
版纳微型猪近交系ZPBP2基因的克隆及生物信息学分析
2021-11-16 05:34查星琴成文敏李海昌潘伟荣霍金龙
家畜生态学报 2021年10期
查星琴,成文敏,李海昌,潘伟荣,霍金龙,黄 英
(1.云南农业大学 动物医学院,云南 昆明 650201;2.云南农业大学 动物科学技术学院,云南 昆明 650201;3.云南农业大学 云南省动物营养与饲料重点实验室,云南 昆明 650201)
哺乳动物精子进入卵子完成受精的生理过程非常精密和复杂,精子首先必须特异性地与卵子的透明带(Zona pellucida,ZP)结合,经历顶体反应,穿越透明带,才能和卵母细胞融合,完成受精[1]。精卵质膜融合是这一生理过程中最为关键的步骤,很多吸附在精子表面的精浆蛋白及雌性生殖道分泌的因子均在精卵质膜融合中发挥重要的调控作用[2-3]。透明带结合蛋白(zona pellucida binding protein,ZPBP)是精子表面能与卵子透明带相结合的蛋白分子,包括透明带结合蛋白1(zona pellucida binding protein 1,ZPBP1)和透明带结合蛋白2(zona pellucida binding protein 2,ZPBP2),参与精子早期结构的形成和精卵融合过程。在大鼠、小鼠、猪、人[4]的精子中研究发现,ZPBP1基因只在人、小鼠和猪的睾丸组织表达,ZPBP2基因除了在人、小鼠和猪的睾丸组织表达,在猪精囊腺和附睾尾均有表达,且在睾丸中表达丰度最高[5]。ZPBP1和ZPBP2基因主要参与精子与卵子的细胞外基质次级互作[6],在精子发生中可能参与精子结构形成[7],对精子顶体形成和形态发生发挥重要作用,可见其在雄性不育的发生中起着十分重要的作用。
版纳微型猪近交系(BMI)是云南农业大学曾养志教授对连续20多代采用全同胞或亲子交配等高度近交方法,经严格选择、精心培育成功的大型哺乳动物近交系。BMI遗传背景清楚,基因高度纯合,实验可重复性高,是动物模型建立、转基因动物生产、生物医学实验及异种器官移植等研究最为理想的实验动物。但BMI雄性动物普遍存在繁殖力低的现象,影响近交系动物的培育进程。因此,研究ZPBP2基因在雄性不育中的作用具有十分重要的意义。本研究使用RT-PCR方法扩增并克隆出BMI的ZPBP2基因,对其进行生物信息学分析,旨在揭示ZPBP2基因的生物学特性,为进一步深入探索BMI 弱精和繁殖力下降的分子机制奠定基础。
1 材料与方法
1.1 主要材料与试剂
采集云南农业大学版纳微型猪近交系重点实验室昆明猪场成年(15月龄) BMI不育公猪睾丸组织,液氮速冻,置于-80 ℃以备RNA提取。
所用试剂Competent Cell Preparation Kit、DL2000、RT-PCR一步法试剂盒、病毒总RNA/DNA提取试剂盒(TaKaRa MiniBEST Viral DNA/RNA Extraction Kit Ver.3.0)、TaKaRa Ex TaqTMTaq酶、逆转录酶XL(AMV)等均购自宝生物工程(大连)有限公司,DNA纯化试剂盒购自全式金生物技术有限公司。其他试剂均为国产分析纯。
1.2 方法
1.2.1 合成引物 参照Genbank公布的猪ZPBP2基因(基因登录号GQ369764和GQ369765)的保守序列,应用 Primer Premier 5.0 和 Oligo 6.0 软件设计特异性引物,由上海生工有限公司合成,引物序列为:5′-CAAAGCAGAAACCCAAGA-3′;5′-CACCTCCACAAGCAAGTA-3′。扩增片断长度约为479 bp。
1.2.2 RNA的提取 称取大约100 mg的BMI睾丸组织样品,按 TaKaRa 公司的RNA Plus说明书步骤提取总RNA,用核酸蛋白测定仪测定其浓度及纯度,稀释成200 ng/μL,取5 μL用1%琼脂糖凝胶电泳检测完整性。
1.2.3 cDNA合成 将检测合格的总RNA样品反转录为cDNA,反转录体系为:在0.2 mL的PCR管中加入Total RNA样品2 μg(10 μL),50 μmol/L oligo(dT)18 1 μL,10 mmol/L dNTP 1 μL,混匀后70 ℃孵育5 min,迅速放在冰上冷却;稍微离心后再加入5×first buffer 4 μL,recombiant RNase inhibitor 1 μL,0.1 mol/L DTT 2 μL,轻微混匀后37 ℃孵育 2 min,室温下加入1 μL的 reverse transcriptase M-MLV(200 u/μL),轻轻吹打混匀,37 ℃孵育10 min,最后42 ℃下将其加热1 h终止反应,产物于-20 ℃保存备用。
1.2.4ZPBP2基因PCR扩增反应体系与程序 PCR反应总体系为25 μL:12.5 μL的Premix Taq,8.5 μL的ddH2O,上下游引物(10 pmol/μL)各1.5 μL,1 μL的cDNA模板(25 ng/μL),轻微混匀。PCR扩增程序为:94 ℃ 5 min;94 ℃ 30 s,60 ℃ 30 s,72 ℃ 30 s,35个循环;72 ℃ 8 min。产物用2%的琼脂糖凝胶电泳检测效果,紫外凝胶成像系统(SYGENE,GeneGenius)照像。
1.2.5ZPBP2基因克隆 参照DNA纯化回收试剂盒(全式金生物技术有限公司)说明书步骤进行PCR产物回收纯化,取一份纯化产物送上海生工有限公司测序,另取一份纯化产物连接入pMD18-T克隆载体,构建10 μL反应体系:目的片段4 μL,pMD18-T克隆载体1 μL,Ligation mix 5 μL。16 ℃连接10 h后转化大肠杆菌DH5α感受态细胞,涂布于LB琼脂固体培养基上,倒置于恒温培养箱(37 ℃)中过夜培养形成单菌落,挑出白色阳性菌落于含有AMP的LB液体培养基中震荡过夜培养(37 ℃,200 r/min),进行菌液PCR鉴定,然后送上海生工有限公司测序。
1.2.6ZPBP2基因序列的生物信息学分析 测序结果用NCBI(National Center for Biotechnology Information)的nucleotide blast软件检验核苷酸的序列;用Generunr软件预测出开放阅读框,并得到相应的氨基酸序列;用在线分析软件ExPASy-PrtPram tool分析ZPBP2基因编码的蛋白质结构;通过NetNGlyc 1.0 Server预测蛋白质N-糖基化位点;通过NetOGlyc 4.0 Server预测蛋白质O-糖基化位点;通过NetPhos 2.0 Server预测蛋白质磷酸化位点;通过SignalP 4.1 Server预测信号肽序列;通过TMpred server预测蛋白质中是否有跨膜结构域;用Lasergene软件分析核苷酸的同源性。
2 结果与分析
2.1 ZPBP2基因扩增结果
ZPBP2引物扩增的特异性片段为479 bp,与预期片段长度相一致(图1)。测序结果与GenBank上猪ZPBP2基因序列比较,可以初步确定扩增序列为BMI的ZPBP2基因序列。
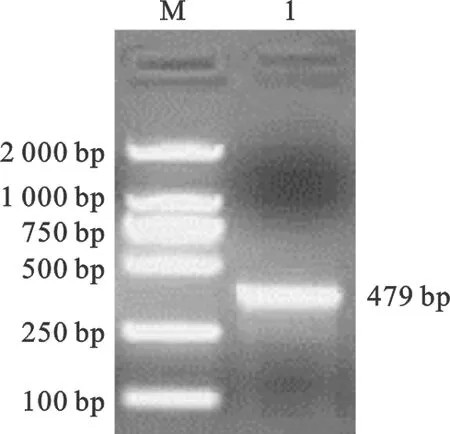
图1 ZPBP2引物扩增的PCR产物2%琼脂糖电泳结果M. DL2000 Marker;1. ZPBP2引物扩增片段Fig.1 2% Agarose gel electrophoresis results ofprimer ZPBP2 PCR productsM. Marker-DL2000;1. PCR product amplified by primers ZPBP2
2.2 ZPBP2基因序列生物信息学分析
用ExPaSy ProtParam在线分析软件预测BMIZPBP2的氨基酸序列,结果显示ZPBP2基因编码蛋白分子量为17.4798 kD,理论等电点为5.63;BMI ZPBP2氨基酸组成上,酸性氨基酸(Asp+Glu)有19个;碱性氨基酸(Lys+Arg)有14个,说明该蛋白为酸性蛋白质,其中以半胱氨酸(Cys)、甘氨酸(Gly)、苯丙氨酸(Phe)、缬氨酸(Val)等含量较高,各占总氨基酸数的7.0%。ZPBP2蛋白质不稳定指数为52.77,属于不稳定蛋白;脂溶指数为66.43,总的亲水性平均系数(Grand average of hydropathicity ,GRAVY)为-0.193,具有弱疏水性。采用SignalP 4.1对ZPBP2蛋白进行跨膜结构预测,结果表明ZPBP2蛋白中没有跨膜信号肽位点(图2);用NetOGly4.0和NetNGly1.0进行预测的结果显示ZPBP2蛋白没有O-糖基化位点,在第32和第68位氨基酸各有一个N-糖基化位点是NGSV和NKTL(图3)。
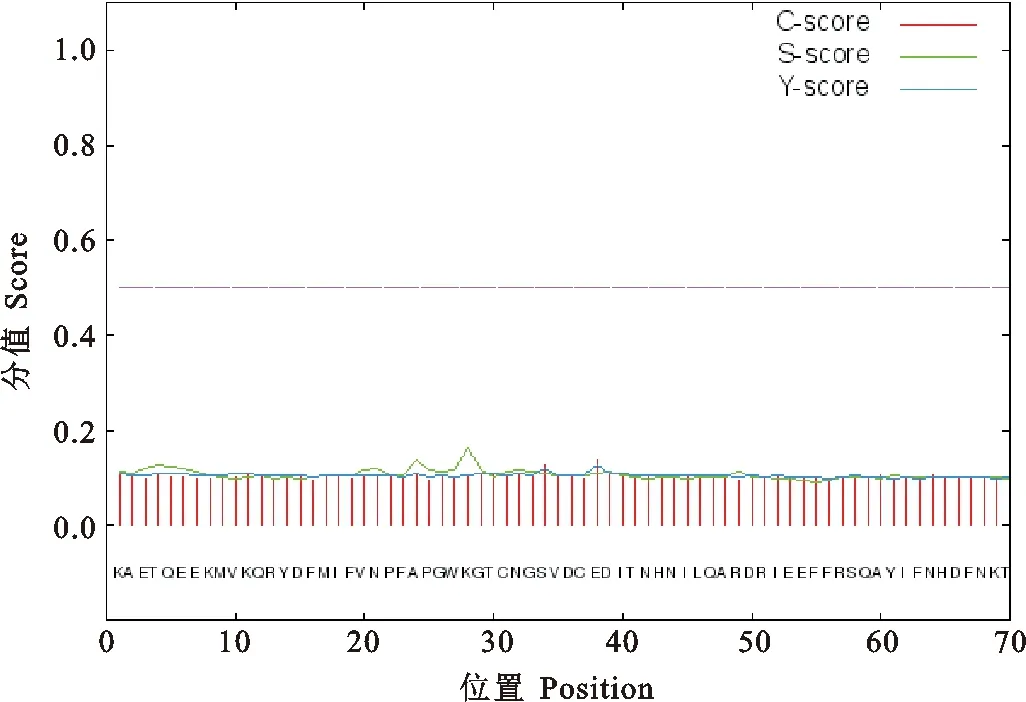
图2 ZPBP2蛋白跨膜信号肽位点预测Fig.2 Predicted transmembrane signalpeptide sites in protein ZPBP2
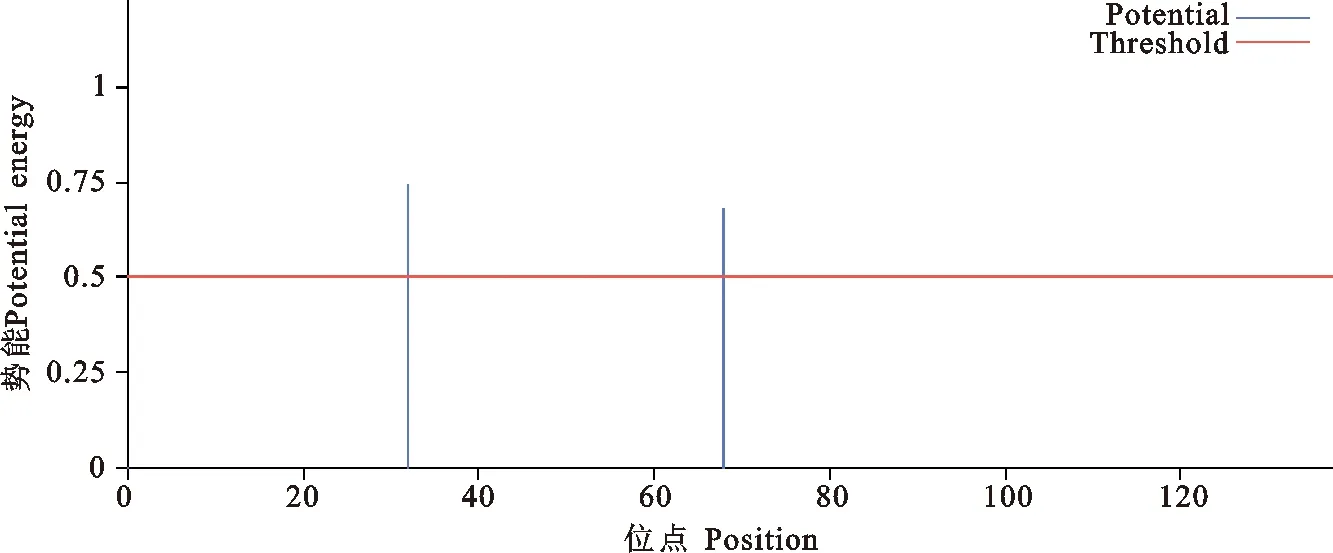
图3 ZPBP2蛋白N-糖基化位点预测Fig.3 Predicted N-glyoosylation sites in protein ZPBP2
利用NetPhos 2.0在线系统预测表明,BMI ZPBP2蛋白有5个磷酸化位点,其中3个丝氨酸(Ser)磷酸化位点(A34、A88、A116)、1个苏氨酸(Thr)磷酸化位点(A4)、1个酪氨酸(Tyr)活性位点(图4)。
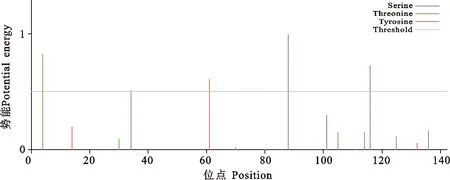
图4 ZPBP2蛋白磷酸化位点推测图Fig.4 Predicted Phosphorylation sites in protein ZPBP2
测序结果经 GenBank 上 Nucleotide Blast检验,结果(图5)显示,在第809位、第854位、第872位分别有A-G、C-T、C-T 3个突变位点;经推导氨基酸序列分析(图6)得出,只有第809位A-G的突变是有义突变,即将丝氨酸突变为甘氨酸。
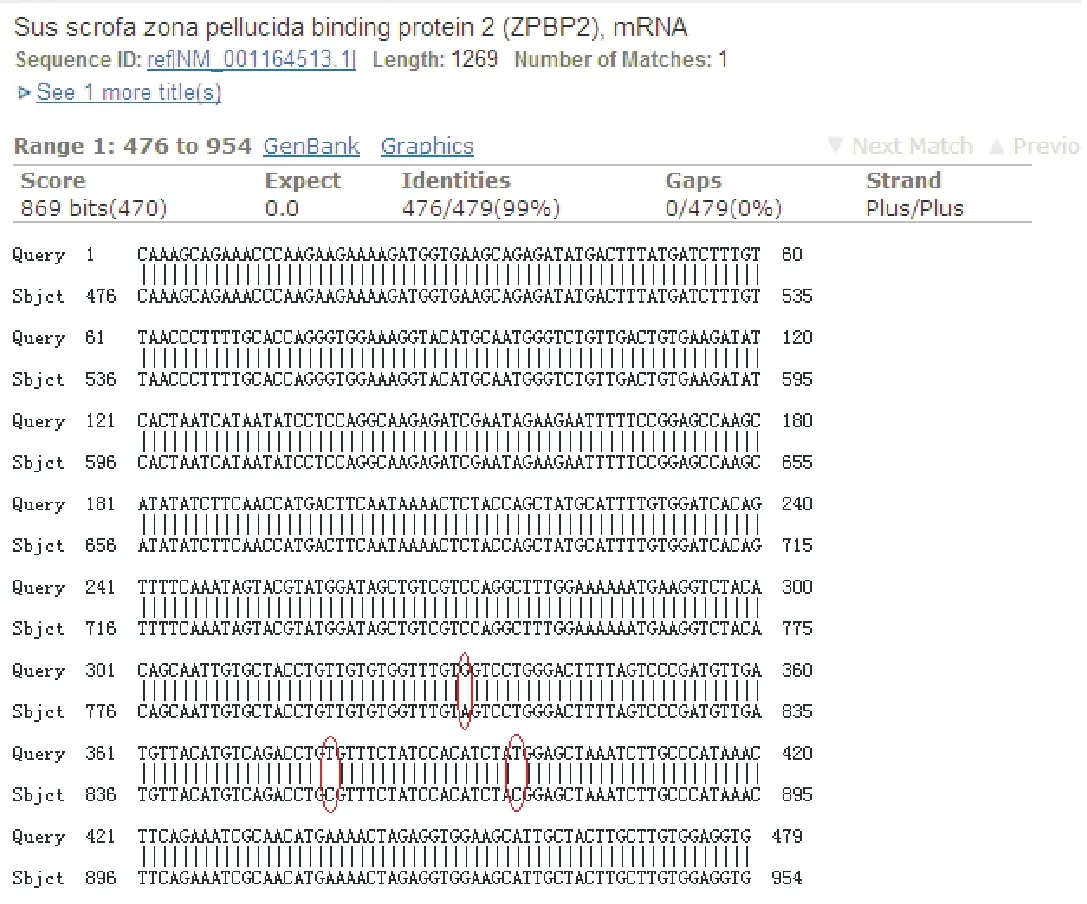
图5 ZPBP2基因核苷酸序列比对Fig.5 Alignment of nucleotide sequences of ZPBP2
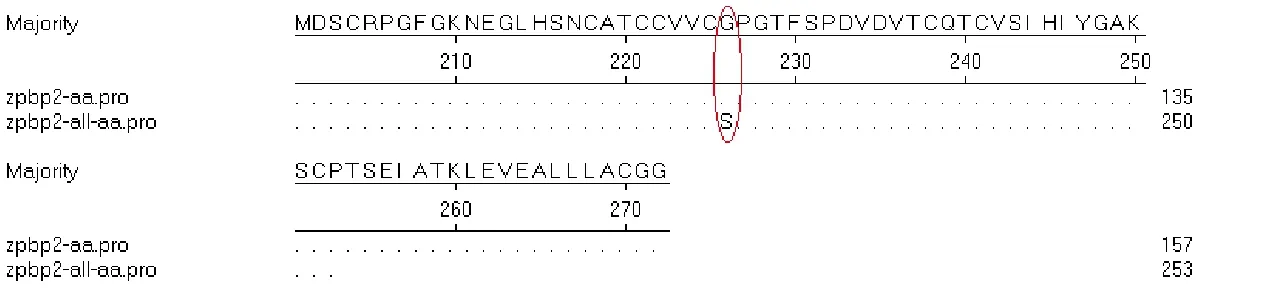
图6 ZPBP2基因推导氨基酸序列比对Fig.6 Alignment of deduced amino acid sequences of ZPBP2
利用SWISS-MODLE软件构建BMI ZPBP2蛋白三级结构模型(图7),通过分析可知预测模型序列与模板序列形成的蛋白质三级结构基本一致,仅扭转角分布不一样,空间构象可能存在细微差异。

图7 ZPBP2的三级结构Fig.7 The tertiary structure of ZPBP2
将BMI ZPBP2 的蛋白质序列与普通猪、牛、绵羊、马、人等6个物种的蛋白质序列导入 MegAlign 计算相似度(图8),相似度分别为99.4% 、90.0% 、88.1% 、87.9% 、85.2%。再将这6条序列导入MEGA7构建系统进化树(图9),结果表明版纳微型猪近交系猪与普通猪遗传距离最近,而与绵羊在进化树中距离最远。可见,版纳微型猪近交系首先与普通猪聚为一类。
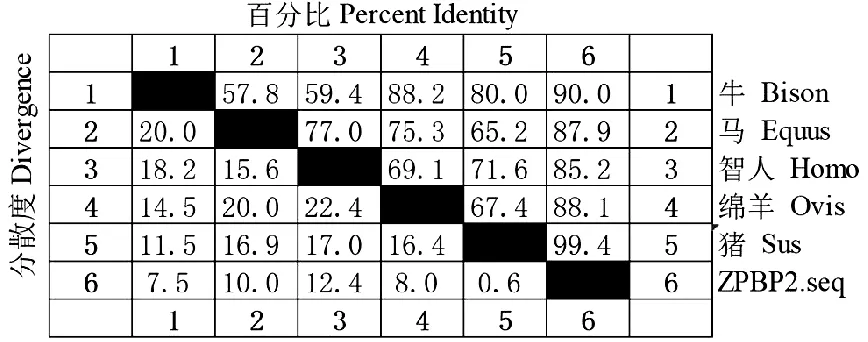
图8 ZPBP2基因核苷酸相似度分析Fig.8 ZPBP2 nucleotide similarity analysis

图9 版纳微型近交系猪ZPBP2基因进化树分析Fig.9 Evolutionary tree for Banna inbred pig ZPBP2 gene
3 讨 论
ZPBP作为一类重要的功能蛋白,是调控精子发生、成熟和精卵融合的重要生物分子,从蛋白质和基因水平研究其生物功能有助于进一步揭示精子发生、成熟以及受精的分子机制,从而为临床上免疫性不育的治疗及新型避孕疫苗的开发开辟新的途径。ZPBP1最早在猪的精子提取物中被鉴定[8],定位于精子的整个顶体,ZPBP2为ZPBP1的旁系同源蛋白,ZPBP2在早期精子细胞中仅限于顶体颗粒中,后期出现在顶体吻脊中。关于ZPBP的生物功能研究一直仅局限于参与精子与卵子透明带的结合,直到2007年Lin等[7]研究发现敲除ZPBP1基因的雄性小鼠表现为无生育力,超微结构研究表明缺失ZPBP1基因会破坏精子细胞与支持细胞的连接;敲除ZPBP2基因的雄性小鼠超微结构显示精子穿越透明带的能力降低并伴有异常的精子顶体膜内陷,表现为生育力低下。这些结果提示,ZPBP不仅参与精卵结合,而且有助于精子的早期结构形成[7]。
利用 RT-PCR技术,本研究扩增了BMIZPBP2基因的部分编码区和部分非编码区系列,成功克隆得到BMIZPBP2基因,序列长度为479 bp,结果表明ZPBP2基因核苷酸序列在第809位A-G、第854位C-T和第872位C-T分别有3个突变位点,但只有第809位A-G的突变是有义突变,即氨基酸由丝氨酸突变为甘氨酸。这种有义突变是否会改变蛋白质的结构、继而影响ZPBP2蛋白的结构和功能还待以后进一步研究才能确定。BMI ZPBP2预测模型序列与模板序列形成的蛋白质三级结构基本一致,空间构象可能存在细微差异。
蛋白质磷酸化在调控蛋白质活力和功能的过程中发挥着重要作用,在推导的氨基酸序列中存在5个磷酸化位点,其中3个丝氨酸(Ser)磷酸化位点(A34、A88、A116)、1个苏氨酸(Thr)磷酸化位点(A4)、1个酪氨酸(Tyr)活性位点,在ZPBP2蛋白参与精子发生过程中,这些氨基酸基序可能发挥了重要的作用。另有研究表明,酪氨酸磷酸化可能参与精子获能和顶体反应[8-10]。在BMI ZPBP2蛋白序列中只存在1个潜在的酪氨酸磷酸化位点,且在哺乳动物ZPBP2蛋白中保守,是否由于酪氨酸磷酸化位点较少,导致对蛋白的修饰和调控较少,才引起BMI公猪繁殖力低下仍需下一步深入研究。
ZPBP2蛋白质属于不稳定蛋白,具有弱疏水性。ZPBP2蛋白没有跨膜信号肽位点和O-糖基化位点,在第32和第68位氨基酸各有一个N-糖基化位点是NGSV和NKTL。
不同物种核苷酸相似度分析表明,BMI的ZPBP2与非近交系猪相似度最高为99.4%,与人相似度较低为85.2%,ZPBP2进化上比较保守,核苷酸物种间相似性较高;系统进化树表明,在亲缘关系上与普通猪同处于一个分支,亲缘关系较近,与绵羊亲缘关系较远,表明ZPBP2基因具有相对的种属特异性,亲缘关系越近,同源性越高。
4 结 论
成功扩增了ZPBP2基因部分编码区序列,序列长度为 479 bp,BMI与普通猪核苷酸相似度较高,同处于一个分支中,亲缘关系较近,与绵羊亲缘关系最远。本研究结果可为进一步研究BMIZPBP2基因的功能和阐明BMI公猪弱精症机理奠定基础。
猜你喜欢
安徽农业大学学报(2022年2期)2022-11-09
中风与神经疾病杂志(2022年9期)2022-10-19
人人健康(2022年18期)2022-10-10
中国生物化学与分子生物学报(2022年8期)2022-09-08
波谱学杂志(2022年1期)2022-03-15
昆明医科大学学报(2022年1期)2022-02-28
现代医学与健康研究电子杂志(2021年21期)2021-12-26
复旦学报(医学版)(2021年5期)2021-10-13
三农资讯半月报(2020年8期)2020-05-13
祝您健康·文摘版(2019年1期)2019-05-14
