
小麦叶绿素酸酯a氧化酶(TaCAO)基因的克隆与表达分析
2021-11-06 03:22樊哲儒张跃强李剑峰
新疆农业科学 2021年9期
王 重,樊哲儒,张跃强,李剑峰,高 新,时 佳
(1.新疆农业科学院核技术生物技术研究所,乌鲁木齐 830091;2.农业部荒漠绿洲作物生理生态与耕作重点实验室,乌鲁木齐 830091;3.新疆农作物生物技术重点实验室,乌鲁木齐 830091)
0 引 言
【研究意义】新疆南疆三地州(喀什地区、和田地区和克孜勒苏柯尔克孜自治州)果麦间作比例占70%左右,形成了南疆三地州特有的林果与种植业复合生产模式[1]。农林复合系统中,林果植物冠层对入射光产生拦截作用,改变了小麦接收到的入射光质量,降低小麦光合有效辐射和产量[2,3]。【前人研究进展】高等植物体内参与光合作用的重要物质是叶绿素,包括叶绿素a、叶绿素b和类胡萝卜素等,其中叶绿素a和叶绿素b主要发挥吸收光能的作用,前者偏向对红光的吸收,而后者更偏向对蓝紫光的吸收。农林复合系统中由于木本植物林冠层的遮蔽作用减少了小麦接收到的入射光,漫射光增加,红光减少,蓝紫光增加,叶片单位叶面积叶绿体体积增大,数量减少,叶绿素含量尤其是叶绿素b的含量增加,叶绿素a/b值下降[4,5],捕捉漫射光中含量较高的较短波长蓝紫光,提高光能利用效率;提高捕光色素复合体LHCP的含量,增加基粒数与基粒片层[6,7];均衡激发光能在光系统PS I和PS II间的分配[7,8];叶绿素b含量增加后,叶绿素a/b值下降,对于光合电子传递具有重要影响;叶绿素a/b值下降,增加了叶绿体对2,6-二氯酚靛酚的还原能力,增强了叶绿体的光合磷酸化活性,使捕光色素维持在较高水平,可吸收更多光能[9,10]。【本研究切入点】叶绿素b是叶绿素a经叶绿素酸酯a氧化酶和叶绿素合成酶共同作用形成的,此途径是叶绿素b形成的惟一途径,叶绿素酸酯a氧化酶(CAO)基因是催化叶绿素b合成的唯一主控基因[11,12]。研究小麦叶绿素酸酯a氧化酶(TaCAO)基因的克隆与表达分析。【拟解决的关键问题】根据GenBank上的已知序列,结合小麦基因组测序信息,以新疆南疆果麦间作主栽小麦品种新冬20号为材料,克隆小麦叶绿素酸酯a氧化酶(CAO)基因ORF,并对其编码的蛋白质序列进行理化性质、进化关系等生物信息学分析,在不同光照强度下通过半定量PCR方法分析TaCAO的表达差异,为农林复合系统中小麦对于弱光环境在叶绿素合成层面的响应奠定基础。
1 材料与方法
1.1 材 料
取新冬20号旗叶迅速置于液氮中,冻存于-80℃冰箱中备用。
Trizol、MightyScript第一链cDNA合成Master Mix、2X 高保真PCR Mix预混液、琼脂糖凝胶回收试剂盒、T4DNA连接酶、TA克隆载体pUCm-T Vector、大肠杆菌(Escherichia coli)菌株DH5α感受态细胞购自上海生工,6×Loading buffer购自TAKARA公司,2×PCR Master Mix购自博迈德公司,GoldViewTM、DL2000购自北京庄盟生物,其余试剂为进口或国产分析纯。PCR引物合成及DNA测序服务均由上海生工提供。
1.2 方 法
1.2.1 RNA提取及反转录
采用Trizol法提取小麦旗叶总RNA,取1 μL进行检测,使用Nanodrop分光光度计检测RNA质量,并跑琼脂糖凝胶电泳琼脂糖凝胶电泳检测RNA纯度。使用MightyScript第一链cDNA合成Master Mix进行反转录,获得的cDNA冻存于-20℃冰箱中备用。
1.2.2CAO基因ORF克隆
根据GenBank上公布的拟南芥CAO基因序列在小麦基因组数据库中进行Blast,筛选出位于3号染色体上的TaCAO基因序列。利用Primer Premier 5.0软件参照序列设计特异性引物:TaCAO-F:GCGTCCATGACCACAGTG;TaCAO-R:TGTTTGCCTATGATCCACTC。
以小麦cDNA为模板进行RT-PCR,将扩增产物在1.0%琼脂糖凝胶进行电泳检测,获得的cDNA为模板进行RT-PCR扩增目的基因,退火温度49~52℃。使用1%琼脂糖凝胶电泳检测PCR产物,回收大小正确条带连接至pUCm-T载体,转化大肠杆菌DH 5α后37℃暗培养20 h。挑取白斑进行PCR验证,阳性克隆送至上海生工测序。
1.2.3 生物信息学
将测序结果通过NCBI数据库(https://www.ncbi.nlm.nih.gov/)的ORF Finder、BLAST、CD-search等工具进行在线比对,确定序列完整编码框、同源序列及保守结构域;使用序列处理在线工具包(http://www.bio-soft.net/sms/index.html)分析TaCAO编码蛋白的氨基酸序列;使用蛋白分析系统ExPASy(http://www.expasy.org/protparam)分析蛋白质基本理化性质[13];使用SoftBerry在线软件(http://linux1.softberry.com/berry.phtml)对蛋白的亚细胞定位进行预测;使用在线工具TMHMM Server v.2.0(http://www.cbs.dtu.dk/services/TMHMM/)分析蛋白质跨膜结构域;使用在线分析工具ProtScale(https://web.expasy.org/protscale/)分析蛋白的疏水性/亲水性[14,15];使用在线软件SOPMA分析蛋白二级结构(https://npsa-prabi.ibcp.fr/NPSA/npsa_sopma.html)[16];使用在线工具Smart(http://smart.embl.de/smart/)分析该蛋白的功能结构域[17,18];通过DNAMAN软件进行同源性比对分析[19],使用MEGA 7.0软件构建进化树[20,21]。
1.2.4 不同光强下TaCAO表达变化
新冬20号生长至三叶一心期,分别置于900、1 800、3 600和7 200 lx光照强度下处理3 h,提取叶片RNA并反转录,以β-actin为内参,进行半定量PCR。
2 结果与分析
2.1 小麦RNA提取与反转录
研究表明,OD260/280=1.94,28S条带最亮,提取RNA质量较好。图1
2.2 TaCAO克隆
研究表明,在1.6 KB左右有明显特异性条带,与已知CAO基因的ORF全长接近,回收目的片段并构建克隆载体进行测序。图2
2.3 TaCAO基因的核酸与蛋白质序列
研究表明,小麦CAO基因开放阅读框的长度为1 653 bp;GC含量为53.4%,编码包含550个氨基酸的蛋白质,含量最高的氨基酸为亮氨酸(11.5%)和丝氨酸(7.8%),半胱氨酸(2.4%)和色氨酸(2.5%)含量最少。表1

表1 TaCAO编码蛋白的氨基酸组成Table 1 Amino acid composition of the protein encoded by TaCAO
2.4 TaCAO编码蛋白的基本理化性质
研究表明,TaCAO编码蛋白的大小约为62.12 KD,原子总数为8 696,理论等电点为7.00,其负电荷残基总数(Asp+Glu)为63个,正电荷残基总数(Arg+Lys)为62个;脂肪指数为83.82;不稳定指数(II)为37.12(不稳定指数<40时,表示稳定),亲水性平均值为-0.309(亲水性平均值<0代表亲水性),小麦CAO为亲水性的稳定蛋白。TaCAO编码蛋白包含一个35个氨基酸残基的叶绿体转运肽,定位于叶绿体中。表2
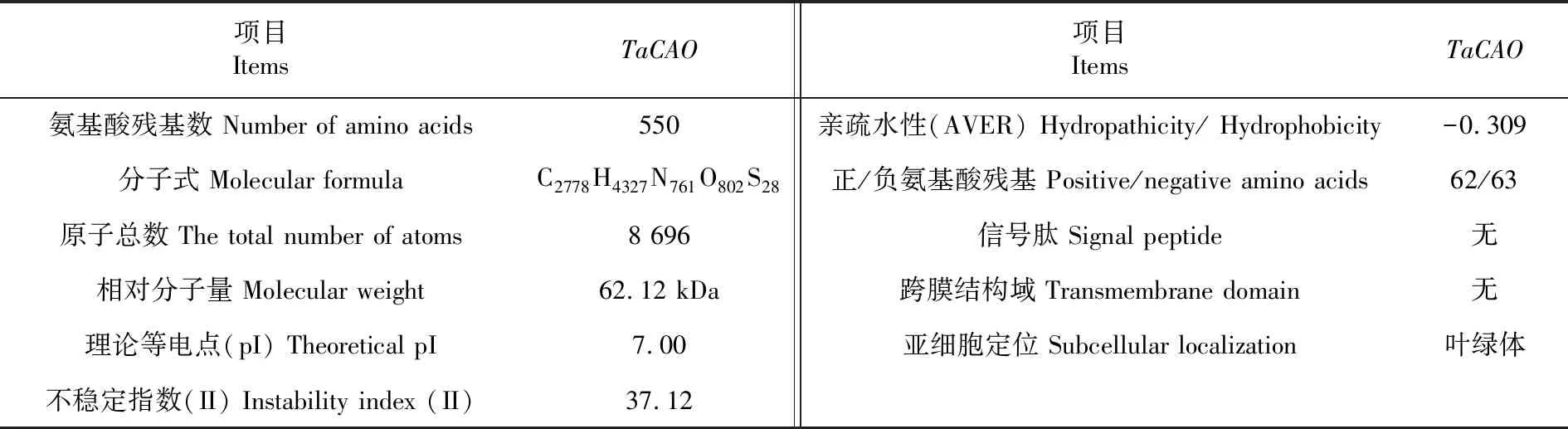
表2 TaCAO编码蛋白的基本理化性质Table 2 The physical and chemical properties analysis of the protein encoded by TaCAO
2.5 TaCAO编码蛋白的二级结构预测
研究表明,该蛋白二级结构主要由无规则卷曲、α-螺旋和延伸链以及β-转角组成,其中无规则卷曲(44.18%)和α-螺旋(37.64%)的含量最高,β-转角(5.09%)含量最低。无规则卷曲均匀分布于整条肽链,而α-螺旋则较为集中的分布于多肽链两端。表3,图3

表3 TaCAO编码蛋白的二级结构比例Table 3 Proportion of secondary structure of the protein encoded by TaCAO
2.6 TaCAO编码蛋白功能结构域的预测
研究表明,该蛋白质包含5个保守结构域,分别是PLN02281、Rieske_RO_Alpha_CAO、Rieske、HcaE和nirD_assim_sml。其中PLN02281和Rieske_RO_Alpha_CAO均是与叶绿素酸酯a氧化酶功能相关的结构域。图4
2.7 TaCAO编码蛋白的同源性
研究表明,TaCAO编码蛋白与大麦(Hordeumvulgare,AF173228.1)同源性最高,为99.45%,二穗短柄草(Brachypodiumdistachyon,XP_003565123.1)次之,同源性为94.73%,与水稻(OryzasativaJaponicaGroup,XP_015614086.1)、矢竹(Pseudosasajaponica,AMH40452.1)、糜子(Panicummiliaceum,RLN22716.1)、高粱(Sorghumbicolor,XP_002459143.1)、玉米(Zeamays,NP_001151343.1)、小米(Setariaitalica,XP_004971349.1)、拟南芥(Arabidopsisthaliana,NP_175088.1)的同源性分别为86.18%、86.18%、84.91%,84.03%、83.67%、83.43%和67.39%。不同植物CAO氨基酸序列具有较高保守性,TaCAO编码蛋白可能与其他植物的CAO具有相同的进化起源。小麦CAO与大麦CAO亲缘关系最近,与糜子和小米亲缘关系较远。图5,图6
2.8 不同光强下TaCAO表达
研究表明,TaCAO相对表达量随着光照强度减弱而逐步增强,呈现递增关系。900 lx光照强度下TaCAO相对表达量最高,7 200 lx光照强度下TaCAO相对表达量最低,弱光环境下TaCAO表达是上调的。图7
3 讨 论
叶绿素酸酯a加氧酶(CAO)是一种Rieske型单加氧酶,能够专一性的识别叶绿素酸酯a,通过对其C7侧链上的甲基(-CH3)进行两步氧化反应使其形成甲酰基(-CHO),转变为叶绿素酸酯b[22];然后再由叶绿素合酶催化在叶绿素酸酯b的D环上加一个植醇合成叶绿素b。叶绿素酸酯a 氧化酶(CAO)在叶绿素a、b的转化和平衡中具有极其重要的作用。
在线分析氨基酸序列,在试验中克隆得到的TaCAO编码蛋白含有一个35个氨基酸残基的叶绿体转运肽,还存在Rieske_RO_Alpha_CAO和Rieske铁硫配位中心2个保守结构域,与姜可以[23]、Yamasato[24]等的研究结果一致。在该蛋白序列中还存在苯丙氨酸双加氧酶和亚硝酸还原酶2个活性位点,可能在叶绿素b合成过程中参与了表达调控作用。
对获得的氨基酸序列进行同源性分析,禾本科植物中的CAO相似性全都在80%以上,大麦、二穗短柄草等亲缘关系较近的植物相似性更是达到了90%以上,进化分析结果表明,小麦CAO与大麦CAO的亲缘关系最近,同时拟南芥CAO与高粱、玉米CAO的亲缘关系也较近,在高等植物中CAO可能存在相同的进化起源,也具有高度的保守性。
通过半定量PCR对TaCAO在不同光照强度下的相对表达量进行分析,弱光环境下该基因表达上调,其相对表达量与光照强度呈现出反比关系。TaCAO表达量的提高能够增加小麦叶片中叶绿素b含量,增强对于较短波长的蓝紫光的吸收能力,使小麦在弱光环境下仍然能够保持较高水平的光合效率。
4 结 论
由小麦品种新冬20号中克隆了小麦叶绿素酸酯a氧化酶基因TaCAO,该基因ORF长度为1 653 bp,编码由550个氨基酸残基组成的蛋白,大小为62.12 kD,含有叶绿素转运肽和Rieske_RO_Alpha_CAO结构域,还存在一个Rieske铁硫配位中心和铁结合位点,具有完整的CAO活性。该蛋白与大麦的亲缘关系最近,并具有较高的保守性。该基因的表达与光照强度呈现反比关系,在小麦适应弱光环境中可能发挥了保持光合效率的作用。
猜你喜欢
环球时报(2022-09-20)2022-09-20
湖北农业科学(2022年11期)2022-07-18
实用肿瘤学杂志(2020年4期)2020-12-08
阅读(科学探秘)(2020年8期)2020-11-06
绿色科技(2019年2期)2019-05-21
女性天地(2016年10期)2017-04-25
小资CHIC!ELEGANCE(2015年14期)2015-09-23
小资CHIC!ELEGANCE(2015年15期)2015-09-01
时代英语·高二(2015年2期)2015-05-18
食品工业科技(2014年6期)2014-05-10
