
物理和化学处理在翅果油树种子萌发中对酶活性的影响
2021-10-30 12:53翟静娟
天津农业科学 2021年10期
翟静娟

摘 要:为探讨不同物理和化学处理对翅果油树种子萌发的影响,本研究对翅果油树种子分别给予变温、激光、超声波、H2O2和PEG处理,以无处理种子作为对照,测定种子萌发过程(3,6,9,12,15,18 d)中的相关酶活性以及丙二醛含量的变化。结果表明:不同处理对翅果油树种子萌发过程中酶活性和丙二醛含量的影响存在差異,主要表现在相关指标随着萌发时间的波动变化方面。其中,超氧化物歧化酶活性变异系数表现为激光>变温>H2O2>超声波>对照>PEG;过氧化物酶活性变异系数表现为H2O2>激光>超声波>变温>对照>PEG;过氧化氢酶活性变异系数表现为超声波>H2O2>激光>PEG>变温>对照;抗坏血酸过氧化物酶活性变异系数表现为变温>H2O2>超声波>对照>激光>PEG;异柠檬酸裂解酶活性变异系数表现为对照>激光>超声波>PEG>H2O2>变温;MDA含量变异系数表现为激光>变温>超声波>PEG>H2O2>对照。综上,不同物理和化学处理会影响翅果油树种子萌发过程中相关酶活性及丙二醛含量的变化,但其与翅果油树种子萌发的关系仍需要进一步研究。
关键词:翅果油树;种子;萌发;酶活性
中图分类号:Q945.34 文献标识码:A DOI 编码:10.3969/j.issn.1006-6500.2021.10.003
Effects of Physical and Chemical Interventions for Enzyme Activities during Seed Germination of Elaeagnus mollis Diels
ZHAI Jingjuan
(Shuozhou Vocational Technology College, Shuozhou, Shanxi 036002, China)
Abstract: To analyze the effects of different physical and chemical interventions during the seed germination of Elaeagnus mollis Diels, seeds of Elaeagnus mollis Diels were dealt with variable temperature, laser, ultrasonic, H2O2, and PEG respectively, no interventions seeds were used for as control. The related enzymes activities and malonaldehyde content were measured during the seed germination (3, 6, 9, 12, 15, 18 d). The results showed that the effects on enzyme activity and malondialdehyde content were various among different interventions during the seed germination of Elaeagnus mollis Diels, which mainly involved the fluctuation of related indexes following with the germination time. The variable coefficient of superoxide dismutase was manifested as laser>variable temperature>H2O2>ultrasonic>control>PEG; variable coefficient of peroxidase was manifested as H2O2>laser > ultrasonic > variable temperature>control>PEG; variable coefficient of catalase was manifested as ultrasonic>H2O2>laser>PEG>variable temperature>control; variable coefficient of ascorbate peroxidase was manifested as variable temperature>H2O2>ultrasonic>control>laser>PEG; variable coefficient of isocitrate lyase was manifested as control>laser>ultrasonic>PEG>H2O2>variable temperature; variable coefficient of MDA was manifested as laser>variable temperature>ultrasonic>PEG>H2O2>control. To sum up, different physical and chemical interventions may impact the related enzymes activities and malonaldehyde content during the seed germination of Elaeagnus mollis Diels, but further studies are needed to investigate the relationship between these interventions and seed germination of Elaeagnus mollis Diels.
Key words: Elaeagnus mollis Diels; seed; germination; enzymatic activity
翅果油树(Elaeagnus mollis Diels)是我国特有的一种乔木状灌丛或小乔木,属于国家Ⅱ级珍稀濒危的保护植物[1-2]。翅果油树具有一定的耐寒性和耐旱性,主要分布于山西省境内,在陕西秦岭等地也有散在分布[3-4]。翅果油树具有非常高的经济价值。翅果仁富含多种蛋白质、维生素、脂肪酸等,能够降低胆固醇、减少动脉粥样硬化等[5]。翅果油树叶中含有的总生物碱具有一定的抗氧化作用,在抗衰老方面具有重要价值[6]。权洪峰等[7]研究表明,翅果油树种子具有一定的抗炎和神经保护活性。但是,翅果油树的种子具有坚硬的外壳,透水性较差,导致翅果油树种子发芽率较低,不利于翅果油树的大规模种植[8]。翅果油树种子的萌发过程受多种因素影响。目前,已有一些研究分析了He-Ne激光、低温、聚乙二醇等对翅果油树种子萌发的作用[9-11],但仅限于评估单一或少数物理、化学处理对翅果油树种子萌发的影响。本研究通过比较多种物理和化学处理对翅果油树种子萌发过程中相关酶活性和丙二醛含量的影响,为进一步研究相关指标与翅果油树种子萌发的关系提供重要的参考依据。
1 材料和方法
1.1 种子来源
山西省临汾市翼城县位于山西省西南部的中条山和太岳山之间,经纬度为(111.72°E, 35.74°N),地势以山区和丘陵为主,是翅果油树的主要生长地之一[12-13]。本研究所使用的翅果油树种子均来自于该地区。
1.2 翅果油树种子的处理
1.2.1 物理处理 变温处理:将带有中果皮的翅果油树种子在-18 ℃的冰箱中冷冻26 h,再经4 ℃冰箱中冷藏168 h(7 d)后,将种子在50 ℃温水中浸泡30 min[14-15]。
激光处理:使用波长为632.8 nm、光斑直径为2 mm的He-Ne激光照射带有中果皮的翅果油树种子7 min[16]。
超声波处理:将带有中果皮的翅果油树种子先用温水浸泡2 h,然后用功率为50 W、频率为40 kHz的超声波对种子进行处理6 min[17]。
1.2.2 化学处理 H2O2处理:先将带有中果皮的翅果油树种子放入清水中浸泡6 h,然后取出,清水冲洗2次,再放入1.5%的过氧化氢溶液浸种2 h[18]。
PEG处理:先将带有中果皮的翅果油树种子在20%的PEG-6 000溶液中浸泡12 h,取出后用蒸馏水清洗数次,用滤纸将水吸干[19]。
上述每种物理和化学处理分别使用至少100颗翅果油树种子,以保证实验室检测所需样本。上述处理完成后,用流动水连续冲泡24 h以软化果皮。然后,手工去掉中果皮。
1.2.3 对照 将未经上述物理和化学处理的、带有中果皮的100颗翅果油树种子用流动水连续冲泡24 h之后,手工去除中果皮,作为对照组。
1.3 试验方法及指标测定
将对照组种子与各处理组种子分别播种于事先准备好的装有原生壤土与细纱混合(混合比例为1∶1)、深11 cm、内径19 cm的花盆内,每盆10颗种子,每个处理组种植10盆,盆中装土2 kg,放置于温室(室温25 ℃,光照12 h),使其萌发。在栽种的第3,6,9,12,15,18 d,分别取样1次。取样时,用小铲子从每盆中随机挖出1颗,清洗去除泥土后,用于实验室检测。采用《种子生物学研究指南》中的方法测量种子萌发过程中相关酶的活性[20]。测量指标包括超氧化物歧化酶(SOD)活性、抗坏血酸过氧化物酶(APX)活性、过氧化物酶(POD)活性、丙二醛(MDA)含量、异柠檬酸裂解酶(ICL)活性、过氧化氢酶(CAT)活性[21-25]。
2 结果与分析
2.1 物理和化学处理对翅果油树种子萌发过程中SOD活性的影响
图1所示,翅果油树种子的超氧化物歧化酶活性随着萌发天数的延长呈现升-降-升-降的双峰变化趋势,其中PEG处理与对照趋势一致,均于9,15 d时达到双峰值而于12 d时为低谷值,变温、超声波和H2O2处理趋势一致,均于6,15 d时达到双峰值而于9 d时达到低谷值,激光处理趋势则在9,15 d时达到双峰值而在12 d时达到低谷值。不同处理的超氧化物歧化酶活性变异系数表现为激光(56.49%)>变温(54.11%)>H2O2(51.92%)>超声波(46.10%)>对照(38.50%)>PEG(38.35%)。
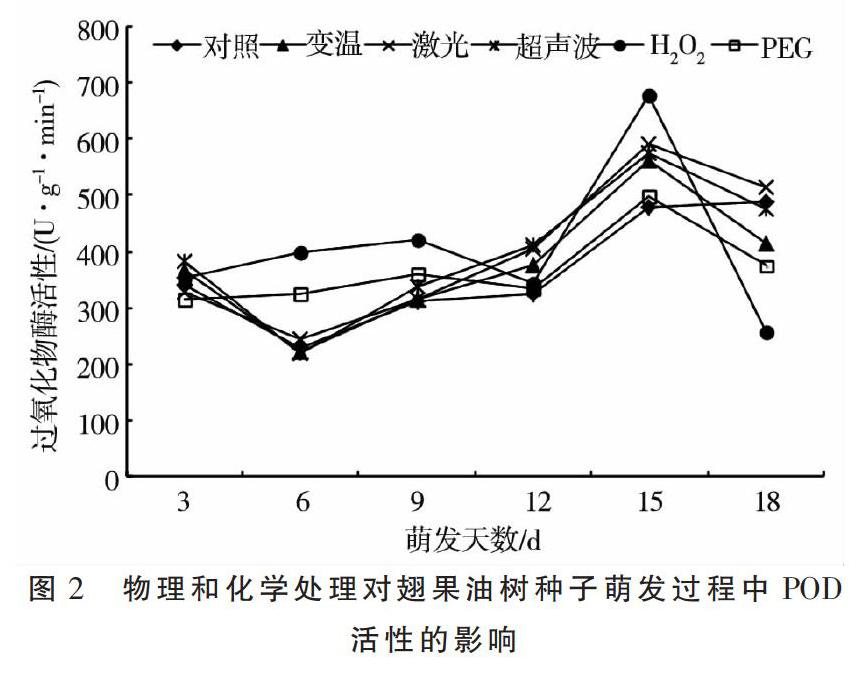
2.2 物理和化学处理对翅果油树种子萌发过程中POD活性的影响
图2所示,随着萌发天数的延长,翅果油树种子的过氧化物酶活性在对照组呈先降低至6 d时达到最低值后波动上升的趋势,在变温、激光和超声波处理呈先降低至6 d时达到最低值后上升至15 d时达到最高值后再下降的趋势,在PEG和H2O2处理呈双峰变化趋势,分别于9,15 d达到峰值且于12 d时为低谷值。不同处理的过氧化物酶活性变异系数表现为H2O2(35.08%)>激光(32.74%)>超声波(30.28%)>变温(29.88%)>对照(28.04%)>PEG(18.30%)。
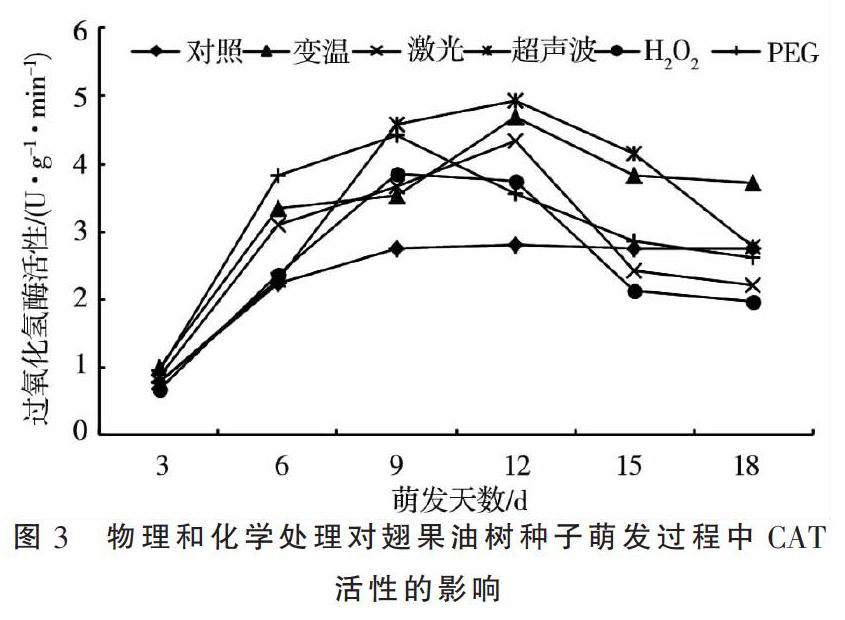
2.3 物理和化學处理对翅果油树种子萌发过程中CAT活性的影响
图3所示,随着萌发天数的延长,翅果油树种子的过氧化氢酶活性在对照组基本呈现3~6 d快速升高后区域平稳的趋势,而在变温、激光、超声波、H2O2和PEG处理基本呈现先升后降的变化趋势,分别于12,12,12,9,9 d时达到最大值。不同处理的过氧化氢酶活性变异系数表现为超声波(48.99%)>H2O2(48.48%)>激光(44.32%)>PEG(40.02%)>变温(37.22%)>对照(33.66%)。
2.4 物理和化学处理对翅果油树种子萌发过程中APX活性的影响
图4所示,各处理翅果油树种子的抗坏血酸过氧化物酶活性均于第6天达到最低值,之后随着萌发天数的延长,其在对照组及变温和H2O2处理呈双峰变化趋势,分别于9,15 d达到峰值,在激光和PEG处理呈升高-降低-升高的趋势,均于12 d达到最大值,在超声波处理呈单峰变化趋势,与9 d达到峰值。不同处理的抗坏血酸过氧化物酶活性变异系数表现为变温(88.32%)>H2O2(79.99%)>超声波(59.87%)>对照(57.83%)>激光(55.47%)>PEG(44.15%)。
2.5 物理和化学处理对翅果油树种子萌发过程中ICL活性的影响
图5所示,随着萌发天数的延长,翅果油树种子的异柠檬酸裂解酶活性在对照组及激光、H2O2和PEG处理均呈现上升(3~9 d)-下降(9~12 d)-上升(12~15 d)-下降(15~18 d)的双峰变化趋势,变温和超声波处理则呈先上升后下降的单峰趋势,其峰值出现在15 d,且所有处理的最大值均出现在15 d。不同处理的异柠檬酸裂解酶活性变异系数表现为对照(44.49%)>激光(44.48%)>超声波(42.81%)>PEG(42.09%)>H2O2(40.51%)>变温(40.11%)。
2.6 物理和化学处理对翅果油树种子萌发过程中MDA含量的影响
图6所示,随着萌发天数的延长,翅果油树种子的MDA含量在對照组及变温和超声波处理呈先降后升的变化趋势,在H2O2处理呈降-升-降-升的趋势,在激光和PEG处理呈升-降-升的趋势。不同处理的MDA含量变异系数表现为激光(143.68%)>变温(120.26%)>超声波(117.91%)>PEG(114.75%)>H2O2(94.60%)>对照(80.62%)。
3 结论与讨论
在自然状态下,翅果油树种子的发芽率较低[26],其萌发过程受多种因素影响。促进种子萌发的方法有很多,如变温[15,27]、激光[16,28]、超声波[28-29]、H2O2 [30]、PEG[19, 31-32]等。宋兆伟等[27]研究发现变温处理可以促进沙芥种子萌发和幼苗生长;郑庆钟等[15]研究发现低于和高于零度变温处理组合可以有效提高沙生针茅种子发芽率。He-Ne激光常被用于种子萌发、增产等,其作用机理可能与种子吸收光能、电磁等有关[28],也可能与萌发过程中SOD和CAT活性有关[16]。超声波作为常用的种子预处理方法之一,可能与产生高温、高压等有关[28],其常被用于有坚硬外壳的种子如油松种子[29]的萌发。H2O2参与种子萌发过程中的多种化学反应,余燕等[30]研究发现H2O2浸种可以促进低温环境下花生种子的萌发。PEG是一种高渗溶液调节剂,具有提高种子活力的作用,常被用于农作物种子萌发,例如:玉米、棉花等[31-32]。本研究着重分析了上述相关处理对翅果油树种子萌发过程中各种酶活性的影响,与对照相比,除PEG外的其他处理均能使翅果油树种子在萌发过程中的超氧化物歧化酶和过氧化物酶活性出现较大幅度的波动,所有处理均能使翅果油树种子在萌发过程中的过氧化氢酶活性和MDA含量出现较大幅度的波动但异柠檬酸裂解酶活性却呈较小幅度的波动,变温、H2O2和超声波处理均能使翅果油树种子在萌发过程中的抗坏血酸过氧化物酶活性出现较大幅度的波动。但相关酶活性或MDA含量与翅果油树种子萌发之间的关系尚有待于进一步开展研究。
参考文献:
[1] 卫晶, 毕润成, 苗艳明. 翅果油树叶面积和叶重的预测模型[J]. 植物分类与资源学报, 2014, 36(4): 497-504.
[2] 鲁甲龙, 张静, 赵红红, 等. 野生和种植翅果油树中脂肪酸的分布[J]. 山西大学学报(自然科学版), 2014, 37(2): 285-288.
[3] 朱志敏, 李艳, 吕金枝, 等. 翅果油树及其群落中7种优势木本种类的叶性特征比较[J]. 植物资源与环境学报, 2017, 26(1): 107-109.
[4] 张文香, 李娟, 张静. 翅果仁食品安全地方标准制定可行性及指标体系的研究[J]. 中国药物与临床, 2020, 20(22): 3840-3841.
[5] 冯笑笑, 李娟, 陈侨侨, 等. 翅果油树种仁蛋白氨基酸组成分析及营养价值评价[J]. 食品科学, 2016, 37(22): 160-165.
[6] 邵芬娟, 闫桂琴, 张直峰, 等. 翅果油树叶片中总生物碱抗氧化活性研究[J]. 西北植物学报, 2008, 28(7): 1339-1342.
[7] 权洪峰, 杨婷, 彭晓东. 翅果油树种子CO2超临界萃取物抗炎及神经保护活性初步研究[J]. 宁夏医学杂志, 2017, 39(11): 980-982.
[8] 郭伟略. 山西濒危植物翅果油资源现状及保护利用对策[J]. 安徽农学通报, 2014, 20(14): 77-78.
[9] 王文君, 翟晓巧, 周玲. 翅果油树种子萌发过程中生理指标的差异性分析[J]. 山西林业科技, 2017, 46(1): 28-30.
[10] 袁丽环, 闫桂琴, 朱志敏. PEG预处理和低温促进翅果油种子萌发的研究[J]. 中国农学通报, 2009, 25(10): 112-115.
[11] 许淑青, 郭春秀, 金红喜, 等. 不同处理对翅果油树种子萌发和育苗的影响[J]. 甘肃科技, 2017, 33(13): 144-146.
[12] 毛婷婷, 毕润成. 山西不同区域翅果油树群落特征的研究[J]. 山西师范大学学报(自然科学版), 2011, 25(3): 98-102.
[13] 张殷波, 刘彦岚, 秦浩, 等. 气候变化条件下山西翅果油树适宜分布区的空间迁移预测[J]. 应用生态学报, 2019, 30(2): 496-502.
[14] 李雄, 尹欣, 杨时海, 等. 温度对高山植物紫花针茅种子萌发特性的影响[J]. 植物分类与资源学报, 2014, 36(6): 698-706.
[15] 郑庆钟, 李发明, 朱淑娟, 等. 不同处理对沙生针茅种子萌发的影响[J]. 生态学杂志, 2016, 35(1): 63-71.
[16] 杨利艳, 高晶, 申圣圣, 等. He-Ne激光辐射对翅果油树种子萌发的生理效应[J]. 应用激光, 2018, 38(5): 812-816.
[17] 郑志远, 余生, 黄演福, 等. 植物激素与超声波对横经席种子萌发的影响[J]. 贵州农业科学, 2020, 48(2): 12-14.
[18] 柳文军, 李爱堂, 喻明曦, 等. 不同处理对柴胡种子萌发的影响[J]. 甘肃农业科技, 2018(9): 25-28.
[19] 尹秀, 王俊, 张二豪, 等. PEG-6000浸种处理对甘青青兰种子萌发及幼苗抗旱性的影响[J]. 江苏农业科学, 2020, 48(13): 168-172.
[20] 宋松泉. 种子生物学研究指南[M]. 北京: 科学出版社, 2005.
[21] 王宁, 昌孝涛, 袁美丽, 等. 香豆素对入侵植物节节麦种子萌发及幼苗生长的影响[J]. 浙江农林大学学报, 2019, 36(4): 835-840.
[22] 苏海兰, 周先治, 李希, 等. 七叶一枝花种子萌发不同阶段生理生化变化研究[J]. 中草药, 2017, 48(22): 4755-4763.
[23] 姚澜, 萨如拉, 范富, 等. 化感物质对玉米种子及苗期生长特性的影响[J]. 中国农学通报, 2020, 36(6): 1-4.
[24] 庞春花, 张媛, 李亚妮. 硝酸镧浸种对藜麦种子萌发及盐胁迫下幼苗生长的影响[J]. 中国农业科学, 2019, 52(24): 4484-4492.
[25] 赵利清, 彭向永, 孙振元. PEG胁迫对西伯利亚冰草种子萌发及幼苗生理特性的影響[J]. 种子, 2017, 36(6): 26-29, 34.
[26] 陈惠, 卢英梅, 申峻沛. 不同栽培基质对翅果油树种子萌发的影响及不同种质幼苗生长发育的表型研究[J]. 山西师范大学学报(自然科学版), 2010, 24(3): 63-66.
[27] 宋兆伟, 郝丽珍, 黄振英, 等. 光照和温度对沙芥和斧翅沙芥植物种子萌发的影响[J]. 生态学报, 2010, 30(10): 2562-2568.
[28] 柳旭, 刘娟, 刘倩, 等. 种子预处理的作用机制研究进展[J]. 应用生态学报, 2016, 27(11): 3727-3738.
[29] 胡立洪. 油松种子催芽处理方法与研究[J]. 农业与技术, 2016, 36(4): 187.
[30] 余燕, 张雅婷, 赵雪, 等. H2O2浸种对低温胁迫下花生种子萌发的调控作用[J]. 中国油料作物学报, 2020, 42(5): 860-868.
[31] 赵璞, 温之雨, 董文琦, 等. PEG-6000模拟干旱胁迫对8个玉米种质种子萌发及活力的影响[J]. 种子, 2020, 39(10): 48-52.
[32] 肖爽, 韩雨辰, 号宇然, 等. 聚乙二醇引发对盐胁迫下棉种萌发及生理特性的影响[J]. 核农学报, 2021, 35(1): 202-210.
猜你喜欢
儿童时代·幸福宝宝(2019年9期)2019-10-28
莫愁·家教与成才(2017年7期)2017-07-11
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
安徽农学通报(2017年1期)2017-02-15
湖北农业科学(2015年17期)2015-10-09
分析化学(2015年4期)2015-06-08
