
百合LbCAT和LbGPX基因的克隆与表达分析
2021-11-09 02:06迟博文阎俊卉赵子贤文锦芬
西北植物学报 2021年9期
迟博文,刘 立,谢 天,阎俊卉,赵子贤,文锦芬
(昆明理工大学 建筑与城市规划学院,昆明 650500)
过氧化氢酶 (catalases,CAT) 是一种含有亚铁血红素的四聚体催化酶,作为抗氧化酶系统中重要的一员,在过氧化物酶体中将H2O2分解成H2O和O2[1]。CAT在植物逆境、衰老以及生长发育等过程中起着关键作用[2]。研究表明拟南芥中AtCAT1参与消除各种胁迫产生的 H2O2[3]。在桃树叶片也观察到中度干旱胁迫诱导CAT、SOD和GR等抗氧化酶活性呈上升趋势[4]。不仅干旱、高盐、重金属等非生物胁迫都可以影响CAT基因的表达[5-7],而且过表达该基因可以提高植物抗性[8-9]。
在植物抗氧化酶系统中另一个重要的成员是谷胱甘肽过氧化物酶(glutathione peroxidase,GPX)[10],属于非血红素硫醇过氧化物酶,通过催化还原型谷胱甘肽(GSH)转化为氧化型谷胱甘肽(GSSG),使H2O2分解进而保护植物[11]。拟南芥中有8个GPX基因,对不同的非生物性胁迫均有应答[12]。在番茄中表达LePHGPX基因明显增强了其对盐、高温等非生物胁迫的耐受性[13]。
百合(Liliumbrowniivar.viridulum)为百合科(Liliaceae)百合属(Lilium)多年生单子叶草本球根类植物,主要分布在北半球的温带与寒带地区[14]。百合不仅有很好的观赏性,还能作为食材、药材,具有极好的市场前景。中国百合切花种植面积约为933.33 hm2,主要集中在云南,每年增幅在20%以上,昆明是中国百合切花种植面积最大且质量最好的产地之一[15-16]。但切花百合在采后及随后的长距离干运输中均会受到失水胁迫,从而可能导致花的畸形及早衰;因此,对于百合抗性基因资源的深入研究,并且从分子生物技术层面增强其抗逆性具有极重要的意义。本研究从百合中分别克隆了1个CAT基因和1个GPX基因,进行生物信息学分析,并对这2个基因在百合的不同器官、花蕾不同发育时期、PEG模拟干旱处理及独角金内酯(SLs)处理下的表达进行了分析,为进一步研究百合中这2个基因的作用机制提供理论基础。
1 材料和方法
1.1 实验材料和材料处理
实验材料为‘卡瓦纳’红色香水切花百合。在研究百合花蕾生长发育过程中基因的表达情况时,将百合花蕾划分为5个阶段取材。现蕾期:植株刚开始出现花蕾;绿蕾期:植株开始出现大部分花蕾,并且整个花蕾呈现为绿色;红蕾期:植株的所有花蕾都会变成红色,花蕾蓬松、不紧实,花瓣刚刚开始彼此分离但尚未完全开放;开花期:植株的花瓣都已经完全开放,花瓣颜色最鲜艳,具有典型的百合花冠花瓣形状和颜色特点;枯萎期:植株部分花瓣开始枯萎。
(1)取盆花百合的不同器官(根、鳞茎、叶和花)和不同发育期花蕾(其中包括绿蕾期、红蕾期、开花期和枯萎期)的花瓣,取材后立即置于液氮中,然后保存在-80 ℃的超低温冰箱中备用。
(2)实验挑选花茎长而挺直,处于开花期、花蕾直径约8~9 cm的百合切花,分别置于蒸馏水(CK)、10%PEG-6000培养液(PEG)和5 μmol/L SLs+10% PEG-6000培养液(PEG+SLs)中处理。分别在0 d、1 d、3 d、5 d收集花瓣样品,立即液氮速冻并于-80 ℃保存备用。
1.2 方 法
1.2.1 RNA提取和cDNA第一链合成百合RNA使用trizol法提取[17],cDNA链合成用赛默飞世尔公司(RevertAid First Strand cDNA Synthesis Kit)试剂盒,参照说明书的步骤进行。
1.2.2 基因克隆根据前期转录组测序结果和在NCBI上的同源比对,用Primer5.0软件设计引物序列,引物序列见表1。PCR扩增用大连宝生物公司Taq酶(TaKaRa TaqTM)。PCR反应程序:94 ℃预变性5 min,94 ℃变性30 s,退火30 s(退火温度分别为55和53 ℃),72 ℃延伸1 min,35个循环;72 ℃延伸10 min。将PCR产物送生物公司测序。
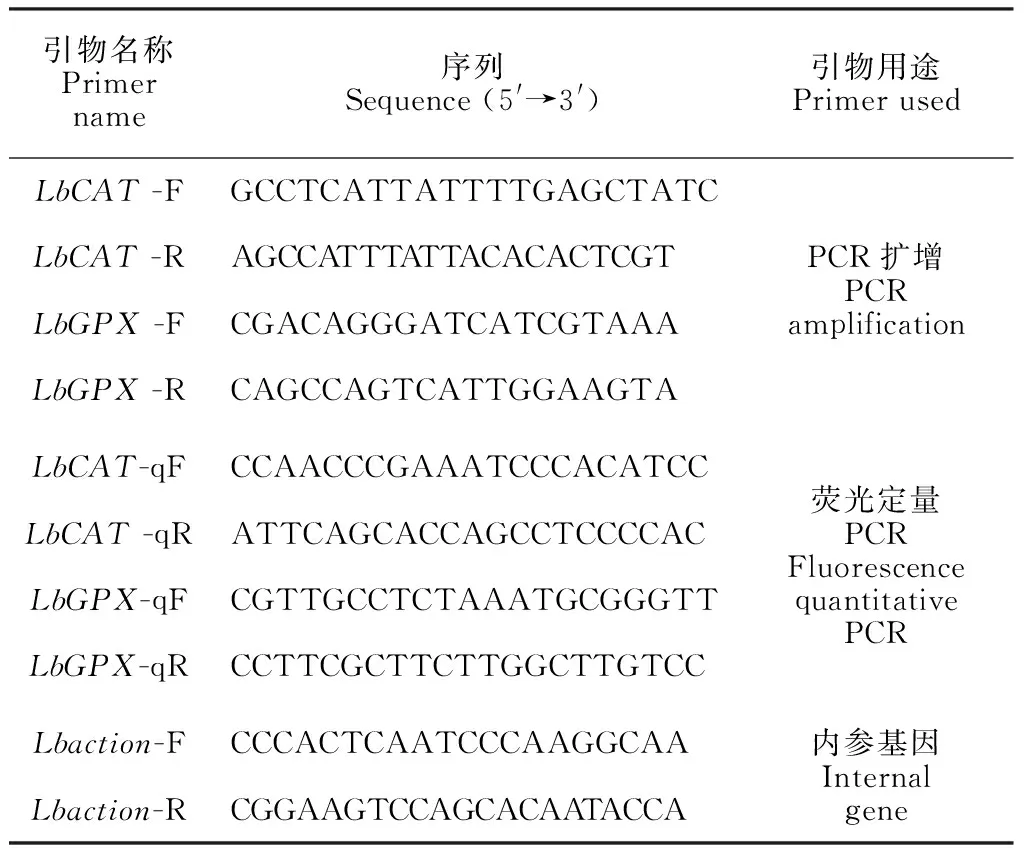
表1 基因克隆和qRT-PCR的引物序列
1.2.3 序列分析用NCBI网站(https://blast.ncbi.nlm.nih.gov)中的ORF Finder(https://blast.ncbi.nlm. nih.gov/orffinder)找出基因的开放式阅读框架(Open reading frame;ORF)和用NCBI网站(https://blast.ncbi.nlm.nih.gov/)的Blastp功能,对序列进行同源性和保守结构域分析。用生物学软件DNAMAN7.0 以及ClustalX2.0.11对核酸进行翻译和多序列比对。用MEGA7.0软件的Neighbor-joining(NJ)计算方法,进行进化树的系统构建与应用分析。
1.2.4 qRT-PCR分析根据克隆结果设计引物,内参基因为Lbaction,引物序列见表1。实时荧光定量PCR使用爱必梦生物科技有限公司的qRT-PCR荧光定量试剂盒(EvaGreen 2X qPCR MasterMix)。反应体系为:10 μL EvaGreen 2× qPCR MasterMix,0.6 μLLbCAT(LbGPX)-qF与0.6 μLLbCAT(LbGPX)-qR,2 μL cDNA模板,加6.8 μL dd H2O至20 μL体系。qRT-PCR反应条件为:95 ℃预变性 10 min,95 ℃变性15 s,退火 60 s(退火温度分别为59和60 ℃),60 ℃延伸 60 s,40个循环。在Bio-Rad CFX96 Optics Module系统上进行反应。每个材料进行3次或多次独立重复。利用2-ΔΔCT算法计算相对表达量,用SPSS进行显著性分析和 SigmaPlot12.5软件作图。
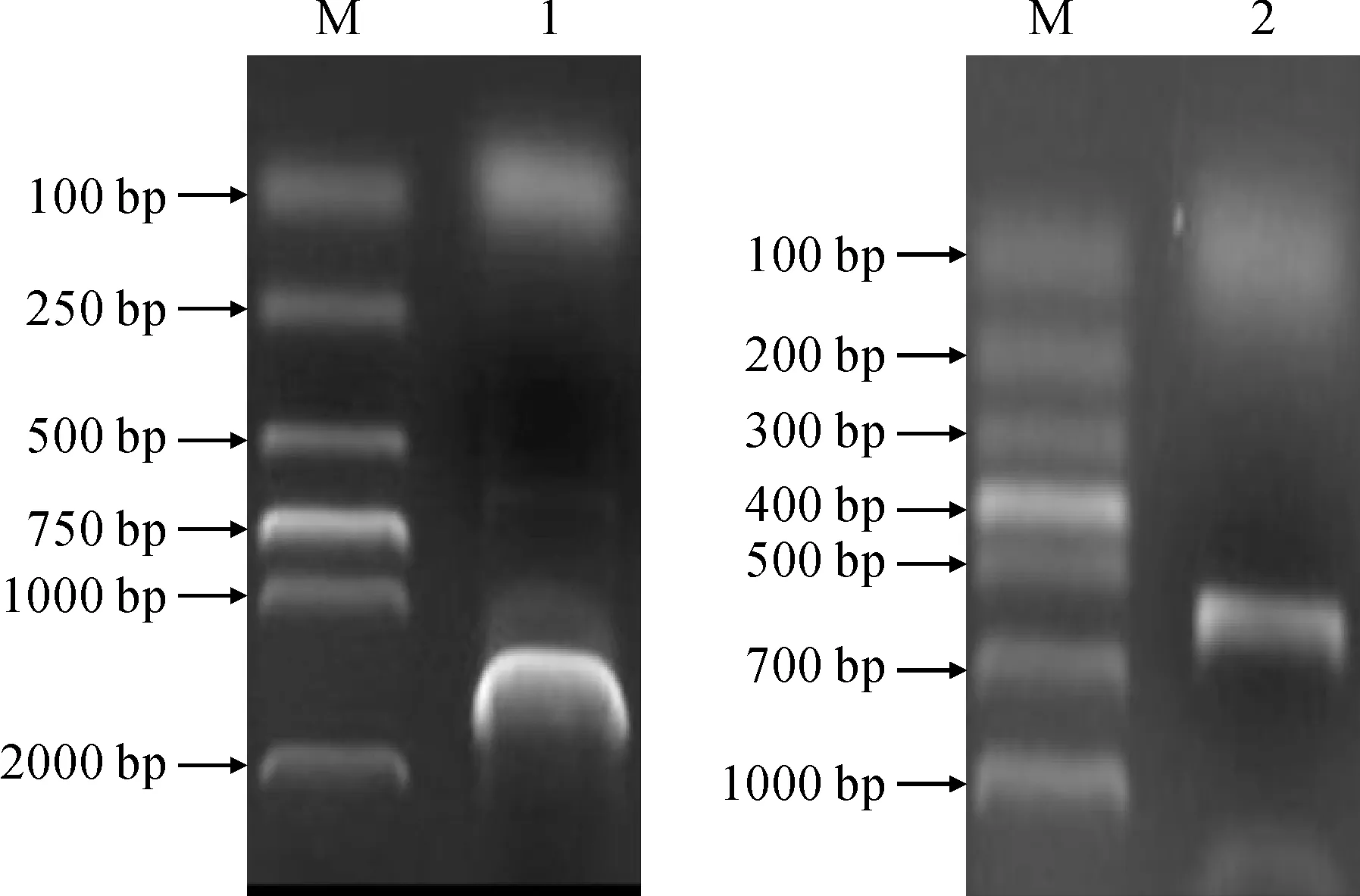
M. DL2000;1. LbCAT;2. LbGPX图1 百合LbCAT和LbGPX基因PCR产物电泳图Fig.1 The electrophoretic map of PCR products of LbCAT and LbGPX in lily
2 结果与分析
2.1 百合LbCAT和LbGPX基因克隆
以百合cDNA为模板,用RT-PCR方法对LbCAT和LbGPX进行特异性引物扩增,分别得到大约1 500和 500 bp的产物(图1)。利用软件ORF finder进行分析LbCAT和LbGPX基因编码的氨基酸序列,其完整的ORF分别为1 479和519 bp,编码的蛋白大小分别为492和172个氨基酸(图2)。

图2 LbCAT(A)和LbGPX(B)全长序列及其推导的氨基酸序列Fig.2 Nucleotides and deduced amino acid sequences of LbCAT (A) and LbGPX (B)
2.2 同源和进化分析
通过NCBI Blast序列比对后发现,LbCAT与岷江百合(Liliumregale)ART33464.1序列相似性最高,为99.19%,与大部分植物序列相似性为77%~99%(图3,A)。LbGPX与油棕(Elaeisguineensis)XP_010938757.1序列相似性最高,为78.61%,与大部分植物序列相似性为67%~78%(图3,B)。LbGPX和LbCAT基因与不同种类植物序列相似性较高说明这2个基因在进化中保守度较高。
用MEGA7.0软件对LbCAT和LbGPX氨基酸序列和其他植物的CAT和GPX进行分析,两种基因都具有显著的种属特征,进化符合植物分类学分类(图4)。整个系统发育树存在两个大分支,其中LbCAT与同科植物的亲缘关系最近,其次是铁皮石斛和小兰屿蝴蝶兰等其他科植物相对较近,与大麦和紫花针茅等的亲缘关系最远(图4,A)。而对LbGPX分析结果显示,整个系统发育进化树存在两个分支,LbGPX与油棕亲缘关系最近,与红梅和水蜜桃等种属关系最远(图4,B)。

A:XP_020092953. 菠萝;XP_020702876. 石斛兰;ART33464. 百合;XP_008791900. 海枣;NP_001306842. 油棕;B:XP_010938757. 油棕;XP_010524011. 醉蝶花;XP_020108868. 菠萝;XP_020267646. 芦笋;XP_020685583. 链状石斛图3 LbCAT(A)和LbGPX(B)分别与不同物种CAT和GPX蛋白氨基酸序列比对A: XP_020092953. Ananas comosus; XP_020702876. Dendrobium catenatum; ART33464. Lilium regale; XP_008791900. Phoenix dactylifera; NP_001306842. Elaeis guineensis; B: XP_010938757. Elaeis guineensis; XP_010524011. Tarenaya hassleriana; XP_020108868. Ananas comosus; XP_020267646. Asparagus officinalis; XP_020685583. Dendrobium catenatumFig.3 Multiple alignment of LbCAT (A) and LbGPX (B) with those of other species
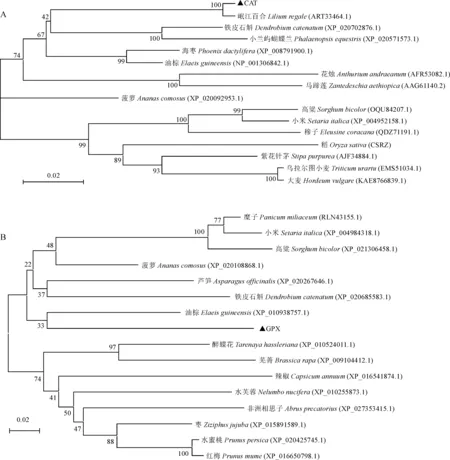
图4 LbCAT(A)和LbGPX(B)蛋白系统进化分析Fig.4 Phylogenetic analysis of LbCAT (A) and LbGPX (B)
2.3 LbCAT和 LbGPX表达分析
2.3.1LbCAT和LbGPX器官和花蕾不同时期的表达分析以Lbactin作为内参,对LbCAT和LbGPX在百合不同器官和花蕾不同发育阶段的表达情况进行qRT-PCR分析。结果表明,LbCAT基因在根、鳞茎、叶和花中都有表达,其中在叶片中的表达最高,根次之。LbGPX在百合的各器官中也均有表达,其中在花中的表达量最高,为鳞茎中表达量的40倍左右(P<0.01)(图5, A)。
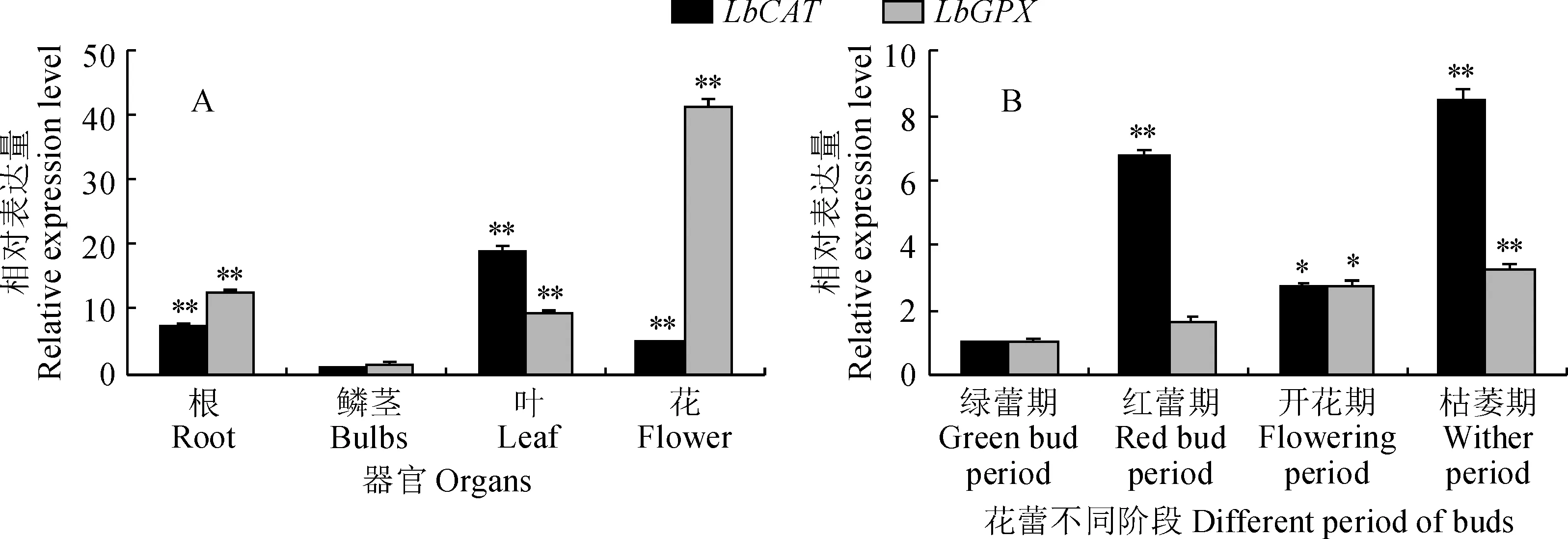
* 、**分别表示同一基因在不同器官相对鳞茎(同一基因在不同花蕾发育阶段相对绿蕾期)的表达差异显著或极显著图5 百合不同组织(A)和花蕾不同发育阶段(B)中LbCAT和LbGPX的表达* and ** stand for the same gene’s expression level in different organ is significantly different relative to the bulb’s (the same gene’s expression level at different bud development stage) at 0.05 and 0.01 levelFig.5 Expression levels of LbCAT and LbGPX in different organs (A) and different bud development stages (B) of Lily
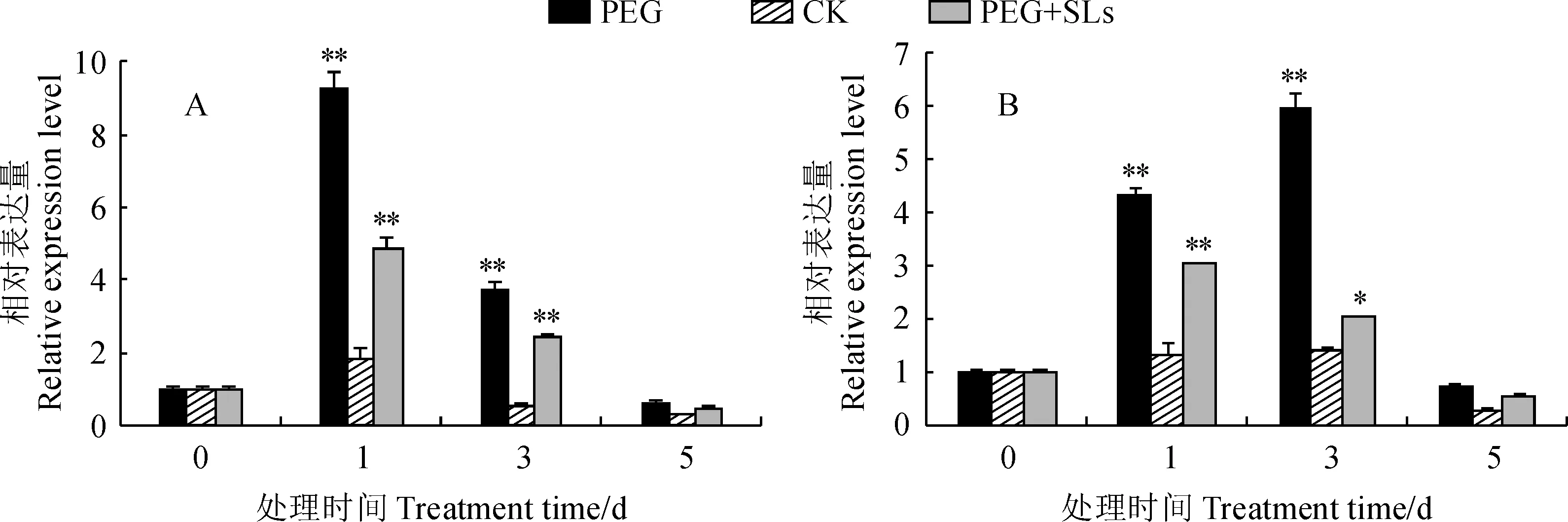
* 、**分别表示同一时期不同处理与对照差异显著或极显著图6 LbCAT (A)和LbGPX (B)在不同处理下的表达* and ** stand for the gene’s expression level in different treatments is significantly different relative to the control’s on the same time at 0.05 and 0.01 levelFig.6 Expression levels of LbCAT (A) and LbGPX (B) under different treatments
在花蕾的不同生长发育阶段(图5, B),LbCAT和LbGPX都有表达,其中LbCAT在枯萎期表达量最高,约为绿蕾期表达量的8倍左右,差异极显著(P<0.01)。LbGPX的表达水平则随着花蕾的发育,总体呈现增长的趋势(图5,B)。
2.3.2 SLs对PEG处理下百合切花LbCAT和LbGPX表达的影响LbCAT和LbGPX基因在不同处理下的表达情况如图6所示。LbCAT表达在第1天最高。PEG处理下LbCAT基因的表达量第1天约为CK的5倍;但SLs+PEG处理的表达量却显著低于PEG处理组,表明SLs处理降低了LbCAT的转录水平(图6,A)。
不同处理下,LbGPX表达量呈先上升后下降的趋势。在PEG处理下,LbGPX的表达量在第3天达到了峰值,约为CK组的4倍;在PEG+ SLs处理下LbGPX的表达量低于PEG处理(图6,B)。
3 讨 论
本研究通过RT-PCR从百合中分别克隆了LbCAT和LbGPX基因,氨基酸序列比对结果表明两种基因在进化中高度保守。qRT-PCR定量分析结果说明,LbCAT和LbGPX在百合不同器官中表达都有特异性,其中分别以在叶片和花中的表达量最高;而在花蕾的不同发育阶段,两个基因表达量均随花蕾的发育而增加。在橡胶中的研究结果表明HbCAT2随着橡胶树的发育其表达量逐渐升高[18],而在香蕉的研究中发现MaGPX在其花中的表达情况与LbGPX一致[19];这种抗氧化酶基因的器官表达差异,可能与不同的器官中活性氧分布的差异有关联[20-22]。
进一步用PEG模拟干旱处理百合切花发现,LbCAT和LbGPX的表达在经过PEG处理后表现出明显的上调。有研究发现水稻叶片组织中的OsCATB基因转录水平能够在受到严重干旱胁迫时升高[23];而在对木薯的研究中也观察到4个木薯栽培种中有3个在干旱诱导下,CAT转录水平上调[24]。
此外,在香蕉、野生近缘种小麦和桑树中也发现GPX基因在干旱胁迫下表达上调[20,25-26]。因此,干旱胁迫诱导CAT和GPX表达上调似乎是植物的普遍反应。
已有文献报道表明,SLs能有效提高植物的抗逆性,如SLs能增强鼠尾草中抗盐胁迫的能力[26],提高羽扇豆幼苗在热胁迫下抗氧化系统和乙二醛酶系统的活性而减轻对种子萌发和光系统Ⅱ功能的不利影响[27];但本研究发现,在转录水平SLs并不能提高LbCAT和LbGPX的表达,这与Parisa Sharifi等在鼠尾草中发现SLs不能提高抗氧化酶活性似乎是一致的[26]。导致这些结果的差异一方面可能是因为物种的不同,另一方面在逆境胁迫下SLs可能在不同的水平调控植物的抗逆性。
本研究结果表明,LbCAT和LbGPX基因在百合中参与了对百合不同器官和花蕾生长发育的调控,以及对水分胁迫的应答。后续对这2个基因的研究主要是通过基因的过表达和沉默进行功能验证,以及克隆启动子及短截实验研究它们的调控位点。百合花蕾是百合全株最具观赏价值的部位,目前关于调控百合抗逆机制的研究较少,本研究为更有效了解百合的抗逆机制和进一步改善百合鲜切花瓶插寿命和培育抗性品种提供了理论基础。
猜你喜欢
新疆农业科学(2022年9期)2022-02-21
现代园艺(2017年23期)2018-01-18
作文与考试·小学低年级版(2017年13期)2017-07-06
上海农业学报(2017年3期)2017-04-10
江苏农业科学(2016年2期)2016-04-11
现代农业(2016年4期)2016-02-28
第二课堂(课外活动版)(2015年5期)2015-10-21
作文与考试·小学低年级版(2015年8期)2015-09-25
农业工程技术·温室园艺(2009年2期)2009-04-17
