣唱类型或音节类型。
鸣唱的参数根据Catchpole[22]和Matessi等[23]分为质量性状和数量性状。质量性状参数包括:每个鸣唱的音节数、音素数,获得每个个体的全部音节型、鸣唱型。本文依据Baker等[24]对音节的划分标准:相邻两音素间隔小于0.02 s为同一音节,间隔在0.1 s以上视为不同音节;间隔在0.02—0.1 s之间,若音素复合体中的每个音素在所统计的鸣唱中单独出现过,视为不同音节,若无则视为一个音节。句子的划分标准为:相邻两个句子(末尾音节和起始音节间)的时间间隔若在1 s以上则为不同鸣唱,否则为同一鸣唱[21]。此外,由于灰腹地莺一般有一个短暂且易于识别的首音节作为鸣唱的开始,我们也将此作为判断是否为同一鸣唱的辅助标准。鸣唱型的划分采用75%原则,即两个句子间共享75%以上的音节类型视作同种鸣唱型[25]。音节型的划分参考雷富民等[26],将声谱图上形状大致相同、频率有微小变化的音节视为同一音节类型。数量性状的测量以句子为单位,对每个灰腹地莺个体的所有鸣唱句子进行测量并取平均值,测量的参数包括句子的最高频率(Fhigh)、最低频率(Flow)、中心频率(Fcent)、峰频率(Fpeak)、句子时间(D)、平均熵(EntrAvg)、起始音节的频率(FFirNot)、起始音节与第二个音节的时间间隔(Tinter)、句子平均音节数(Nsyll)、句子平均音素数(Nnote)。其中中心频率(Fcent)指位于句子频宽中间的频率;峰频率(Fpeak)为句子中能量最高时的频率。
1.3 声谱分析
首先验证所录制的个体是否覆盖足够的鸣唱型和音节型,使用R包”iNEXT”计算鸣唱型和音节型的取样覆盖度[27]。结果表明,四个种群录制的个体足够覆盖各个种群的鸣唱型和音节型(图2,图3)。然后,采用Sørensen相似性指数[28]来衡量不同地理种群的音节型及鸣唱型的共享程度:
Si=2c/(a+b)
式中,c为种群A与种群B共享数;a和b分别为种群A和种群B的类型数。相似性系数(Si)的变化范围为0—1,当Si值为0,说明种群间不存在音节或鸣唱型共享,若Si值为1,则两种群间的音节型和鸣唱型完全共享。

图2 灰腹地莺音节型取样覆盖度曲线Fig.2 The sampling coverage (SC) curve of the syllable types of the Tesia cyaniventer
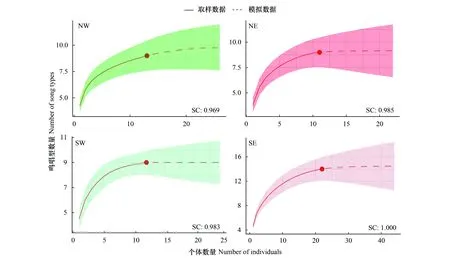
图3 灰腹地莺鸣唱型取样覆盖度曲线Fig.3 The sampling coverage (SC) curve of song types of the Tesia cyaniventer
为对比不同种群灰腹地莺的鸣唱变异的程度,首先利用单因素方差分析(One-way ANOVA)对所有鸣声特征分别进行种群间差异的比较,随后对种群间差异显著的鸣声特征进行两两种群间的多重比较(Tukey′s HSD)。我们将数据进行对数转换(log-transformed)以使其符合正态分布。以上分析在R 3.5.3 软件中进行[29]。文中数据以Mean±SD表示。
2 结果
2.1 灰腹地莺的鸣唱特征
野外共录制到76个灰腹地莺个体的鸣唱声,选择语图质量较高的个体58只进行后续分析,其中南段东坡(SE)22只、西坡(SW)12只;北段东坡(NE)11只、西坡(NW)13只。共计测量了1268个句子的数量性状(表1)。

表1 高黎贡山灰腹地莺的鸣唱特征
2.2 鸣唱型、音节型及共享
灰腹地莺4个种群的鸣唱型共38种(图4),其中南段东坡(SE)、西坡(SW)分别为14种和9种;北段东坡(NE)、西坡(NW)分别为9种。在山体两侧,仅SW和SE共享1种鸣唱型 (Si=0.087),其他无共享(Si=0);在山体同侧,仅西坡的NW和SW间共享2种(Si=0.222),东坡无共享(Si=0)(表2)。灰腹地莺的音节型共有58种(图5),其中SE和SW分别为28和22种;NE和NW分别为23种和19种。山脉两侧SW和SE共享11种 (Si=0.440),NW和NE共享8种 (Si=0.381);山脉同侧的SE和NE共享7种 (Si=0.275),SW和NW共享10种 (Si=0.488) (表2)。鸣唱型和音节型的共享情况具有显著相关性(r=0.819,P<0.05)。

图4 灰腹地莺的58种音节型Fig.4 The 58 syllable types of male songs of Tesia cyaniventer
2.3 不同种群鸣唱的差异
单因素方差分析表明:种群间的鸣唱差异主要体现在6个特征上:包括最低频率(Flow)、频率宽度(Fdelta)、中心频率(Fcent)、起始音节频率(FFirsNot)、首二音节的时间间隔(Tinterv)、平均音节数(Nsylla),其他鸣声参数上无差异(表3,图1C)。两两种群间鸣声特征比较结果显示:在既存在山体隔离且空间距离较远的种群间(NW-SE和NE-SW)均有3个鸣声特征存在显著差异;而存在山体隔离但空间距离较近的种群间(NW-NE和SW-SE)均有2个鸣声特征存在显著差异;而不存在山体隔离但空间距离较远的种群间(NW-SW和NE-SE)仅有1个鸣声特征存在显著差异(表4,图1C、1D)。以上结果表明,山体隔离和空间距离均对灰腹地莺的鸣声特征的种间差异有影响,但山体隔离比空间距离对鸣声特征的影响更加显著。
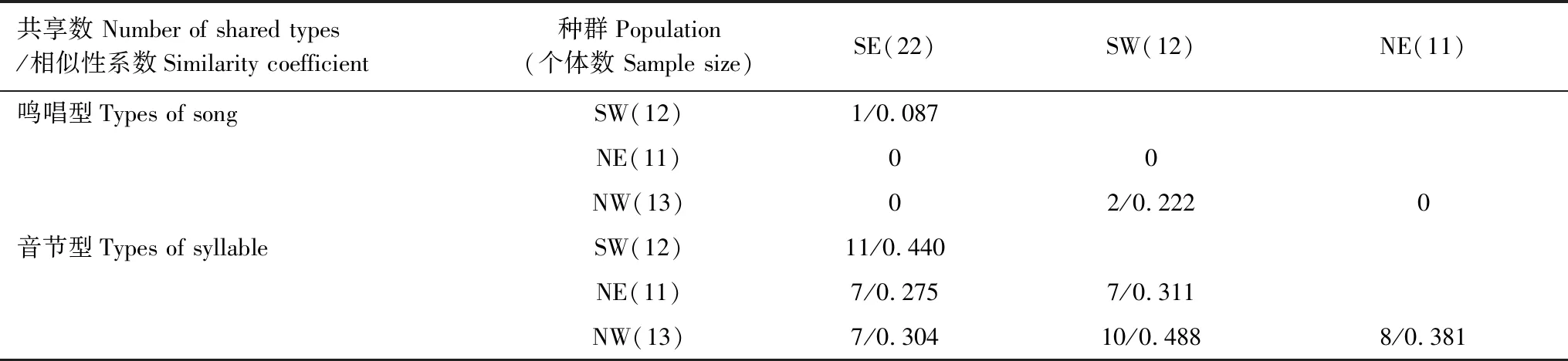
表2 高黎贡山灰腹地莺4个种群鸣唱型与音节型的共享程度
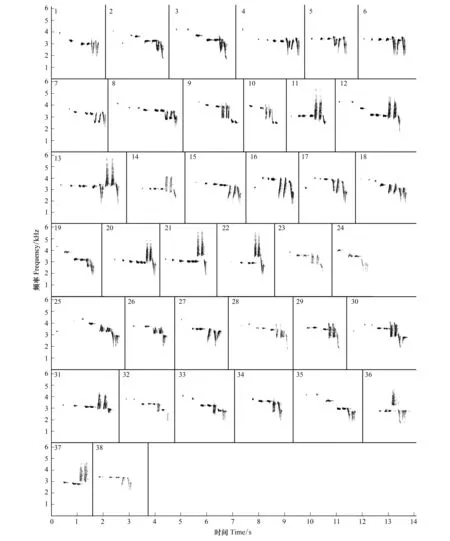
图5 灰腹地莺的38种鸣唱型Fig.5 The 38 song types of male songs of Tesia cyaniventer
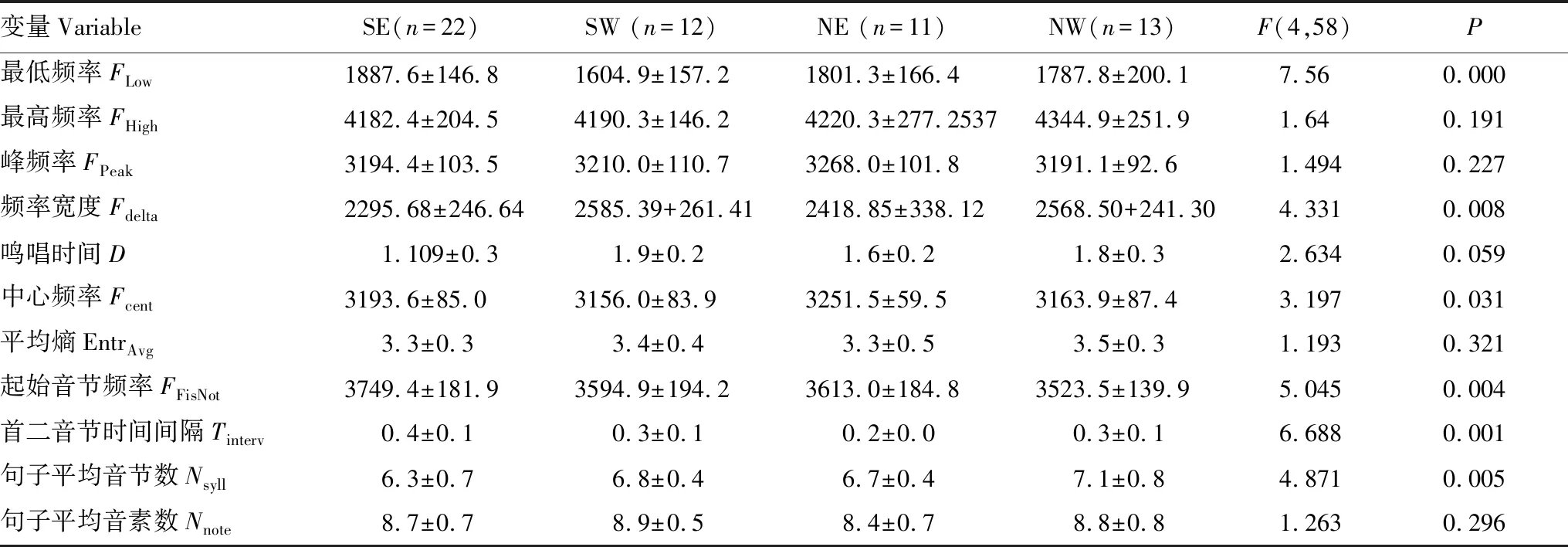
表3 不同种群灰腹地莺鸣唱特征及单因素方差分析
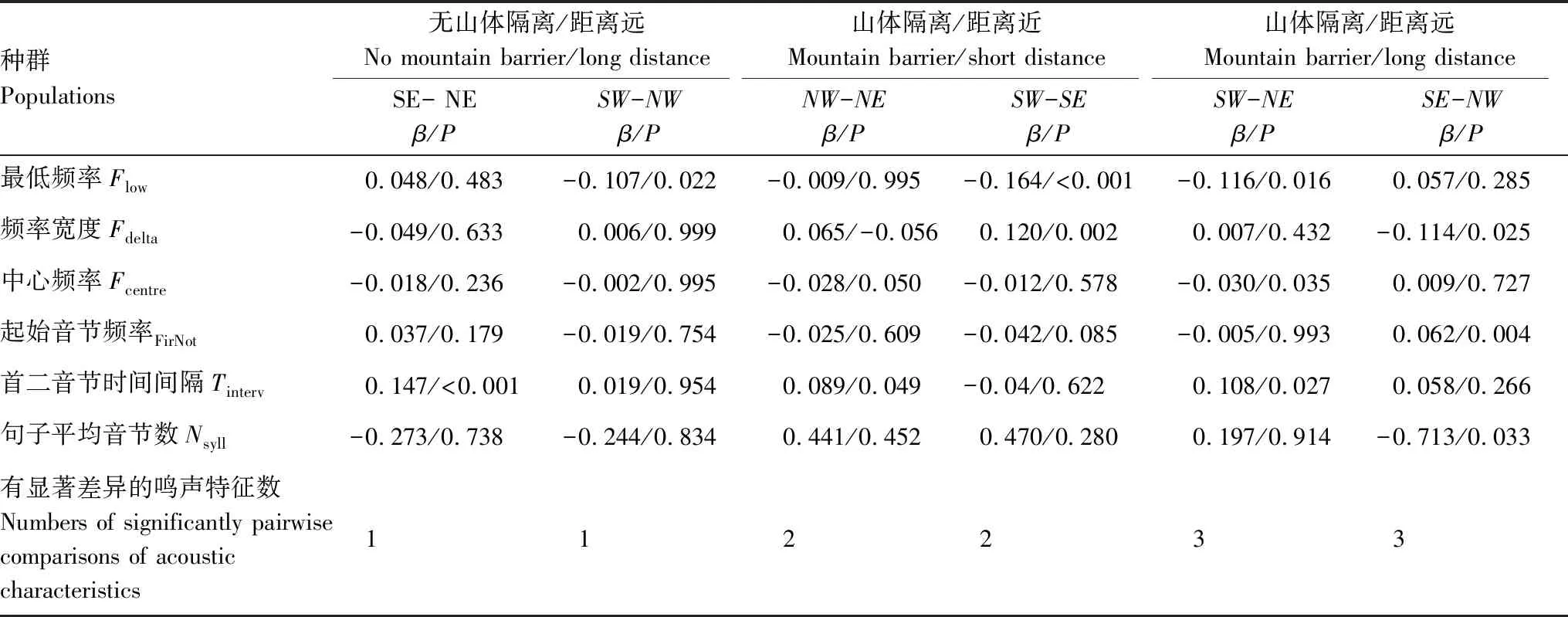
表4 灰腹地莺4个种群间鸣唱特征的两两比较
3 讨论
3.1 灰腹地莺的鸣唱特征
本研究从质量性状和数量性状上对灰腹地莺的鸣唱进行了分析,根据鸣唱的组成来看,灰腹地莺的曲目中有多个鸣唱型,平均每个个体的鸣唱型为(4.43±1.2)种,属于多才多艺型[30]。灰腹地莺的鸣唱通常是唱一种句子后无规律的转换为下一种,属于“瞬时变化型”(immediate variety, ABCABC…),区别于重复的唱同一种鸣唱型然后再转为另一鸣唱型的“平稳过渡型” (eventualvariety, AAAAABBBBC…)[30]。本研究中,灰腹地莺的不同鸣唱句子的时长和节奏较为相似,主要差异源于音节,和红翅黑鹂(Agelaiusphoeniceus)[31]的鸣唱句型结构相似。这被认为是性选择的结果,因为已有研究表明,音节不同但句子时长和节奏类似形成的多曲目是由于雌性偏好选择的结果[32],句子时长和节奏具有较大差别形成的多曲目则是由于鸣唱的功能分化引起的[33]。
从数量性状上看,灰腹地莺的鸣唱频率范围在(1790.37±192.11)—(4227.65±225.11)Hz之间,与同科其他鸟类相比,其鸣唱低频及高频均明显低于鳞头树莺(Urosphenasquameiceps)[34]和强脚树莺[35],可能与灰腹地莺偏好密闭度高且潮湿的林下环境有关。生活在森林密闭地区的鸟类鸣声频率普遍低于开阔地区的鸟类,因为森林鸟类鸣声的高频部分容易发生反射和衍射,造成能量的衰减和声源位置的模糊,减少接收者得到的信息量[36],为保证声音的有效传递,灰腹地莺的鸣唱频率偏低有利于适应高黎贡山植被茂密的环境特征。
在野外录音时还发现,灰腹地莺个体有存在仿效其他鸟类鸣唱的情况,效鸣对象为丽色奇鹛(Heterophasiapulchella)。效鸣句子中也存在起始音节,但丽色奇鹛的鸣唱并不存在该音节。研究表明,15—20%的雀形目鸟类有效鸣现象[37]。错误学习是效鸣产生的一个重要原因[38],因为周边环境中充满其他鸟类的歌曲[39],如大盘尾(Dicrurusparadiseus)[40]和绿篱莺(Hippolaisicterina)[41]的幼鸟就存在这种错误学习的现象。研究区域内丽色奇鹛较多而且鸣声清晰洪亮,我们推测灰腹地莺在声音学习的敏感期误学了丽色奇鹛的鸣声。
3.2 灰腹地莺鸣唱的地理变异
鸣唱质量性状反映的是行为特征,是通过鸣声学习行为传承下来的结果,鸣唱型、音节型等是都通过学习而来[42]。灰腹地莺4个种群间的鸣唱型和音节型的地理变异并未表现出一致性,鸣唱型明显受到山脉阻隔和空间距离的共同作用,种群间共享程度非常低,这与鸣唱在各个种群中的遗传有关[43]。由于鸣唱具有明显与繁殖相关的功能,性选择的作用以及种群内个体鸣唱漂变的影响[1],可能导致鸣唱的快速进化。音节型在4个种群间的共享程度明显高于鸣唱型,不论同侧或异侧种群都能接近50%共享,表明高黎贡山山脉和距离的阻隔作用主要体现在更高层次的鸣声特征变化(鸣唱型),而不是音节型的差异,因为音节的不同以及音节组合的变化均会带来鸣唱型的变化,这在画眉的研究中也有类似结果[11]。
鸟类鸣唱数量性状反映的信息与质量性状有所差异,数量性状更多地受到环境影响[5,42]。有研究表明,即使地理因素存在时,鸟类鸣声的数量性状也会保持遗传的相似性[44],如暗绿柳莺的鸣声变异[8]。因此,鸟类鸣唱的数量性状比质量性状可能会更多地受到遗传与环境因素的影响。灰腹地莺鸣唱差异主要体现在频率、首二音节时间间隔等6个参数上。最低频率(Flow)、中心频率(Fcent)等参数在不同种群中体现出不一致性,可能灰腹地莺适应微生境而在时间增量和频率特征上作出的调整,以进行高效的声通讯[45]。比如鸣唱最低频率(Flow),南段西坡(SW)显著低于其他3个种群(图1C),推测是因为高黎贡山片马风雪垭口西坡常年盛行印度洋西南季风[17],声音在风中传播时容易能量衰减[46],因此,该地种群以降低低频来保证声音传播效率。而北段高黎贡山西侧有担当力卡山脉阻挡西南季风,因此并未出现低频降低的现象。
本文比较的两种地理因素均可对灰腹地莺的不同种群的鸣声特征产生影响,但山体隔离比空间距离对鸣声特征的影响更为显著(图1D),这说明受地理隔离的种群可能比连续分布的种群更快出现鸣唱特征的差异[47]。这个结果与高山鹪鹩(Thryorchilusbrowni)的研究相近[48],不连续生境中的种群易在鸣声、形态和遗传上产生各自的适应,而连续生境依旧可以连接长距离种群间的沟通,使分布上连续的种群积累变异慢于隔离种群。
综上,灰腹地莺鸣唱特征具有明显的地理变异,山脉同侧的差异小于山脉两侧,说明山体隔离和空间距离均对该物种鸣声产生影响,但山体隔离作用更大。
猜你喜欢
英语世界(2023年12期)2023-12-28 03:36:06
东坡赤壁诗词(2023年2期)2023-05-30 15:02:14
光明少年(2023年4期)2023-04-29 00:44:03
植物多样性(2022年2期)2022-04-25 07:33:56
诗歌月刊(2021年8期)2021-08-25 18:54:10
中国宝玉石(2021年4期)2021-08-20 07:56:52
小天使·二年级语数英综合(2021年6期)2021-08-09 10:25:54
安徽师范大学学报(自然科学版)(2018年5期)2018-12-11 03:32:00
汉语世界(2017年6期)2017-11-24 06:27:28
中国三峡(2016年10期)2017-01-15 14:00:01
