
紫花苜蓿内生细菌的拮抗作用及优良拮抗菌株的鉴定和生物功能
2021-11-22 09:49刘彦超张贞明金梦军施兆荣崔凌霄魏立娟仲健新杨成德
草业科学 2021年10期
刘彦超,张贞明,金梦军,施兆荣,崔凌霄,魏立娟,仲健新,杨成德
(1. 甘肃农业大学植物保护学院,甘肃 兰州 730070;2. 甘肃省林业和草原局,甘肃 兰州 730070)
紫花苜蓿(Medicago sativa)是发展畜牧业的重要基础,有“牧草之王”的美誉,在世界各国广泛种植[1]。随着紫花苜蓿种植年限不断增长,种植面积不断扩大,苜蓿病害成为影响产业发展的重要问题,其中根腐病是苜蓿栽培中的主要病害之一,在美国、加拿大、澳大利亚、俄罗斯、日本、阿根廷和中国苜蓿主要种植区均严重发生[2]。紫花苜蓿根腐病多由镰刀菌(Fusariumspp.)引起,已报道的35 种苜蓿根腐病病原中镰刀菌达20 种[3]。镰刀菌产生的病害主要采取农业和化学药剂防治,其中,培育抗病品种是防治根腐病最有效的方法,但该病害的病原物比较复杂,因此苜蓿抗病品种的选育较为困难[4];化学防治在病害防治中效果显著,但长期、反复和大量使用农药会加剧环境污染问题,并且使病原物产生抗药性,与生态农业的发展目标相悖。微生物菌剂主要是利用自然界中微生物对病原菌的抗生作用、诱导产生系统抗性等来减少病原菌数量或提高寄主植物的抗病性。目前,利用植物内生细菌进行生物防治已成为研究热点之一。
植物内生细菌广泛定殖在植物体内,是会与植物建立和谐共生关系的一类微生物[5]。内生细菌可以提高植物对恶劣环境的适应能力,保证植物系统的生态平衡和健康发展,具有广泛的生物学作用,是不可多得的天然生物资源[6]。倪婕等[7]发现从白骨壤分离得到的内生细菌多样性较丰富,且部分具有广谱抑菌活性;李小杰等[8]从烟草根茎中分离、筛选出9 株对烟草(Nicotiana tabacum)青枯病菌(Ralstonia solanacearum)具有拮抗效果的菌株,并且在温室盆栽试验中发现菌株Rsa3 发酵液的防病效果可达79.17%,同时能够促进烟草苗根系生长;金梦军等[9]从青藏苔草内生细菌中筛选出具有广谱抑菌活性的菌株1Y4,对马铃薯炭疽病菌(Colletotrichum coccodes)和孜然根腐病菌(Fusarium solani)的抑菌率分别高达71.43%和73.33%,其固体发酵菌剂对黄瓜(Cucumis sativus)和辣椒(Capsicum annuum)均具有明显的促生效果。为此,本研究通过分离紫花苜蓿内生细菌,筛选、鉴定紫花苜蓿根腐病菌(Fusarium chlamydosporum)的优良拮抗菌株,并测定优良拮抗菌株的固氮、溶磷(无机磷)和分泌生长素(IAA)的能力,以期为苜蓿根腐病的生物防治提供菌种资源和依据。
1 材料与方法
1.1 材料
1.1.1 供试菌株
紫花苜蓿于2020 年6 月在甘肃农业大学校园内(103°42′26″ E,36°5′48″ N)采集,并对根、茎和叶内生细菌进行分离和保存。
1.1.2 供试指示菌
辣椒根腐病菌尖孢镰刀菌 (F. oxysporum)、紫花苜蓿根腐病菌厚垣镰孢菌(F. chlamydosporum)、茄镰孢菌(F. solani)、黄瓜枯萎病菌(F. oxysporum)、马铃薯枯萎病菌(F. oxysporum)、核桃枝枯病菌[燕麦镰刀菌(F. avenaceum)和锐顶镰刀菌(F. acuminatum)]均由甘肃农业大学植物保护学院植物病原细菌及细菌多样性实验室提供。
1.1.3 供试培养基
供试培养基分别为肉汁胨培养基(nutrient agar,NA)[9]、马铃薯葡萄糖培养基(potato dextrose agar,PDA)[9]、金 氏 培 养 基(King)[10]、Pikovaskaia 培 养基(pikovaskaia’s,PKO)[9]、阿 须 贝 无 氮 培 养 液(ashby nitrogen-free, Ashby)[10]。
1.2 方法
1.2.1 内生细菌的分离和纯化
将紫花苜蓿表面用灭菌蒸馏水清洗干净,用灭菌后的滤纸吸干表面水分,称取1 g 在无菌条件下用75%酒精表面消毒30 s,用无菌水冲洗5 次后再用0.1% 升汞表面消毒2~3 min,再次用无菌水冲洗5 次,彻底消毒后放入研钵,加入适量石英砂和5 mL 生理盐水研磨,静置数分钟,取上清液梯度稀释为10−1、10−2和10−3不同倍数的稀释液,每个梯度的稀释液各取0.1 mL 涂布于NA 培养基上,以最后一次洗涤水为对照,设置3 次重复,28 ℃培养3~5 d,通过观察进行分类保存备用[11]。
1.2.2 优良拮抗菌株的筛选
采用平板对峙法[10]测定抑菌效果。将供试指示菌紫花苜蓿根腐病菌接种于PDA 培养基上,25 ℃培养7 d 后于菌落边缘打取5 mm 直径的圆形菌饼,接种于PDA 培养基平板中央,在距菌饼四周30 mm处分别点接供试内生细菌,以只接种病原菌为对照,设置3 次重复,置于25 ℃条件下培养7 d 后,采用十字交叉法测量病原菌菌落直径并计算抑菌率[10],并对优良拮抗菌株进行抑菌谱测定,方法与拮抗菌株筛选方法相同。

1.2.3 优良拮抗菌株的生物功能测定
固氮能力测定:将震荡培养所得到的菌悬液接种于阿须贝无氮培养液中,并将细菌点接在阿须贝无氮平板上,7 d 后观察平板上是否有菌落以及液体培养基是否变浑浊,具体步骤参见文献[12]。
溶磷能力测定:将待测菌株涂布于Pikovaskaia培养基上,28 ℃条件下培养14 d,观察有无解磷圈以及解磷圈大小,确定菌株解磷能力,具体步骤参见文献[13]。
产IAA 能力测定:采用Salkowski 比色法测定菌株是否具有产IAA 的能力,具体步骤参见文献[13]。
1.2.4 优良拮抗菌株的鉴定
培养性状及形态观察:将内生细菌接种于NA培养基上,在28 ℃条件下培养,将连续培养12 h 的细菌进行革兰氏染色,显微观察菌体特征,将连续培养72 h 后的细菌进行菌落形态特征的观察。
16S rDNA 基因序列分析:利用TIANGEN 细菌基因组DNA 试剂盒提取拮抗菌株DNA,并用引物27F (5′-AGAGTTTGATCCTGGCTCAG-3′)和1492R(5′-GGTTACCTTGTTACGACTT-3′) 进 行PCR 扩增,PCR 扩增反应体系为50 μL:2×TSINGKE Master Mix 25 μL、DNA 2 μL、上下游引物各2 μL、双蒸水19 μL;扩增程序为:94 ℃预变性5 min;94 ℃变性1 min,48 ℃退火30 s,72 ℃延伸1 min,32 个循环;72 ℃延伸8 min。之后,将PCR 产物送兰州天启基因生物科技有限公司测序,将测序结果导入EzBioCloud数据库进行BLAST 比对,使用Mega7.0 软件构建系统发育树,确定其系统发育学地位。
1.3 数据分析
采用Excel 2010 进行数据整理,采用SPSS 17.0软件对数据进行统计分析。
2 结果与分析
2.1 内生细菌的分离和纯化
根据菌落形态、颜色等特征,从紫花苜蓿茎部、根部共分离纯化得到80 株内生细菌,编号为MS-1,MS-2,…,MS-80。
2.2 优良拮抗菌株的筛选
80 株内生细菌中有39 株对紫花苜蓿根腐病菌抑菌率在50%以上,占分离菌株总数的48.75%,抑菌率介于55.63%~63.96%;9 株抑菌率在61%以上,分 别 为MS-43 (63.96%)、 MS-40 (63.75%)、MS-46(63.54%)、 MS-2 (63.13%)、 MS-31 (62.29%)、MS-52(62.08%)、MS-80 (61.88%)、MS-55 (61.46%)和MS-33(61.25%),后续对部分内生细菌的抑菌谱进行了测定(表1)。
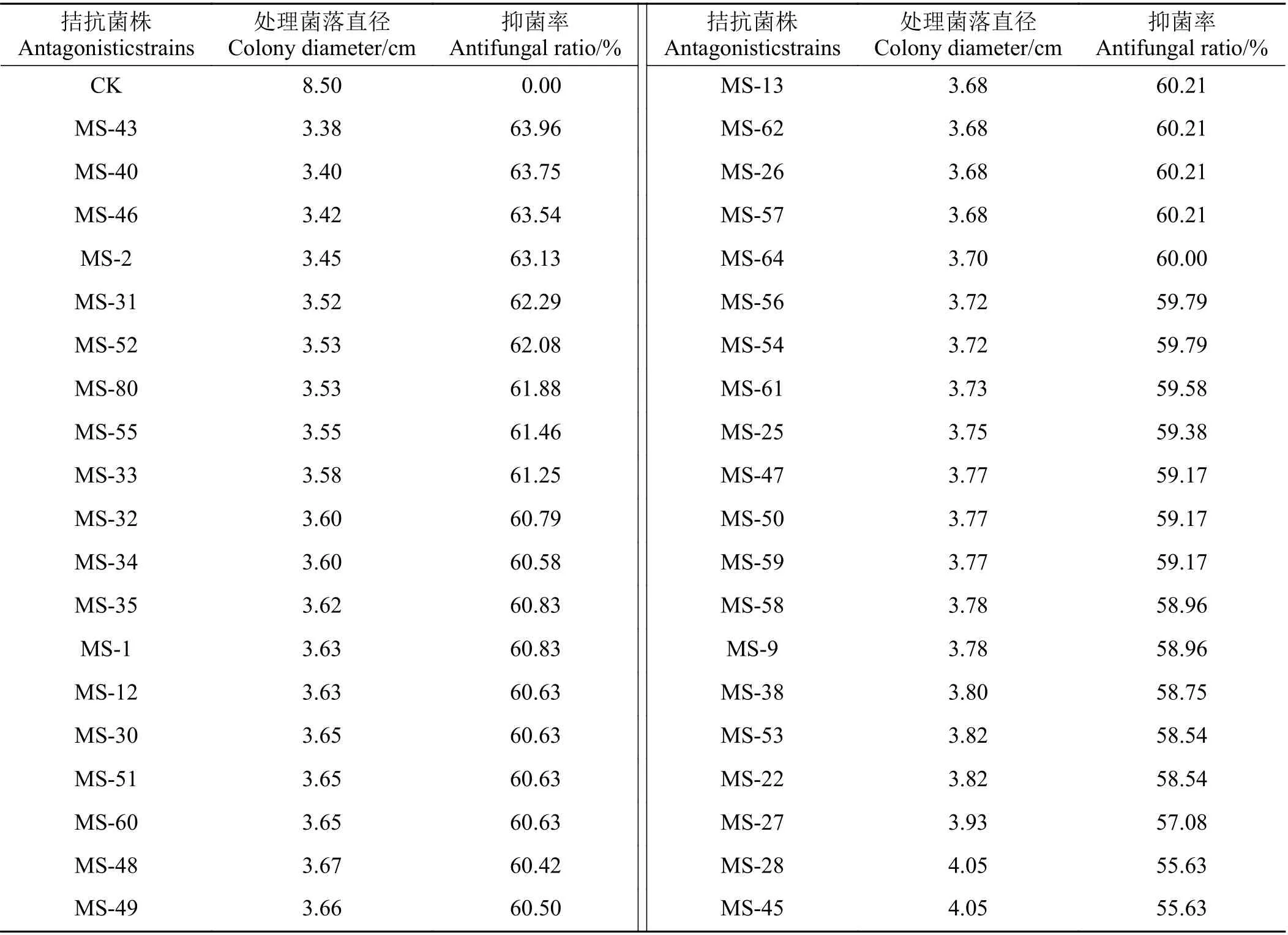
表1 部分内生细菌对厚垣镰孢菌的抑菌能力测定Table 1 Inhibitory activity of endophytic bacteria against Fusarium chlamydosporum
经抑菌谱测定,初步筛选出的5 株苜蓿内生细菌对6 株植物病原真菌有不同的拮抗作用,由图2可明显看出,6 种病原菌与5 株内生细菌对峙培养时,抑菌带宽度有明显差异。MS-43 对尖孢镰刀菌(辣椒)的拮抗作用最大,抑菌率达62.50%;MS-43 对茄镰孢菌的抑菌作用最大,抑菌率达60.63%;MS-43对锐顶镰孢菌的抑菌率最高,为70.63%;MS-43 对燕麦镰孢菌的抑菌率最高,为70.42%;MS-43 对黄瓜枯萎病菌的抑制作用最大,抑菌率达65%;MS-43 对马铃薯枯萎病菌的抑制效果较大,抑菌率为67.50%。总体上,MS-43 对6 株植物病原真菌的抑菌率均高于其他4 株内生细菌,其对燕麦镰孢菌的抑菌率最高,对其他5 株病原菌的抑菌率均在60%以上,其次为MS-33,抑菌率均高于60% (表2)。该结果说明内生细菌MS-43 抑菌性强,抑菌谱广,具有开发潜力。

表2 紫花苜蓿优良拮抗内生细菌的抑菌率Table 2 Antibacterial spectra of excellent antagonistic endophytic bacteria in alfalfa%

图2 紫花苜蓿优良拮抗内生细菌对病原菌的抑菌作用Figure 2 Inhibition of fungal pathogens by antagonistic endophytic bacteria in alfalfa
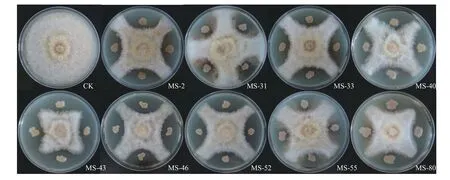
图1 紫花苜蓿内生拮抗细菌对紫花苜蓿根腐病菌的抑菌作用Figure 1 Inhibition of Fusarium chlamydosporum by antagonistic endophytic bacteria
2.3 优良拮抗菌株的生物功能测定
内生细菌MS2、MS31、MS33、MS40、MS43、MS46、MS52、MS55 和MS80 均具有固氮能力,不具解无机磷能力,MS-43 具有分泌IAA 能力(表3)。

表3 紫花苜蓿内生细菌的生物功能测定Table 3 Determination of the biological functions of endophytic bacteria in alfalfa
2.4 优良拮抗菌株对菌丝生长的影响
对峙培养后,挑取与菌株MS-43 交界处的紫花苜蓿根腐病菌菌丝于显微镜下观察,菌株MS-43 对紫花苜蓿根腐病菌菌丝的生长具有显著影响。与对照正常生长的根腐病菌菌丝相比,受菌株MS-43抑制的根腐病菌菌丝易断裂,且有弯曲变形现象(图3)。

图3 对峙培养时菌株MS-43 对紫花苜蓿根腐病菌菌丝生长的影响Figure 3 Inhibitory effect of strain MS-43 on the mycelial morphology of Fusarium chlamydosporum
2.5 优良拮抗菌株的鉴定
2.5.1 培养性状
菌株MS-2 为淡黄色,其他8 菌株均为乳白色。菌株MS-46 菌落直径最小,为2 mm;菌株MS-31 和MS-55 的菌落直径相对较大,为3 mm;其他菌株直径在2.5~2.8 mm。菌株MS-2、MS-40、MS-80 边缘整齐近圆形,其他5 株边缘不整齐呈不规则状,其中MS-46 呈棉絮状;菌株MS-2、MS-43 表面光滑圆润有光泽呈粘稠状,菌株MS-55 表面平整雪花状,其他菌株表面菌粗糙有褶状突起(表4、图4)。

图4 内生细菌培养性状Figure 4 Culture characteristics of endophytic bacteria
2.5.2 形态特征
9 株拮抗内生细菌均为杆状革兰氏阳性菌;MS-2 菌体长达3.09 μm,其次为MS-80,为3.07 μm,MS-43 菌体最短,为1.83 μm;MS-31 和MS-55 菌体较宽,为0.67 μm,MS-2 和MS-33 菌体较窄,为0.50 μm(表4、图5)。

图5 内生细菌的革兰氏染色Figure 5 Gram staining of endophytic bacteria
2.5.3 16SrDNA 基因序列分析
将测序结果与EzBioCloud 数据库中的序列比对,构建系统发育树后发现菌株MS-43 与类芽胞杆菌属的Paenibacillus maysiensis(JN873138.2) 一致性达100%,初步将菌株MS-43 鉴定为类芽孢杆菌(图6)。

图6 内生细菌Ms-43 与类芽孢杆菌菌株之间关系的系统发育树Figure 6 A phylogenetic tree showing relationships between the endophytic bacterium MS-43 and strains of Paenibacillus species
3 讨论与结论
本研究首次从紫花苜蓿茎部、根部分离筛选对紫花苜蓿根腐病菌(F. chlamydosporum)有抑制作用的拮抗内生细菌,结果表明,分离纯化出的9 株内生细菌对紫花苜蓿根腐病菌等多种镰刀菌有明显抑制作用,具有广谱抑菌活性,其中菌株MS-43 对紫花苜蓿根腐病菌的抑制效果最佳,广谱抑菌活性最高,同时具有固氮能力和分泌IAA 的能力,经鉴定属于类芽孢杆菌属。
苜蓿根腐病是重要的土传病害,且病原种类复杂,防治较为困难,已有的防治方法主要为化学防治,但化学防治会造成环境污染等问题,而生物防治紫花苜蓿根腐病可以克服这一缺点。生物防治紫花苜蓿根腐病的相关研究和报道相对较多,胡进玲等[14]报道了两株对紫花苜蓿根腐病(F. solani和F.oxysporum)具有较强抑制效果的芽孢杆菌菌株;曾亮等[15]首次报道了从苜蓿根际土壤中分离得到的对紫花苜蓿根腐病病原菌抑菌率较高的菌株ZY56 和GY37,经鉴定分别为哈茨木霉(Trichoderma harzianum)和康宁木霉(Trichoderma koningii),并进行了盆栽防效试验,结果表明,ZY56 对盆栽苜蓿根腐病的防效达到66.4%,且促进生根效果明显,GY37防治效果为55.1%。本研究在前人研究的基础上首次从紫花苜蓿内生细菌中筛选生物防治紫花苜蓿根腐病菌(厚垣镰孢菌)的优良拮抗菌株MS-43。植物内生细菌种类繁多,在植物不同部位均可分离得到,因其可以诱导植物产生抗病性,并对植物生长有促进作用,因此是植物病害生物防治中重要的微生物资源库,具有重要研究意义。目前已经从多种植物中分离得到内生拮抗细菌,本研究从紫花苜蓿体内共分离获得80 株内生细菌,与陈孝利[16]分离纯化获得的409 株苜蓿内生细菌相比,本研究获得的菌株数量较少,其原因可能与选取的苜蓿品种较单一以及采用不同苗期的苜蓿有关。目前以内生细菌作为生防菌剂已经成为生防菌剂研制开发的热点,如Wen 等[17]从苜蓿根部分离筛选出的枯草芽孢杆菌MB29 可以抑制多种苜蓿根腐病菌的生长,同时对苜蓿的生长有明显的促进作用;刘莎莎等[18]报道,从紫花苜蓿根际土壤中筛选出的芽孢杆菌CYY-6 和CYY-42 对尖孢镰刀菌有拮抗作用,抑菌率分别为49.30%和56.30%。本研究从紫花苜蓿茎、叶内分离得到的菌株MS-43 对紫花苜蓿根腐病菌的抑菌率达63.96%,效果优于CYY-6 和CYY-42。
菌株MS-43 经鉴定属于类芽孢杆菌属,因植物内生细菌中的芽孢杆菌可以产生具有较强抗逆性的芽孢,因此被各领域广泛应用,类芽孢杆菌是一种产芽孢的G+植物非致病性细菌,具有分泌植物激素和提供氮素等功能,可促进植物生长[19],可产生丰富的代谢化合物,尤其是拮抗微生物类的脂肽化合物,其生防机制与脂肽化合物有关[20],受菌株MS-43抑制的根腐病菌菌丝易断裂,且有弯曲变形现象,培养过程中菌株MS-43 可能产生一些抑制苜蓿根腐病菌菌丝生长的代谢产物,或产生营养竞争现象,其具体作用机理需要进一步的研究。
本研究从紫花苜蓿植物体内分离得到的类芽孢杆菌属菌株MS-43 对多种镰刀菌具有较好的抑菌作用,同时有固氮能力和分泌生长素的能力,具有一定的商业开发潜力,也为紫花苜蓿根腐病的生物防治提供了新的微生物资源。
猜你喜欢
今日农业(2022年15期)2022-09-20
今日农业(2021年21期)2021-11-26
现代畜牧科技(2021年9期)2021-10-13
湖北农机化(2020年4期)2020-07-24
东北农业大学学报(2020年3期)2020-05-14
世界农药(2019年4期)2019-12-30
今日农业(2019年11期)2019-08-15
生物技术通报(2019年2期)2019-03-15
四川农业大学学报(2018年5期)2018-11-09
乡村地理(2018年2期)2018-09-19
