
性别、年龄、血脂和位置影响肠系膜淋巴细胞的组成和表型*
2021-10-20 08:03:50吴紫欣罗青青梁文龙魏建昌李冠炜李旺林
中国病理生理杂志 2021年9期
吴紫欣, 王 菲, 罗青青, 杨 平, 梁文龙, 魏建昌,李冠炜, 魏 芳, 曹 杰, 李旺林
(华南理工大学医学院,广东广州 510006)
随着人们对肠系膜解剖学及功能的认识,肠系膜逐渐被定义为一个新的、独立的器官[1]。肠系膜是一种双层皱褶腹膜,附着于肠道及腹腔内壁,从十二指肠延伸到直肠呈扇形展开,连续地连接着整个肠道,悬吊、固定大小肠于腹腔内以保证肠道的相对位置[1]。肠系膜由表面的间皮细胞、结缔组织间隔及大量脂肪细胞群组成,脂肪组织通过表达被称为脂肪因子的激素和介质参与调节代谢、炎症和免疫功能[2-3]。肠系膜内除了有血管、淋巴管和神经纤维走行,也有较多淋巴结,与肠道建立了大量血液及淋巴液的交通,是收集肠道信息的最佳位置[1],参与介导局部及全身反应。除淋巴管和淋巴结外,肠系膜脂肪细胞间也散在分布着免疫细胞,正常状态下,肠系膜中有大量抑炎免疫细胞,包括M2型巨噬细胞和调节性T(regulatory T,Treg)细胞[4]。然而,肠系膜免疫微环境内淋巴细胞的组成和表型尚无文献报道。
肠系膜作为肠道血液及淋巴液的输送途径,可参与多种肠道疾病,包括结直肠癌(colorectal cancer,CRC)和炎性肠病(inflammatory bowel disease,IBD)等。许多腹腔内肿瘤,如卵巢癌[5]、子宫内膜癌[6]、胃癌[7]和胰腺癌[8]等都有转移到腹膜的倾向,并且出现腹膜转移的患者疾病进展快,预后非常差。此外,多个研究显示脂肪组织尤其内脏肥胖显著促进肿瘤进展[9-10]。同样值得注意的是,肠系膜的临床意义也不仅仅局限于腹部疾病[11-12]。肠系膜是内脏脂肪的最大贡献者[13],它在肥胖、糖尿病和代谢综合征等的病理生理过程中也发挥关键的作用[14-15]。因此,深入探究肠系膜免疫微环境具有重要的科学及临床意义。
在本研究中,我们探讨了肠系膜的免疫微环境,尤其是淋巴细胞的组成和表型及其影响因素,描绘了肠系膜淋巴细胞图谱。我们观察到肠系膜存在与外周血不同的特定免疫微环境,其内的免疫细胞主要由淋巴谱系组成,能在一定程度上发挥重要的免疫调节和细胞毒性等功能,同时性别、年龄、血脂和肠系膜位置影响肠系膜淋巴细胞的组成和表型,为探讨肠系膜在多种疾病中的作用提供了参考资料。
材 料 和 方 法
1 新鲜肠系膜组织及外周血样本
本研究收集了2018~2020 年间在广州市第一人民医院结直肠肛门外科接受手术的26 例CRC 患者的距肿瘤组织>10 cm 的正常肠系膜新鲜组织(normal mesentery,NM)和其中13 例CRC 患者的术前外周血样本以及所纳入患者的临床资料。所有患者均无乙肝病毒、梅毒和HIV 感染,无IBD 病史。广州市第一人民医院伦理审查委员会已通过本实验方案,所有患者均签署知情同意书。
2 方法
2.1 肠系膜组织及外周血样本单细胞悬液的制备仔细分离并去除NM中的淋巴结与血块,将剩余组织用PBS进行多次清洗,剪碎后转入15 mL离心管并加入10 mL组织消化液(含1 g/L Ⅱ型胶原酶和1%BSA的DMEM/F-12培养液)在37 ℃条件下200 r/min 摇床震荡30 min。消化结束后使用200 目尼龙网过滤未消化组织,收集滤液到新的离心管中,离心收集细胞沉淀。随后加入2 mL 红细胞裂解液重悬细胞,4 ℃孵育10 min后加满PBS,离心收集细胞沉淀并进行活细胞计数。2 mL外周血离心去除血清后与生理盐水1∶1 混合稀释,在离心管下层加入3 mL 人淋巴细胞分离液,外周血稀释液沿倾斜的管壁慢慢加到人淋巴细胞分离液上层,室温800×g离心20 min。仔细收集中间白膜层,即外周血单个核细胞(peripheral blood mononuclear cells,PBMCs)于新离心管中,PBS洗涤后离心收集细胞沉淀并进行活细胞计数。
2.2 流式细胞术 NM 和PBMCs 的单细胞悬液首先与野生型小鼠血清孵育进行封闭,并在4 ℃条件下与流式抗体(信息见表1)混合液孵育20 min。孵育结束后加满PBS 洗涤去除未与抗原结合的流式抗体,离心收集细胞沉淀,随后将细胞重悬至300 μL并使用200 目尼龙网滤到流式管中,加入6 μL DAPI 染色液后,使用流式细胞分析仪检测荧光表达。
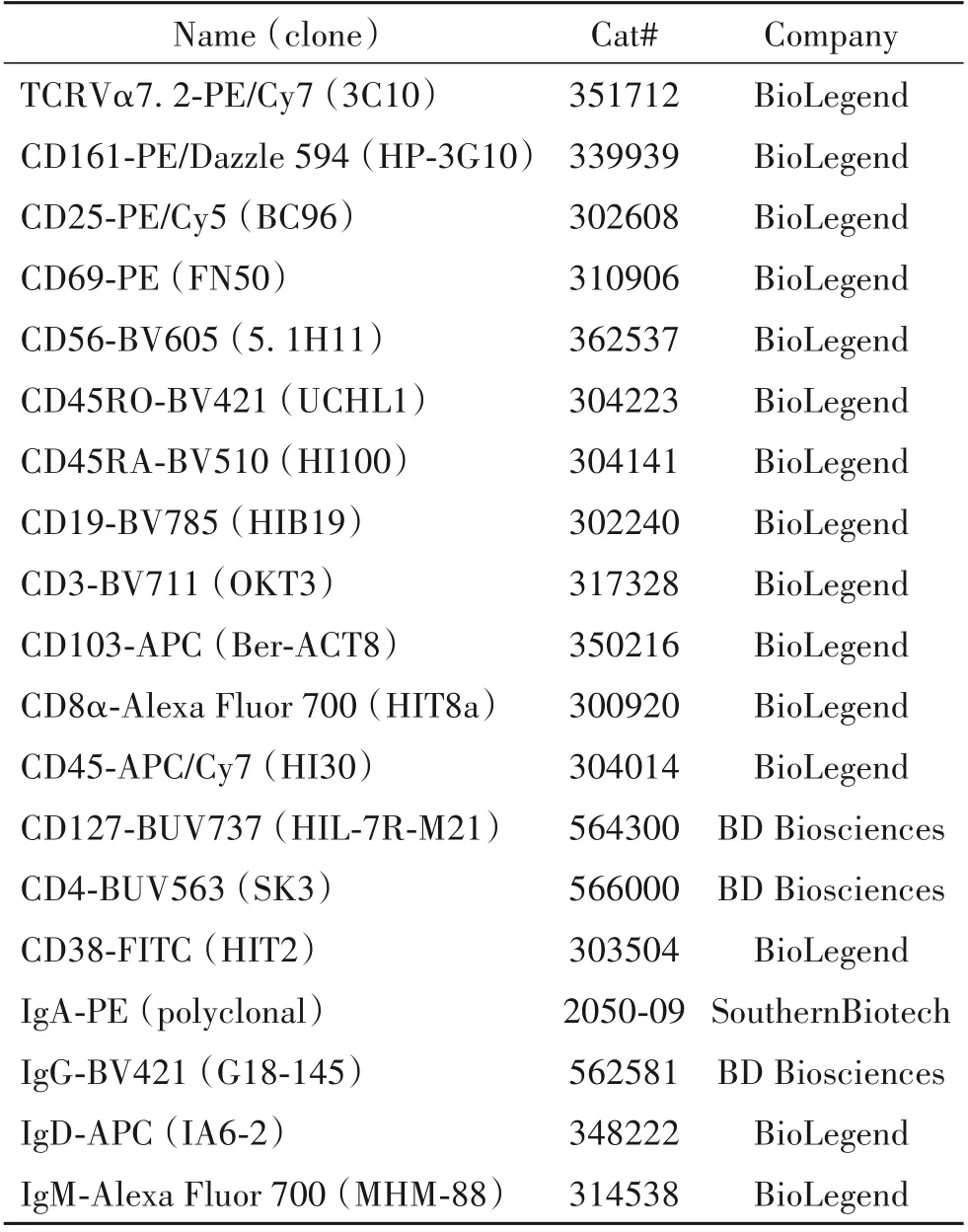
表1 流式细胞术的抗体信息Table 1. Antibody information for flow cytometry
2.3 t 分布随机近邻嵌入(t-distributed stochastic neighbor embedding,t-SNE)算法分析多参数流式细胞术数据 在FlowJo 软件中分别从PBMCs 和NM 的CD45+单个核细胞(mononuclear cells,MNCs)进门后,导出包含各个通道荧光信号值的表达矩阵。分别从PBMCs 和NM 的矩阵中随机选择10 000 个CD45+MNCs,在R3.5.3软件中将它们合并成一个矩阵。使用Seurat 包(版本3.1.5)NormalizeData 函数中的“CLR”算法对表达矩阵的每个通道的荧光表达值进行标准化。使用Seurat 包的RunTSNE 函数对标准化后的表达矩阵进行t-SNE 降维分析,并通过FindNeighbors 与FindClusters 函数进行细胞的聚类和分群,随后基于每个通道的表达分布情况对不同的细胞亚群进行鉴定。用同样的方法分别从PBMCs和NM的T细胞或B细胞进门后,分别随机选择5 500个T细胞或2 000个B细胞进行t-SNE降维分析。
2.4 免疫组织化学(immunohistochemistry,IHC)染色和多重荧光IHC(multiplex IHC,mIHC)染色 (1)IHC染色即厚度为6 μm的肠系膜组织石蜡切片经过环保透明剂脱腊、梯度乙醇水化、高温抗原修复、3%过氧化氢-甲醇消除内源性过氧化物酶活性、10%山羊血清封闭孵育后,每张切片滴加Ⅰ抗(CD4、CD8和CD19)后在湿盒中室温孵育2 h 或4 ℃孵育过夜,TBST 洗涤后滴加Ⅱ抗,TBST 洗涤后使用DAB 显色液显色。随后经苏木素复染、大量自来水返蓝、梯度乙醇脱水、环保透明剂透明和中性树胶封片,最后使用Aperio CS2 数字病理扫描系统扫描。(2)mIHC 染色则是TBST 洗涤Ⅱ抗后使用酪胺信号放大(tyramide signal amplification,TSA)荧光染料标记,TBST洗涤后切片重新抗原修复、封闭后进行下一轮染色,使用不同的酪胺-荧光团偶联物进行染色,直至染色完成。本研究mIHC 的Ⅰ抗配色方案为CD69 配620荧光染料、CD4 配690 荧光染料、Foxp3 配570 荧光染料和CD8 配520 荧光染料。最后滴加DAPI 染色液染细胞核,在湿盒中室温孵育5 min,超纯水漂洗后中性树胶封片,使用Vectra Polaris 全自动定量病理成像系统扫描。
3 统计学处理
利用FlowJo 10 软件和t-SNE 算法分析多参数流式细胞术数据;利用AperioImageScope 12.4.0 软件和Phenochart 1.0.12 软件进行IHC 图像分析;利用GraphPad Prism 8.0.1 软 件、SPSS 25.0 软 件 和R 3.5.3软件进行统计分析。计量资料以均数±标准差(mean±SD)表示。组间差异分析采用非参数Mann-WhitneyU检验;相关性分析采用Pearson 相关分析。以P<0.05为差异有统计学意义。
结 果
1 患者临床资料和正常肠系膜组织的位置
本研究共纳入26例2018年10月~2020年9月在广州市第一人民医院结直肠肛门外科接受手术的CRC 患者,其中14 例为男性(53.80%),12 例为女性(46.20%);平均年龄为61.92岁,其中55岁以下的有5 例(19.20%),55 岁及以上的有21 例(80.80%);身体质量指数(body mass index,BMI)及血液指标如白细胞总数、中性粒细胞绝对值、淋巴细胞绝对值、白蛋白、甘油三酯(triglyceride,TG)、总胆固醇(total cholesterol,TC)、高密度脂蛋白胆固醇(high-density lipoprotein cholesterol,HDL-C)和低密度脂蛋白胆固醇(low-density lipoprotein cholesterol,LDL-C)如表2 所示。收集的NM 位于左半结肠的有15 例(57.70%),位于右半结肠的有11例(42.30%),见表2。

表2 患者的临床资料及正常肠系膜组织的位置Table 2. Clinical data of patients and the location of the normal mesentery(NM)
2 肠系膜免疫细胞以T细胞为主
首先对肠系膜进行IHC 染色和mIHC 染色,结果显示肠系膜脂肪细胞之间散在分布了CD4+T 细胞、CD8+T 细胞、B 细胞(CD19+)、CD4+CD69+T 细胞、CD8+CD69+T 细胞和Treg 细胞(CD4+Foxp3+),见图1。接着我们根据细胞表面分子包括CD3、CD56、CD161、TCRVα7.2 和CD19 的表达对PBMCs 和NM的CD45+MNCs 进行t-SNE 降维分析并界定出6 个免疫细胞群,包括B 细胞(CD19+)、黏膜相关恒定T(MAIT)细胞(CD3+TCRVα7.2+CD161+)、NK 细胞(CD3-CD56+)、T细胞(CD3+CD56-)、CD3+CD56+细胞和其他细胞(单核细胞、巨噬细胞和树突状细胞等),见图2A 及 图3A、B。从PBMCs 和NM 中的CD45+MNCs 的t-SNE 降维图可以看出二者的B 细胞、T 细胞、NK 细胞和CD3+CD56+细胞的组成和分布存在差异,见图3A。同时,根据图2A所示CD45+MNCs的画门策略对流式数据进行分析,结果显示NM 中B 细胞、MAIT 细胞、CD3+CD56+细胞、NK 细胞、T 细胞和其他细胞(包括单核细胞、巨噬细胞和树突状细胞等)分别约占CD45+MNCs 的9.96%、2.42%、4.79%、5.12%、58.65%和18.01%,即NM 中的免疫细胞以T细胞为主,见图3C。与PBMCs 相比,NM 中的NK 细胞的比例显著降低(P<0.01),而T 细胞的比例显著升高(P<0.01),见图3C。
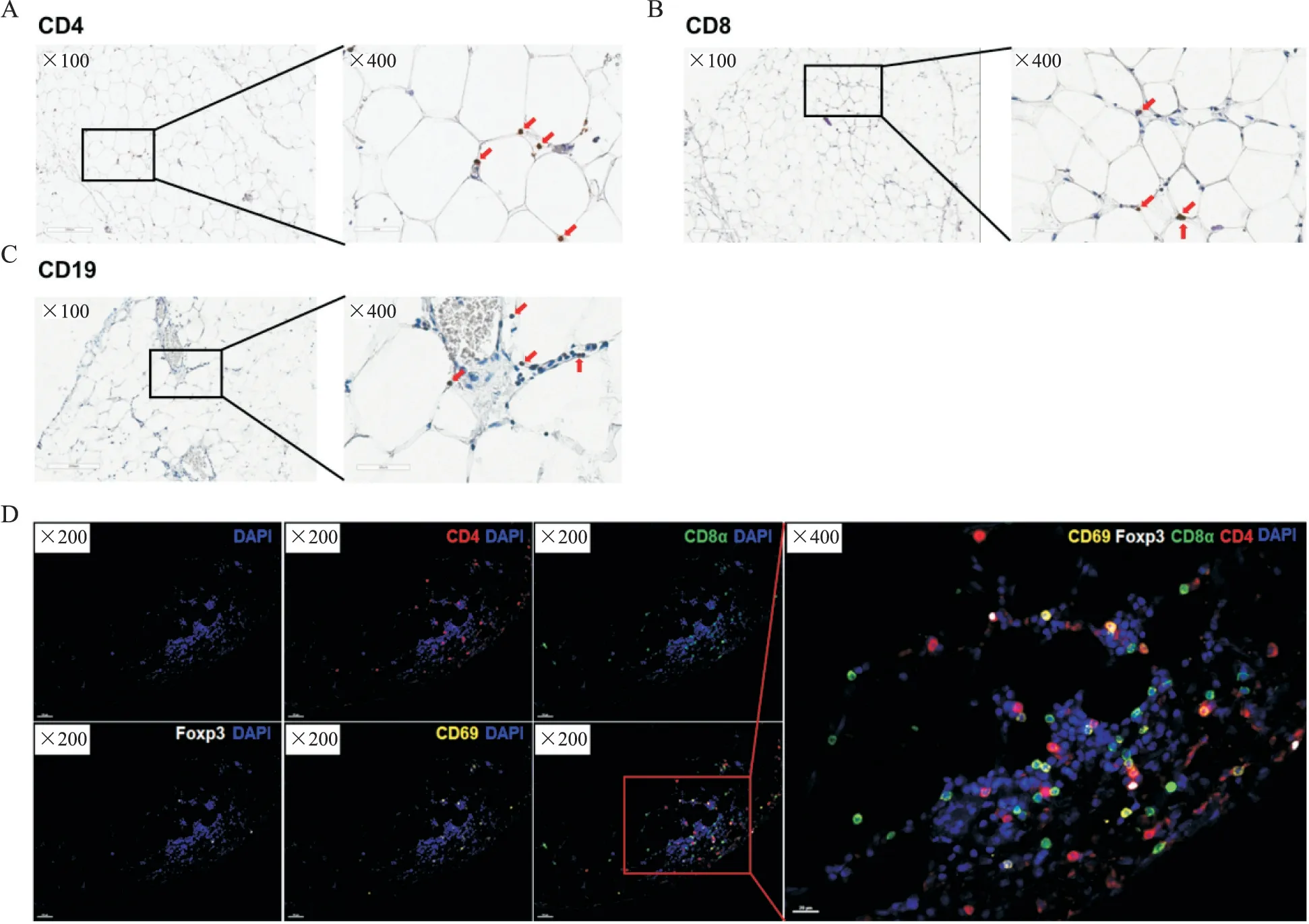
Figure 1. Scattered distribution of T cells and B cells in the mesentery. A:scattered distribution of CD4+ T cells(red arrows)in the mesentery;B:scattered distribution of CD8+ T cells(red arrows)in the mesentery;C:scattered distribution of B cells(CD19+;red arrows)in the mesentery;D:mIHC showed the expression of CD4(red),CD8(green),Foxp3(white)and CD69(yellow)in the mesentery[DAPI(blue)indicated the nuclei],and CD4+T cells,CD4+CD69+T cells,CD8+cells,CD8+CD69+T cells and Treg cells(CD4+Foxp3+)were scattered in the mesentery.图1 肠系膜内散在分布着T细胞和B细胞亚群
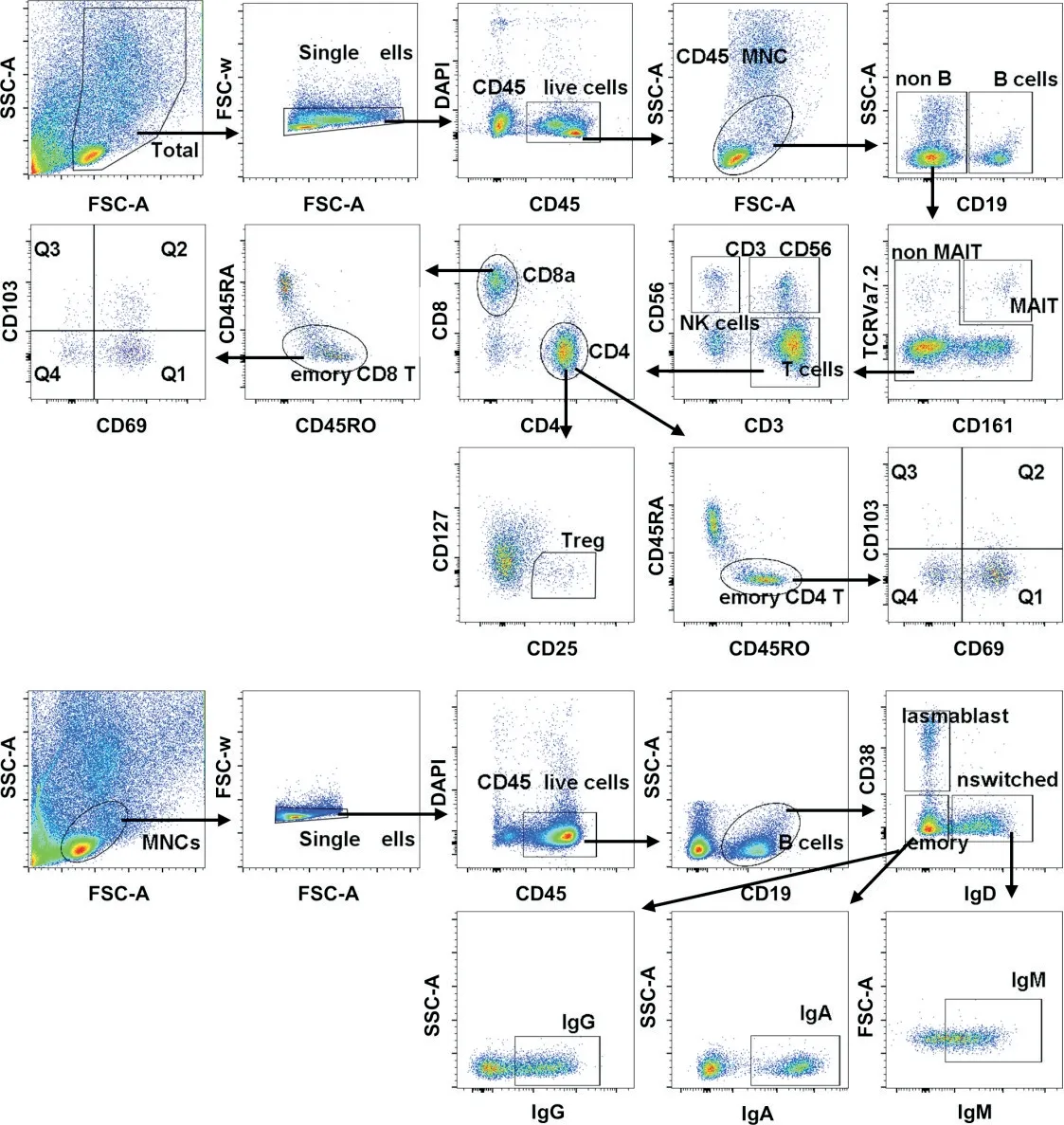
Figure 2. Gating strategy of CD45+MNC subsets,T-cell subsets and B-cell subsets. A:gating strategy of CD45+MNC subsets and Tcell subsets;B:gating strategy of B-cell subsets.图2 CD45+MNCs、T细胞和B细胞亚群的画门策略
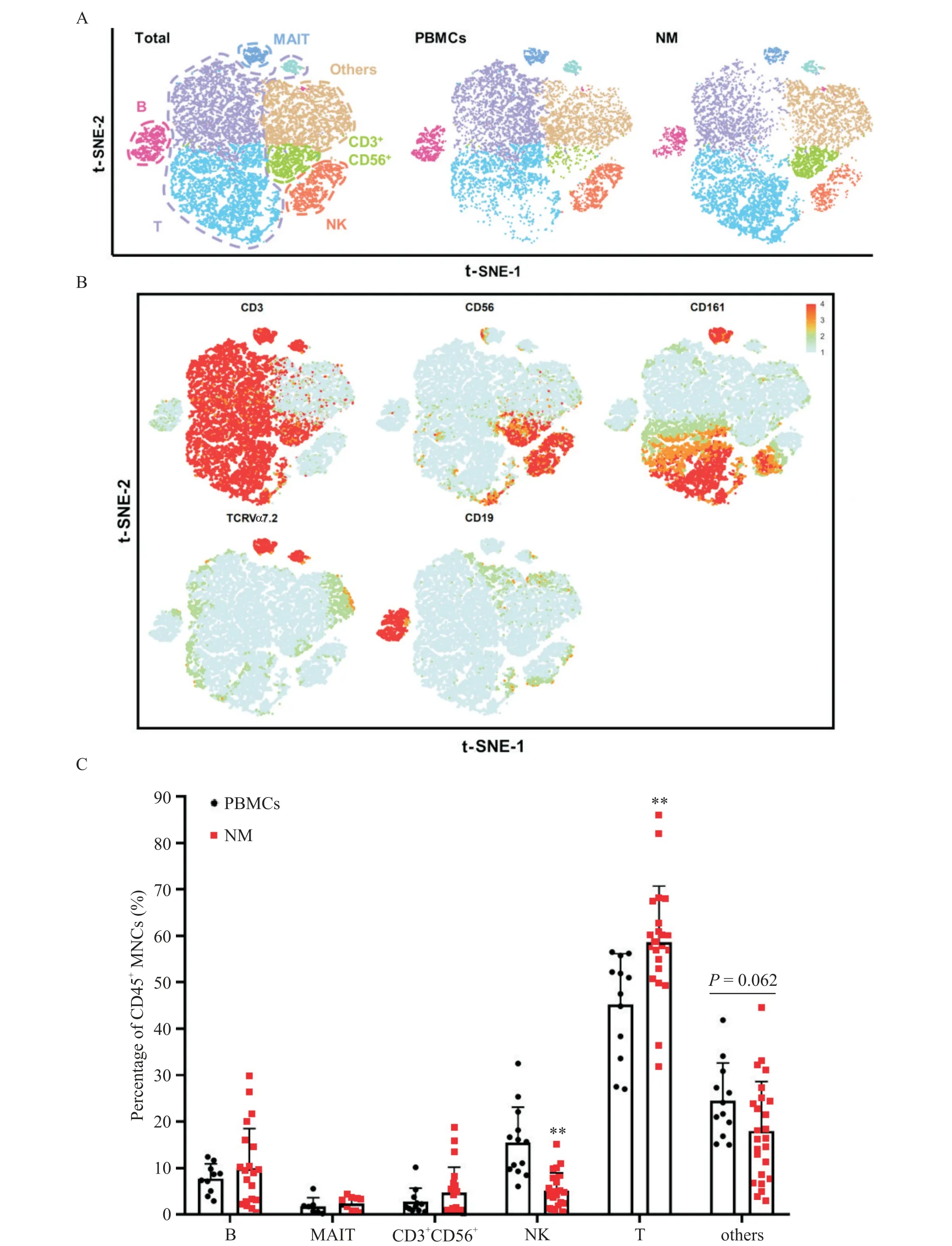
Figure 3. Immune cells in the mesentery were mainly T cells. A:t-SNE plot of CD45+ MNCs in the PBMCs and NM with main subsets indicated;B:feature plot of CD45+ MNC markers;C:percentages of CD45+ MNC subsets in the PBMCs(n=13)and NM(n=26). Mean±SD.**P<0.01 vs PBMCs.图3 肠系膜免疫细胞以T细胞为主
3 肠系膜中浸润高比例的记忆T 细胞和组织驻留记忆T(tissue-resident memory T,TRM)细胞
根 据 CD4、CD8、CD25、CD127、CD45RA、CD45RO、CD69 和CD103 等特定T 细胞表面分子对PBMCs和NM 的T细胞作进一步降维及细分亚群,鉴定出包括记忆CD4+T 细胞(CD4+CD45RO+)、幼稚CD4+T 细胞(CD4+CD45RA+)、记忆CD8+T 细胞(CD8+CD45RO+)、幼稚CD8+T细胞(CD8+CD45RA+)、双阴性(DN)细胞(CD4-CD8-)、Treg 细胞(CD4+CD25+CD127-)、CD4+TRM 细 胞(CD4+CD45RO+CD69+)和CD8+TRM 细胞(CD8+CD45RO+CD69+)等不同的T 细胞亚群,见图4A、B。结果显示CD69 在PBMCs 中的T 细胞几乎不表达,而在NM 的T 细胞中有着显著表达,即NM 存在大量CD4+TRM 细胞和CD8+TRM 细胞,见图4A。此外,根据图2A 所示T 细胞亚群的画门策略对流式数据进行分析,统计结果表明NM 中的CD4+T 细胞与CD8+T 细胞的比值约为2.26;Treg细胞、记忆CD4+T细胞、记忆CD8+T细胞、CD4+TRM 细胞和CD8+TRM 细胞分别约占CD4+T 细胞 或CD8+T 细 胞 的10.71%、80.26%、61.90%、58.27%和51.36%,见图4C~F。与PBMCs 相比,NM的记忆CD4+T 细胞(P<0.01)、记忆CD8+T 细胞(P<0.01)、CD4+TRM 细胞(P<0.01)和CD8+TRM 细胞(P<0.01)的比例显著升高,见图4E、F。
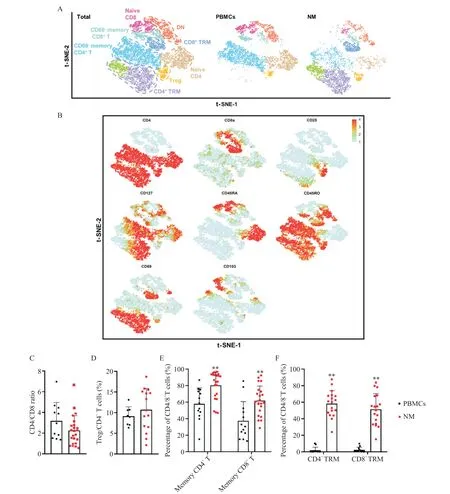
Figure 4. Higher percentages of memory T cells and tissue-resident memory T(TRM)cells were infiltrated in the mesentery. A:t-SNE plot of T cells in the PBMCs and NM with main subsets indicated;B:feature plot of T-cell markers;C:CD4/CD8 ratio in the PBMCs(n=11)and NM(n=23);D:percentage of Treg cells in the PBMCs(n=8)and NM(n=15);E:percentages of memory CD4+ T cells and memory CD8+ T cells in the PBMCs(n=13)and NM(n=22);F:percentages of CD4+TRM cells and CD8+TRM cellsin the PBMCs(n=12)and NM(n=18). Mean±SD.**P<0.01 vs PBMCs.图4 肠系膜中浸润高比例的记忆T细胞和组织驻留记忆T细胞
4 肠系膜中浆母细胞和记忆B细胞浸润增加
根据CD38、IgD、IgM、IgA 和IgG 等特定B 细胞表面分子对B 细胞进行降维分析,将B 细胞分成5 个亚群,包括浆母细胞(CD38+IgD-)、IgM+和IgM-未转化的B 细胞(CD38-IgD+)及IgA+和IgG+记忆B 细胞(CD38-IgD-),见图5A、B。降维图显示PBMCs和NM之间表达IgA 或IgG 的记忆B 细胞的组成和分布存在差异,见图5A。根据图2B 的B 细胞亚群画门策略对流式数据进行分析,结果显示NM 中浆母细胞、记忆B 细胞、未转化B 细胞、IgA+记忆B 细胞、IgG+记忆B 细胞和IgM+未转化B 细胞分别约占B 细胞的24.93%、31.99%、35.60%、12.83%、13.72% 和32.31%,见图5C、D。与PBMCs 相比,NM 的浆母细胞(P<0.05)、记忆B 细胞(P<0.01)和IgA+记忆B 细胞(P<0.05)的比例显著升高,而IgM+未转化B 细胞比例显著降低(P<0.01),见图5C、D。
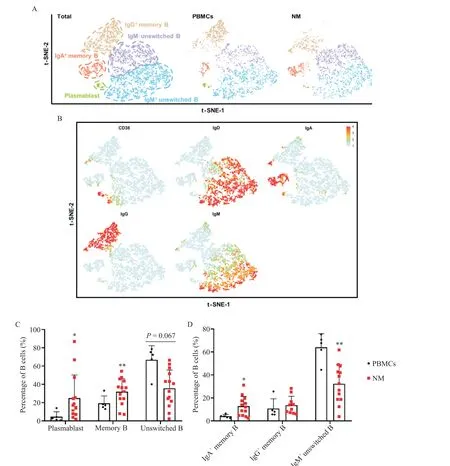
Figure 5. Higher percentages of plasmablasts and memory B cells were infiltrated in the mesentery. A:t-SNE plot of B cells in the PBMCs and NM with main subsets indicated;B:feature plot of B-cell markers;C:percentages of plasmablasts,memory B cells and unswitched B cells in the PBMCs(n=5)and NM(n=14);D:percentages of IgA+ memory B cells,IgG+ memory B cells and IgM+unswitched B cells in the PBMCs(n=5)and NM(n=13). Mean±SD.*P<0.05,**P<0.01 vs PBMCs.图5 肠系膜中浆母细胞和记忆B细胞浸润增加
5 性别、年龄、血脂和肠系膜位置影响肠系膜T 细胞和B细胞亚群的组成
相比于男性,女性肠系膜B 细胞的比例更高(P<0.01),而记忆CD8+T 细胞的比例更低(P<0.05),见图6A、B。年龄也会影响肠系膜T细胞和B细胞的组成,其中55 岁及以上患者肠系膜中有更高比例的CD8+TRM 细胞(P<0.05)和CD8+CD103-TRM 细胞(P<0.01),见图6C;同时年龄与肠系膜CD8+TRM 细胞、CD8+CD103-TRM 细胞和浆母细胞比例呈正相关,与肠系膜未转化B 细胞比例呈负相关(P<0.05),见图6D~H。此外,血脂也会影响肠系膜T 细胞和B细胞的组成,血TC 和LDL-C 的含量均与肠系膜记忆CD8+T 细胞(P<0.05)、CD4+TRM 细胞(P<0.01)、CD4+CD103-TRM 细胞(P<0.01)、CD8+TRM 细胞(P<0.05)和CD8+CD103-TRM 细胞(P<0.05)比例呈正相关,见图7A~J;血HDL-C含量与肠系膜IgA+记忆B 细胞比例呈正相关(P<0.05),见图7K。患者的血TG 含量、血白蛋白含量和BMI 与肠系膜淋巴细胞亚群均无显著相关性(P>0.05)。以脾曲为界划分右半结肠和左半结肠,结果显示左半结肠肠系膜与右半结肠肠系膜的T细胞和B细胞亚群组成存在差异,右半结肠肠系膜的CD4+T 细胞与CD8+T 细胞比值(P<0.01)和CD8+CD103+TRM 细胞(P<0.05)比例更高,见图6I~K。
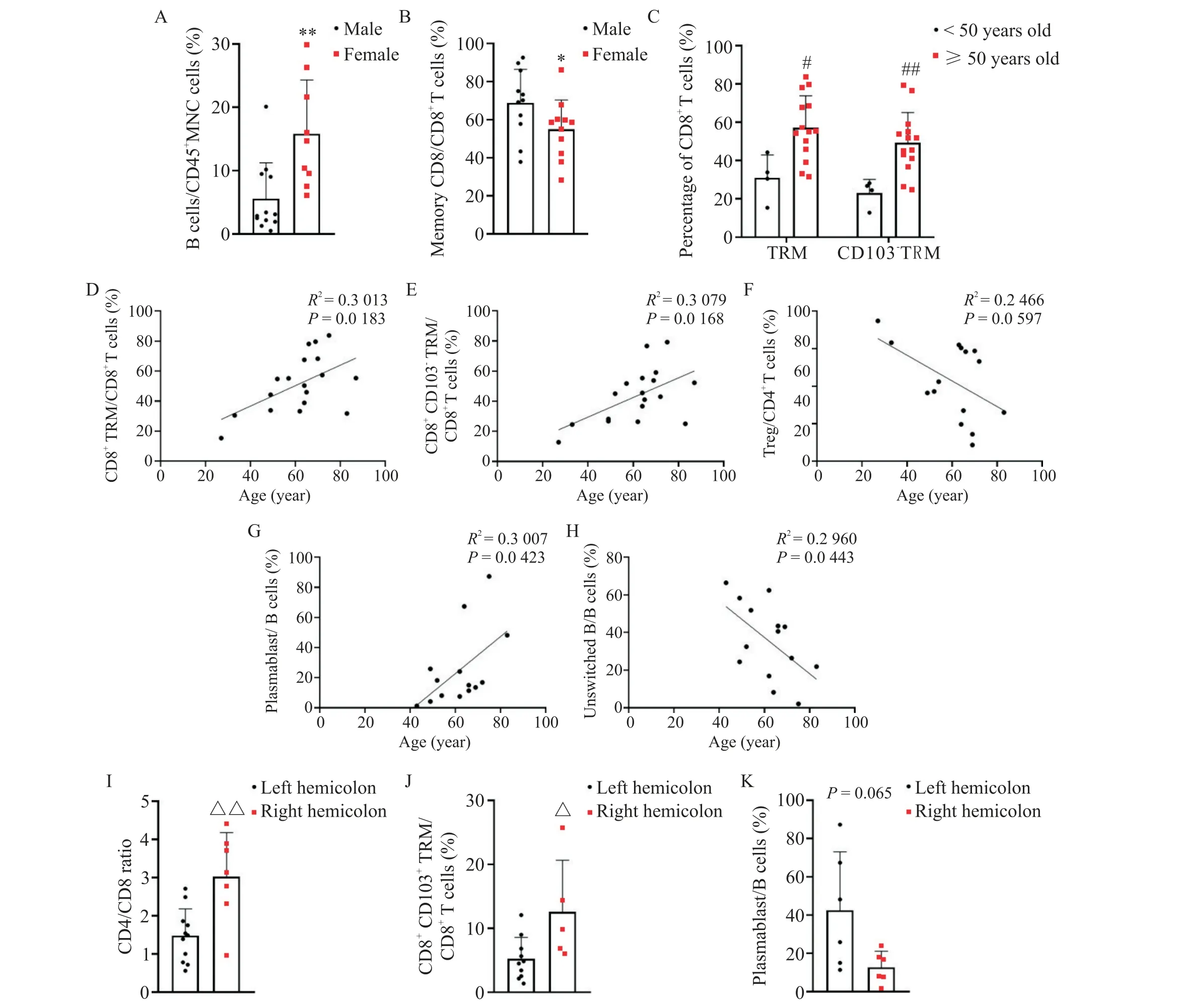
Figure 6. Sex,age and location of the mesentery affect the composition of mesenteric T-cell and B-cell subsets. A:percentage of mesenteric B cells in males(n=12)and females(n=9);B:percentage of mesenteric memory CD8+ T cells in males(n=11)and females(n=11);C:percentages of mesenteric CD8+ TRM cells and CD8+ CD103- TRM cells in the patients <50 years old(n=4)and ≥50 years old(n=14);D,E and F:correlations between age and the percentages of mesenteric CD8+TRM cells,CD8+ CD103- TRM cells and Treg cells(n=18);G and H:correlations between age and the percentages of plasmablasts and unswitched B cells(n=14);I:CD4/CD8 ratio in the left hemicolon(n=11)and right hemicolon(n=7);J:percentage of mesenteric CD8+ CD103+ TRM cells in the left hemicolon(n=10)and right hemicolon(n=5);K:percentage of mesenteric plasmablasts in the left hemicolon(n=6)and right hemicolon(n=6). Mean±SD.*P<0.05,**P<0.01 vs male;#P<0.05,##P<0.01 vs <50 years old;△P<0.05,△△P<0.01 vs left hemicolon.图6 性别、年龄和肠系膜位置影响肠系膜T细胞和B细胞亚群的组成
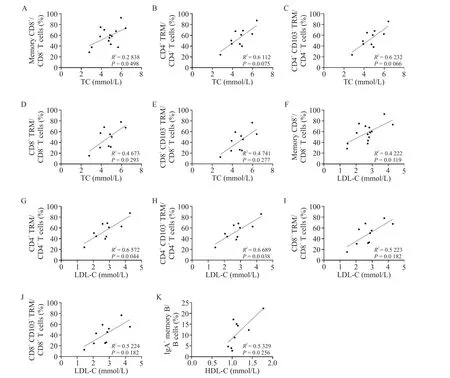
Figure 7. Blood lipids affected the composition of mesenteric T-cell and B-cell subsets. A to E:correlations between blood TG levels and the percentages of mesenteric memory CD8+ T cells,CD4+ TRM cells,CD4+ CD103- TRM cells,CD8+ TRM cells and CD8+CD103-TRM cells;F to J:correlations between blood LDL-C levels and the percentages of mesenteric memory CD8+T cells,CD4+TRM cells,CD4+CD103-TRM cells,CD8+TRM cells and CD8+CD103-TRM cells;K:correlation between blood HDL-C and the percentage of IgA+memory B cells. n=10 to 14.图7 血脂影响肠系膜T细胞和B细胞亚群的组成
讨 论
肠系膜近年来被认为是一个独立的器官,其组织学特征是表面的间皮细胞、结缔组织间隔及大量脂肪细胞群[1]。肠系膜作为肠道血液及淋巴液的输送途径,在多种腹部及非腹部疾病的病理生理过程中具有重要作用,如CRC可以侵袭到邻近的肠系膜,发生肠系膜转移的CRC 患者往往累积生存率较差[16-17],而且研究表明肠系膜是一个潜在的肿瘤残留部位,可导致CRC 的术后复发[18-20]。此外,许多腹腔内肿瘤,如卵巢癌[5]、子宫内膜癌[6]、胃癌[7]和胰腺癌[8]等倾向转移腹膜,发生腹膜转移的患者疾病进展快、预后差。肠系膜是内脏脂肪的最大贡献者[13],多个研究也显示脂肪组织尤其内脏肥胖显著促进肿瘤进展[9-10]。然而,肠系膜的免疫微环境尚不清楚。在本研究中,我们首次证实了肠系膜中存在与外周血不同的特定免疫微环境,其内的免疫细胞主要由淋巴谱系组成,以T 细胞为主。与PBMCs 相比,肠系膜中显著浸润更高比例的记忆T 细胞、TRM 细胞、浆母细胞和记忆B 细胞。此外,性别、年龄、血脂和肠系膜位置可影响肠系膜T细胞和B细胞亚群的组成。总的来说,本研究揭示了肠系膜中淋巴细胞亚群的组成与表型及其影响因素,为探讨肠系膜在多种疾病中的作用提供了参考资料。
根据行使免疫功能的不同,淋巴细胞亚群可分为效应细胞、调节细胞和记忆细胞。目前研究最多的调节细胞是Treg 细胞,它具有免疫应答低下和免疫抑制两大特征,可通过细胞接触或分泌抑制性细胞因子作用于抗原提呈细胞或效应T 细胞,发挥免疫负向调节的作用[21]。记忆T 细胞由记忆CD4+T 细胞和记忆CD8+T 细胞组成,各自又可分成效应性记忆T(effector memory T,TEM)细胞、中枢性记忆T(central memory T,TCM)细胞和TRM 细胞。其中TEM 细胞承担保护性记忆形式速发型效应功能;TCM 细胞负责反应性记忆,在抗原再次刺激时重新分化为效应细胞,如CD4+辅助性T(T helper,Th)细胞、CD8+细胞毒性T 淋巴细胞(cytotoxic T-lymphocytes,CTLs)等[22]。其中CD4+Th 细胞主要通过分泌多种细胞因子在肿瘤免疫、自身免疫等发挥免疫调节作用[23];而CD8+CTLs 是免疫应答的主要效应细胞,可以特异性结合靶细胞并释放颗粒酶和穿孔素使其裂解和分泌TNF-α、IFN-γ 和IL-2 等协同杀伤靶细胞,在肿瘤免疫和抗病毒免疫中发挥重要作用[24-25]。CD69 的表达意味着T 细胞的早期活化及其在组织中的驻留[26-27],TRM 细胞表现为上调CD69 的表达从而维持在组织中的驻留及其细胞毒性特征[28-30]。TRM 细胞在成年人中数量居所有记忆T 细胞之首,定居于与外周非淋巴组织,是在应答第一线发挥作用的记忆细胞[28-30]。CD8+CD69+T 细胞已经被证明具有更高的细胞毒性[31]。此外,B细胞是体液免疫中最主要的免疫细胞,同时B 细胞还可以诱导性表达参与提呈抗原的MHC II 类分子和共刺激分子,具有摄取、加工和提呈抗原,协助启动T 细胞应答的功能。B 细胞同时还能分泌细胞因子调节免疫应答[32]。根据CD38 和IgD 的表达,B 细胞可以分为浆母细胞(CD38+IgD-)、未转化的B 细胞(CD38-IgD+)和记忆B 细胞(CD38-IgD-)[33]。B 细胞接受抗原刺激之后可分化为体积较大的浆母细胞,而后进一步分化为浆细胞,浆细胞细胞质中有大量糙面内质网,能合成和分泌特异性抗体,介导体液免疫。而记忆B细胞在再次遇到抗原刺激时,记忆B细胞可再次分化为浆细胞,产生记忆性应答。此外,在抗原的刺激下,B 细胞可以发生类别转换重组,从初始表达膜结合IgM 和IgD到表达膜结合的IgG、IgA或IgE[34]。我们的研究结果显示肠系膜免疫细胞主要由淋巴谱系组成,以T 细胞为主,与外周血相比,肠系膜中显著浸润更高比例的T 细胞(P<0.01)、记忆T 细胞(P<0.01)、TRM 细胞(P<0.01)、浆母细胞(P<0.05)和记忆B 细胞(P<0.01),即肠系膜除了作为肠道的支撑结构,其内也浸润多种淋巴细胞亚群,表明肠系膜一定程度上在抗感染免疫、自身免疫和肿瘤免疫中能发挥重要的免疫调节和细胞毒性等免疫功能。
此外,我们的数据表明性别、年龄、血脂和肠系膜位置影响肠系膜T细胞和B细胞亚群的组成,其中女性肠系膜B 细胞的比例更高(P<0.01),记忆CD8+T 细胞的比例更低(P<0.05),不同性别的免疫差异可能是性染色体、基因、激素和环境等共同影响的结果[35-37]。我们还观察到年龄越大的患者的肠系膜CD8+TRM 细胞和CD8+CD103-TRM 细胞的比例更高(P<0.05),这可能是由于机体衰老伴随着免疫功能降低,机体长期暴露在抗原(包括肿瘤抗原、病原体或自身抗原)下,导致初始细胞向记忆细胞分化[38]。已有文献报道,多种脂质参与调节许多生理活动,如能量储存、细胞凋亡和信号传导等,胆固醇和LDL-C 等脂质在调节淋巴细胞增殖、分化、激活及分泌细胞因子能力中发挥重要作用[39-40]。我们的研究结果显示,血TC 和LDL-C 含量均与肠系膜记忆CD8+T 细胞(P<0.05)、CD4+TRM 细胞(P<0.01)、CD4+CD103-TRM 细胞(P<0.01)、CD8+TRM 细胞(P<0.05)和CD8+CD103-TRM 细胞(P<0.05)比例呈正相关,HDL-C含量与肠系膜IgA+记忆B细胞比例呈正相关(P<0.05),表明TC 和LDL-C 等血脂的增加可能促进肠系膜T 细胞向记忆T 细胞和TRM 细胞分化。此外,临床上常以脾曲为界将结肠划分为右半结肠和左半结肠;它们的胚胎起源也不同,右半结肠起源于胚胎的中原肠,而左半结肠则是起源于后原肠[41];左右半结肠疾病如结肠癌等在临床表现、病理分型、免疫环境、分子生物学特征及预后等方面均存在差异[42]。我们的研究结果显示,左半结肠肠系膜与右半结肠肠系膜的T 细胞和B 细胞亚群组成也同样存在差异,即右半结肠肠系膜CD4+T 细胞与CD8+T 细胞的比值(P<0.01)和CD8+CD103+TRM 细胞的比例(P<0.05)更高,表明不同位置的肠系膜在疾病中的作用可能存在差异。
综上所述,我们揭示了肠系膜中存在与外周血不同的特定免疫微环境,并描绘了肠系膜的淋巴细胞图谱,这些细胞在一定程度上能发挥重要的免疫调节和细胞毒性等免疫功能,同时性别、年龄、血脂和肠系膜位置影响肠系膜淋巴细胞的组成和表型。
猜你喜欢
现代临床医学(2023年1期)2023-03-24 08:30:06
中国医药科学(2022年5期)2022-05-05 23:58:07
天津医科大学学报(2021年4期)2021-08-21 02:14:52
中国民间疗法(2020年22期)2021-01-07 07:39:34
医学新知(2019年4期)2020-01-02 11:03:52
基层中医药(2018年7期)2018-12-06 09:25:46
中国医学影像学杂志(2018年9期)2018-10-17 01:27:12
中国卫生标准管理(2015年3期)2016-01-14 03:41:45
中国卫生标准管理(2015年3期)2015-01-27 00:57:55
西南军医(2015年4期)2015-01-23 01:19:04
