
气候变化背景下三尾凤蝶潜在适生区模拟及保护空缺分析
2021-10-20 06:42柳晓燕李秀山赵彩云李飞飞朱金方纪文婧
环境昆虫学报 2021年5期
柳晓燕,李秀山,赵彩云*,李飞飞,朱金方,纪文婧
(1.中国环境科学研究院,北京 100012;2.西华师范大学,教育部西南地区野生动植物保护重点实验室,四川南充 637002)
三尾凤蝶Bhutanitisthaidina又称三尾褐凤蝶、中华褐凤蝶,为昆虫纲Insecta鳞翅目Lepidoptera凤蝶科Papiloinidae尾凤蝶属Bhutanitis昆虫(周尧,1992;吴静等,2007),因具有较高的科学研究价值和观赏价值,被列入中国国家Ⅱ级保护野生动物名录,2019年1月,世界自然保护联盟濒危物种红色名录(IUCN,Red List of Threatened Species)将其列为近危(NT,Near Threatened)物种(IUCN 2020),在我国主要分布于陕西、甘肃、四川、湖北、云南、西藏等地区(杨萍等,2006;吴静等,2007)。受气候变化及人类干扰影响,三尾凤蝶在我国正面临着栖息地丧失、生境破碎化、寄主植物保护力度欠缺等威胁(Hilletal.,2002;易传辉等,2011),开展气候变化背景下濒危蝶类保护空缺研究已成为我国生物多样性保护研究的重要问题之一。
气候变化作为当今世界面临的重大科学问题,对全球范围内绝大多数生态系统结构和功能、生物类群组成及物种分布格局产生深刻的影响(Waltheretal.,2002)。研究表明,气候变化常伴随气温上升、局部地区降雨模式改变等非生物因子影响物种生存(Somero,2010;郑巧燕和卫万荣,2019),这为运用物种分布模型定量化的探讨气候变化对物种的分布影响提供了可能。MaxEnt作为一种基于最大熵原理的物种分布模型(Phillipsetal., 2006),能够很好地处理物种分布数据与降水、温度等非生物因子间复杂的交互作用,是评估和预测物种过去及未来地理分布格局的重要研究工具(Iverson and McKenzie,2013),被广泛应用于气候变化背景下物种适生区模拟和生物多样性保护研究(Huetal., 2019;白雪红等,2019;蒋丽华等,2019)。
建立自然保护区是保护生物多样性最有效的方式之一(薛达元和蒋明康,1995;马建章等,2012)。截止目前,我国已建立了2 750个不同级别的自然保护区,极少保护区专门针对三尾凤蝶等濒危蝶类开展保护,三尾凤蝶等濒危蝶类保护现状仍不明朗。此外,国内外三尾凤蝶研究主要集中于实地调查、生物学特性等方面(易传辉等,2011),三尾凤蝶保护性研究多以定性描述为主,国内外尚无基于三尾凤蝶适生区模拟结果定量化开展保护空缺分析的先例。为此,本研究基于物种分布数据和气候因子数据,运用MaxEnt物种分布模型和地理信息系统分析三尾凤蝶当前及未来气候情景下的适生区分布格局,探讨不同气候情景下三尾凤蝶适宜区变化规律,同时基于自然保护区对三尾凤蝶适生区覆盖状况识别保护空缺范围,以期为我国自然保护区保护成效评估以及三尾凤蝶等濒危蝶类物种保护工作提供参考。
1 材料与方法
1.1 数据来源
1.1.1物种数据
三尾凤蝶分布数据主要来源于2019年7月-8月在甘肃小陇山、秦岭太白山、安康旬阳坝、四川芦山县、泸定县、云南省大理市、丽江玉龙雪山、高黎贡山等地开展的野外调查,并结合历史文献、标本数据库、博物馆等收集的分布数据(Gaoetal., 2014; 易传辉等,2017;Huetal., 2019)。为了降低采样点的空间自相关以及与环境数据范围一致,将研究区分成1 km×1 km的网格,随机获取其中唯一分布点,最终得到50条三尾凤蝶分布记录(图3)。
1.1.2环境变量数据
研究考虑了23个环境因子,包括19个气候因子、3个地形因子(海拔、坡度和坡向)、1个土地利用数据。当前气候因子源于WorldClim 2.1(http://www.worldclim.org/),包括生物气候因子(bio1~bio19),分辨率为30 arc-sec(1 km×1 km),数据年份为1970-2000年。海拔源于GTOPO(http://edcftp.cr.usgs.gov/pub/data/gtopo30/global/),基于GIS技术衍生为坡度(slope)及坡向(aspect)。土地覆被数据来源于国家地球系统科学数据中心,数据年份为2015年。
未来时期气候数据采用全球气候数据库WorldClim 2.1中我国国家气候中心开发的BCC_CSM2_MR模型相对应数据,其在全球温度和降水分布、全球温度变化、东亚降水季节性移动等方面的模拟能力更高(Wuetal.,2019;辛晓歌等,2019)。未来气候情景包括4种共享的社会经济途径(SSPs,Shared Socioeconomic Pathways),其中SSPs 1~2.6代表了低辐射强迫的可持续发展路径,2100年辐射强迫稳定在2.6 W/m2;SSPs 2~4.5代表着中等辐射强迫的中间发展路径,2100年辐射强迫稳定在4.5 W/m2;SSPs 3~7.0代表了区域竞争发展路径,2100年辐射强迫稳定在7.0 W/m2;SSPs 5~8.5为高辐射强迫下的常规发展路径,2100年辐射强迫稳定在8.5 W/m2(van Vuurenetal.,2011;张丽霞等,2019)。辐射强迫越高,增温越快。分辨率为2.5′(4.5 km×4.5 km),基于GIS 10.6重采样为1 km×1 km。数据年份为2050年(2041-2060年)。
1.1.3自然保护区图层
自然保护区边界来自原环境保护部提供的2015年国家自然保护区边界。
1.1.4基础地理数据
从自然资源部标准地图服务中心(http://bzdt.ch.mnr.gov.cn/)下载的1∶1100万中国地图。
1.2 研究方法
1.2.1环境变量筛选
为避免环境变量过度拟合,本研究基于各环境变量多重共线性分析,对具有较高相关度的环境变量进行筛选(|r|>0.8)(Yangetal.,2013),最终确定了年平均温(bio1)、昼夜温差月均值(bio2),温度变化方差(bio4)、年平均湿度(bio12)、湿度变化方差(bio15)、海拔(Altitude)、坡度(Slope)、坡向(Aspect)、土地覆被(Landuse)等9个环境变量。
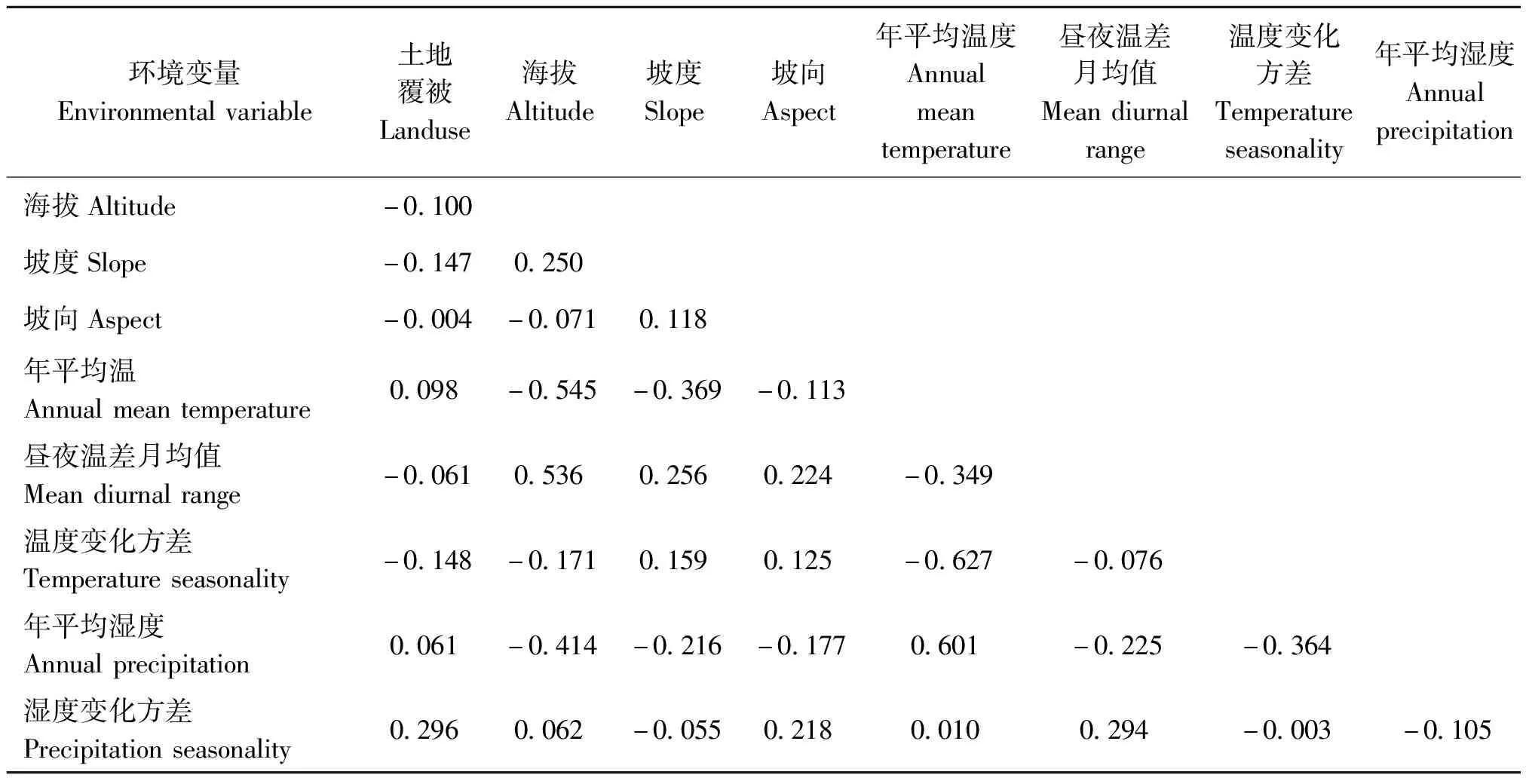
表1 本研究中环境变量相关系数矩阵
1.2.2 物种分布模型建模与验证
将三尾凤蝶的地理分布数据、当前时期和未来时期气候数据分别导入MaxEnt 3.4.1中,随机选取75%的点建模,25%的点验证模型,采用刀切法(Jackknife)获得每个环节变量对物种适生性的贡献率,分析结果以Logistic格式、ASCII类型文件输出。为避免过度拟合,研究通过调用ENMeval数据包来调整调控倍频(regularization multiplier,RM)和特征组合(feature combination,FC)参数,选择AICc最小值作为最优设置并建立最终模型(朱耿平和乔慧捷,2016)。调控倍频RM设置为0.5、1、1.5、2、2.5、3、3.5、4共8个水平,特征组合(FC)在模型中有线性(linear-L),二次型(quadratic-Q),片段化(hinge-H)和乘积型(product-P)等4种类型,本研究选用了6个特征组合,即L、LQ、H、LQH、LQP 和LQHP。为提升模型预测性能,研究结合三尾凤蝶幼虫寄主植物马兜铃科植物分布范围对研究区进行了合理界定(朱耿平等,2014),囊括了云南省、西藏藏族自治区、四川省、陕西省、青海省、甘肃省、宁夏回族自治区、贵州省、重庆市、湖北省、湖南省、浙江省、江西省、江苏省、福建省、河南省、广东省、广西壮族自治区、安徽省、山东省等20个省、市、自治区(易传辉等,2011; 2017)。
模型建立后,基于受试者工作特征曲线(receiver operating characteristic curve,ROC)与横坐标围成的面积,即AUC(area under curve)值来评价模型预测结果的精准度。AUC值越接近1,表明预测与随机分布相距越远,模型预测效果越好(Hanley and McNeil,1982)。一般来言,AUC值位于0.5~0.6为失败,0.6~0.7为较差,0.7~0.8为一般,0.8~0.9为良好,0.9~1.0为优秀(Swets,1988)。
1.2.3物种分布适宜区界定
MaxEnt模型预测的结果是适生值位于0~1之间的连续栅格数据,适生值越接近1,表示物种存在的适生概率越高。结合相关学者分析结果(孔维尧等,2019;柳晓燕等,2019),研究基于建模数据敏感度与特异性相等的阈值判定方法将预测结果划分为四个等级:适生值<0.15为非适生区;0.15≤适生值<0.4为低适生区;0.4≤适生值<0.7为中适生区;0.7≤适生值<1为高适生区;为实现三尾凤蝶景观格局和保护空缺研究,研究聚焦于三尾凤蝶中适生区和高适生区,参考了IPCC第四次评估报告中的阈值划分方法和相关学者(白雪红等,2019)的划分方法,以适生值≥0.4的概率值对三尾凤蝶适生区二值化,并将符合要求的三尾凤蝶适生区统称为“中-高度适生区”。
1.2.4不同气候情景下三尾凤蝶适生区景观格局评价
选取了三尾凤蝶适生斑块数量、最大斑块指数、景观形状指数、平均邻近度指数、结合度指数、分离度指数等6个景观指数,基于Fragstats 4.2软件,在景观水平上开展了不同气候情景下三尾凤蝶中-高适生区景观格局分析。
2 结果与分析
2.1 气候变化背景下三尾凤蝶适生区空间分布格局
2.1.1三尾凤蝶适生区预测模型的检验
MaxEnt模型在默认参数设置时,其RM=1,运行的特征组合有L,Q,H和P。而基于AICc信息准则构建的优化模型参数为RM=1、特征组合为L、Q和P(图1)。优化模型的AUCdiff低于默认模型(图1),表明优化模型可以避免物种数据过度拟合(朱耿平和乔慧捷,2016)。预测模型训练集和测试集的AUC值分别为0.953和0.919,两者均高于随机模型且相差不大,模型预测结果较可靠。
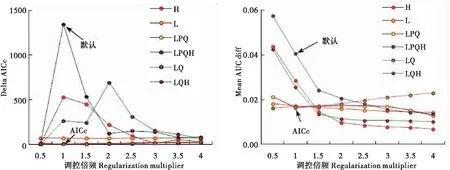
图1 不同参数下模型的平均AICc值和AUCdiff值
2.1.2当前气候条件下三尾凤蝶适生区预测
研究发现,三尾凤蝶在中国的适生面积为8.46×105km2,其中高适生区面积为1.52×105km2,占18.01%,中适生区面积为2.33×105km2,占27.50%,低适生区面积为3.69×105km2,占54.49%。三尾凤蝶在中国的适生范围为21.26°N~36.30°N、91.38°E~112.50°E,且集中于秦岭、大巴山、子午岭、黄龙山、横断山及其周边区域。高适生区主要分布于陕西秦岭、大巴山、子午岭、黄龙山,六盘山地区,甘肃东南部秦岭山脉向西延伸地带,河南省伏牛山地带,湖北省西部,四川盆地西北缘,云南省横断山地区等;中适生区大都沿高适生区向周边延伸分布,此外,云南省东北部、贵州省西北部、西藏藏南地区发现点状或面状的三尾凤蝶中适生区,低适生区则沿中适生区继续向周边延伸(图2)。
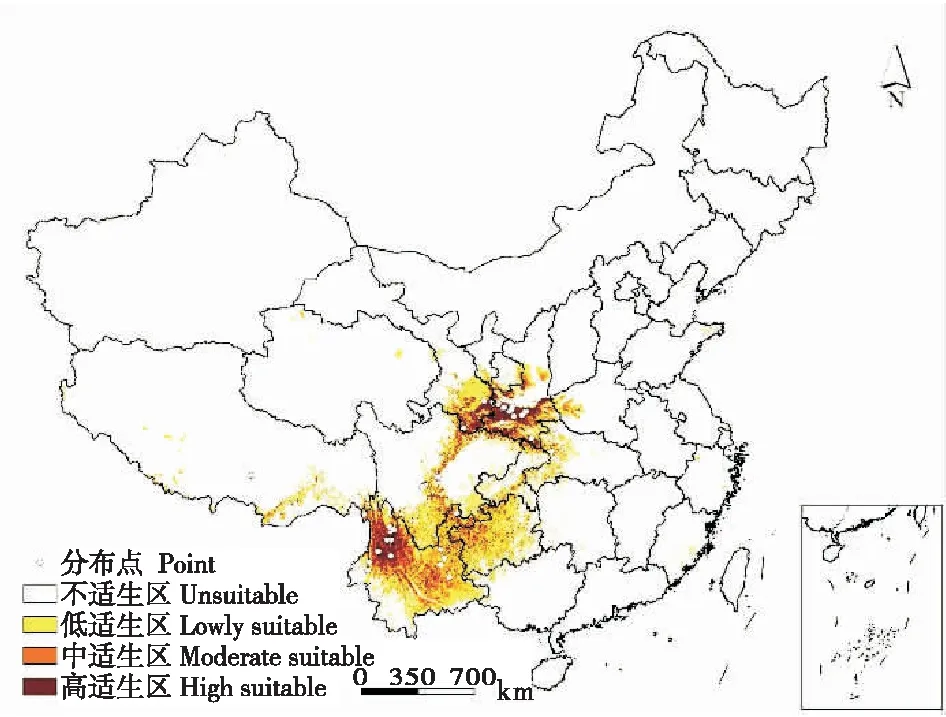
图2 当前气候条件下三尾凤蝶在中国的潜在适生区分布
比较三尾凤蝶实际记录点和MaxEnt预测的适生区划图发现,三尾凤蝶已知分布点外围仍有较大面积的三尾凤蝶适生区,其中陕西省宝鸡市陇山、子午岭地区、延安黄龙山地区、汉中南部大巴山地区,以及河南省伏牛山地带具有较高的三尾凤蝶适生区域(适生值≥0.4)。
2.1.3气候变化背景下三尾凤蝶适生区面积变化
与当前气候条件相比,当采取低辐射强迫可持续发展路径(SSPs 1~2.6)时,三尾凤蝶适生区新增2.04×104km2,新增区域集中于云南省东南部、贵州省大部、四川盆地西缘零星区域;三尾凤蝶适生生境丧失2.20×105km2,丧失生境主要位于云南省中部、湖北省西部、河南省西部、六盘山北侧、陕西省子午岭北侧和黄龙山北侧。三尾凤蝶适生区总面积缩减1.99×105km2,其中高适生区、中适生区、低适生区面积分别缩减5.65×104、8.34×104、5.93×104km2,面积缩减率依次为28.36%、41.88%、29.76%(表2、图3)。
当采取中等辐射强迫的中间发展路径(SSPs 2~4.5)时,三尾凤蝶适生区新增2.34×104km2,新增区域集中于云南省东南部、贵州省中部、四川盆地西北缘零星区域,三尾凤蝶适生生境丧失2.22×105km2,丧失生境主要位于云南省中部和东北部、贵州省西部、湖北省西部、六盘山北缘等。三尾凤蝶适生区总面积缩减1.98×105km2,其中高适生区、中适生区、低适生区面积分别缩减4.65×104、6.99×104、8.20×104km2,面积缩减率依次为23.44%、35.25%、41.31%(表2、图3)。
当采取区域竞争发展路径(SSPs 3~7.0)时,三尾凤蝶适生区新增2.71×104km2,新增区域集中于贵州省中部和东南部、四川盆地西缘零星区域;三尾凤蝶适生生境丧失2.67×105km2,丧失生境主要有云南省除横断山外的其它区域、贵州省西部、湖北省西部、河南省西部、六盘山北侧等;三尾凤蝶适生区总面积缩减2.40×105km2,其中高适生区、中适生区、低适生区面积分别缩减7.92×104、8.71×104、7.38×104km2,面积缩减率依次为32.99%、36.27%、30.74%(表2、图3)。
当采取高辐射强迫下的常规发展路径(SSPs 5~8.5)时,三尾凤蝶适生区新增1.48×104km2,新增区域零星分布于四川盆地西缘;三尾凤蝶生境丧失面积进一步增加,为3.39×105km2,丧失生境包括云南省除横断山外的其它区域、湖北省西部、河南省西部、秦岭-大巴山周边地区、六盘山北侧、陕西子午岭北侧,陕西黄龙山北侧;三尾凤蝶适生总面积缩减3.24×105km2,其中高适生区、中适生区、低适生区面积分别缩减0.89×104、1.07×105、1.28×105km2,面积缩减率依次为27.40%、33.02%、39.58%(表2、图3)。
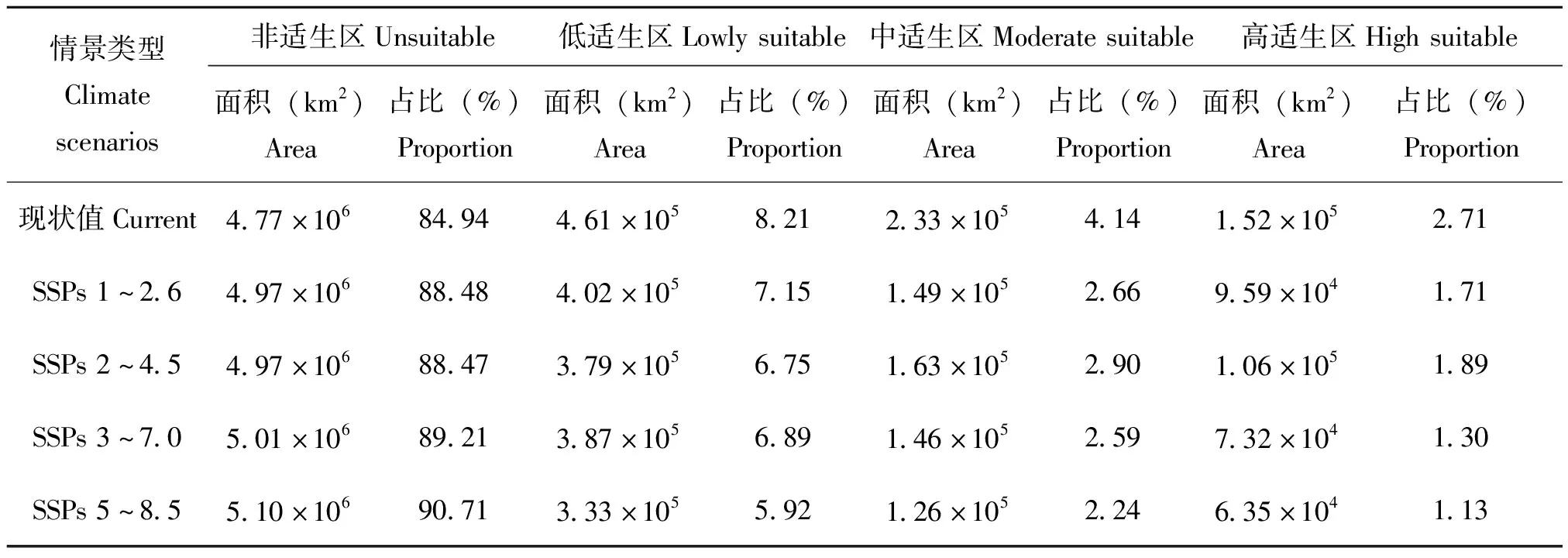
表2 气候变化背景下三尾凤蝶各适生等级面积
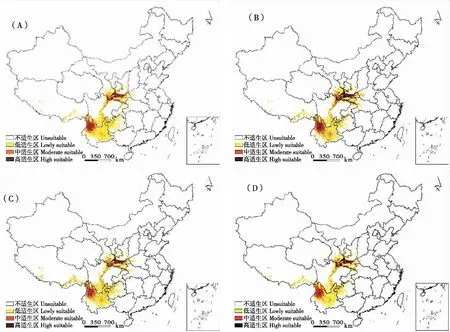
图3 未来不同情景下三尾凤蝶适生空间分布格局
2.2 三尾凤蝶适生区环境因子综合分析
基于MaxEnt模型中正规化训练增益的刀切法获得各变量对预测物种存在概率的相对重要性,结果表明,海拔、土地覆被对三尾凤蝶适生分布预测有重要影响,其次为年平均湿度、年平均温度、昼夜温差月均值、温度变化方差、湿度变化方差,坡度和坡向对三尾凤蝶适生分布预测影响较小(图4)。环境因子对MaxEnt模型的贡献率表明,土地覆被、年平均温、海拔是影响三尾凤蝶适生分布的最重要环境因子,贡献率依次为31%、18.17%、17.34%,其它环境因子对三尾凤蝶分布预测影响较小。
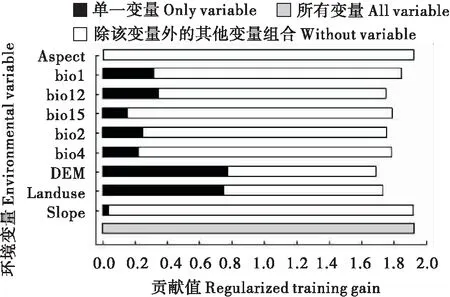
图4 刀切法检测环境因子对三尾凤蝶在中国分布的重要程度
为进一步明确气候变化背景下海拔对三尾凤蝶适生分布的影响,基于GIS技术提取了不同情境下三尾凤蝶适生区内的海拔范围和平均值,结果显示,在当前气候条件下,随着三尾凤蝶适生等级升高,区内海拔平均值大都呈升高趋势,中、高海拔区域更适合三尾凤蝶生存。在未来气候条件下,气候变暖进一步增强了三尾凤蝶适生区向高海拔区域迁移力度(表3)。
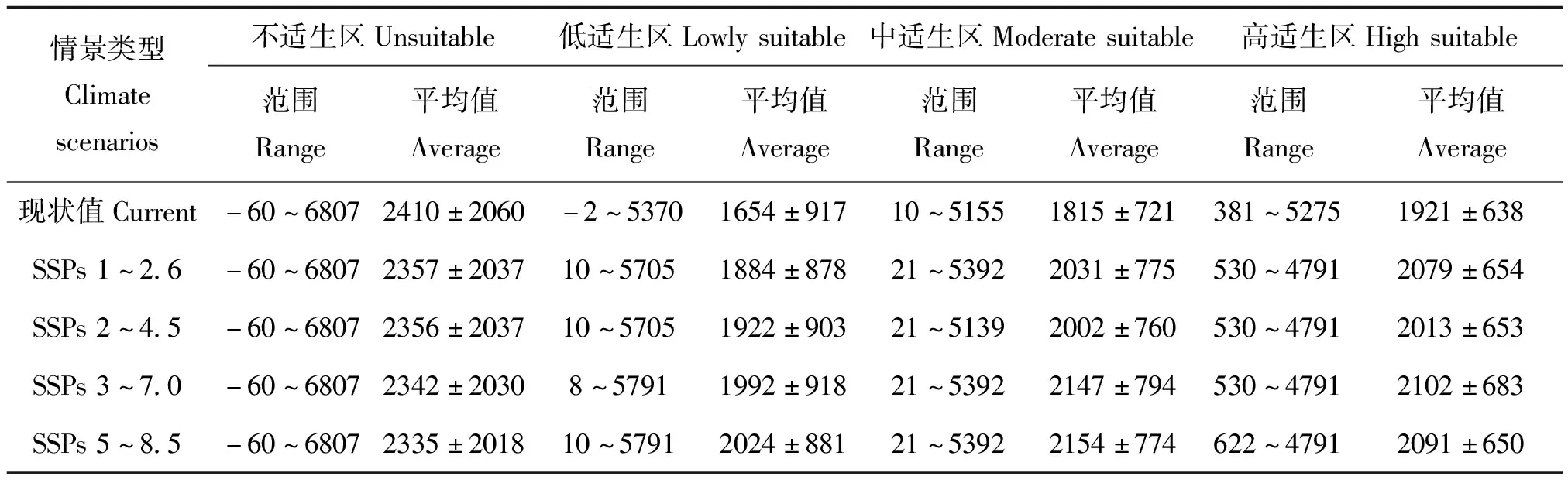
表3 气候变化背景下三尾凤蝶适生区内海拔变化(单位:米)
2.3 气候变化背景下三尾凤蝶适生区景观格局
基于Fragstats 4.2软件从全局角度对当前时期和2050年不同社会经济情景下三尾凤蝶适生分布的景观生态水平进行分析,结果表明,与当前时期相比,SSPs 1~2.6情景下三尾凤蝶中-高度适生斑块数量由3 155块降至1 312块,最大斑块指数由3.05降为1.37,景观形状指数由66.19降至43.22,平均邻近度指数由3 727.84降为262.63,斑块结合度由99.51降为98.95,分离度指数由761.66升至2 965.01。随着气候变暖,三尾凤蝶适生面积和斑块数量逐渐降低,最大斑块优势度减弱且斑块形状趋于简单化,斑块间相似度和结合度降低,斑块分离度增加,三尾凤蝶适生区域破碎化日趋严重(表4)。
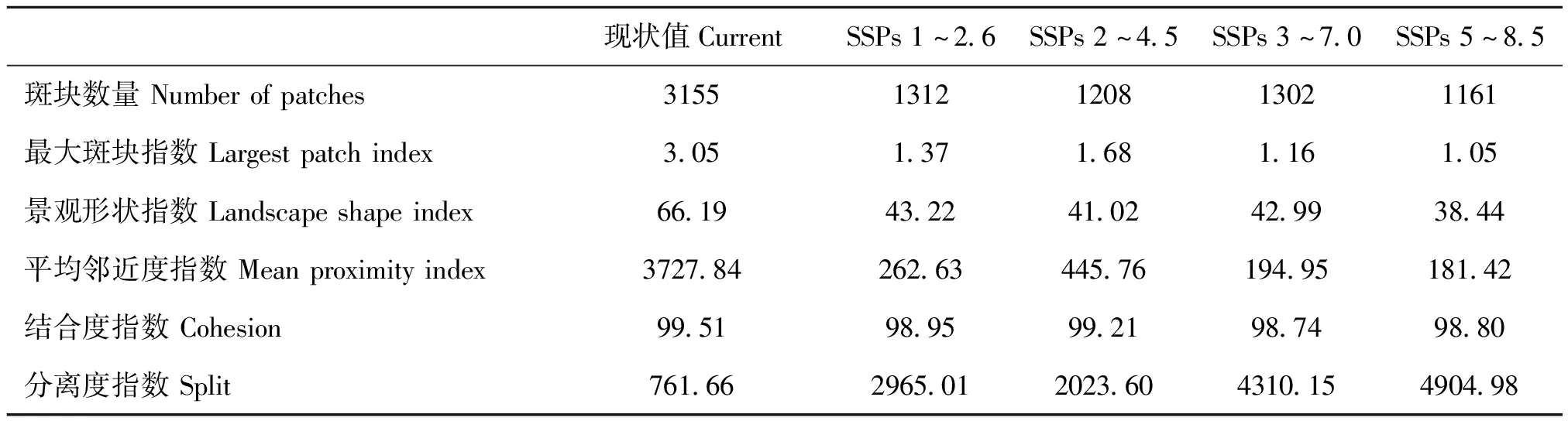
表4 气候变化背景下三尾凤蝶适生区内景观指数
2.4 气候变化背景下三尾凤蝶保护空缺分析
基于ArcGIS 10.6计算了中国428个国家级自然保护区与三尾凤蝶中-高度适生区二值图的重合比例,发现我国117个国家级自然保护区内存在三尾凤蝶中-高度适生区,小陇山、神农架等53个国家级自然保护区内三尾凤蝶中-高度适生面积比重过半。三尾凤蝶中-高度适生面积为3.85×105km2,受保护面积为1.94×104km2,94.97%的三尾凤蝶适生区域尚未得到有效保护。秦岭东段南麓、滇东北等地区存在较大面积的保护空缺。
假定我国三尾凤蝶受保护状况保持不变,2050年SSPs 1~2.6、SSPs 2~4.5、SSPs 3~7.0、SSPs 5~8.5等情景下适合三尾凤蝶生存的国家级自然保护区数量依次为89、94、87、79个,三尾凤蝶受保护面积依次为2.05×104、1.60×104、1.99×104、1.38×104km2。气候变化导致部分国家级自然保护区不再适合三尾凤蝶栖息,三尾凤蝶受保护现状进一步恶化。
3 结论与讨论
本研究基于MaxEnt模型,结合三尾凤蝶分布数据及BCC_CSM2_MR模式气候数据,预测了三尾凤蝶现在及未来的潜在分布格局。同时本文充分考虑高相关性的环境因子对生态模型模拟结果的影响(朱耿平等,2014),选择少量关键因子(Morrisonetal., 1998)进行模拟。结果表明三尾凤蝶适生范围较广,且集中于秦岭、大巴山、子午岭、黄龙山、横断山及其周边区域,这与Hu等人得出的三尾凤蝶适宜生存于滇西北、滇东北、贵州西北部、四川盆地西缘、秦岭等地区的结论基本一致(Huetal., 2019)。其它地区三尾凤蝶多呈点状或零星状分布,三尾凤蝶适生生境存在一定程度上的景观破碎化。尾凤蝶属昆虫是高山寡食性昆虫,寄主植物主要是马兜铃科植物(曾全等,2014),寄主植物种类单一且极易受人为干扰。三尾凤蝶幼虫主要分布于海拔1 600~2 280 m的林地空地、且具有马兜铃科植物分布的生境(Gaoetal., 2014),苛刻的野外生存环境以及人类干扰导致的生境破碎化或是导致三尾凤蝶濒危的主要成因。
蝶类物种因对温度变化敏感且能迅速作为响应,是无脊椎动物中颇受关注的类群和环境指示生物(Parmesanetal., 1999;Warrenetal., 2001;Nowickietal., 2008)。欧洲国家已开展大量关于气候变化与蝶类分布格局的研究(Parmesanetal., 1999;Pöyryetal., 2009)。本研究基于三尾凤蝶分布记录及我国国家气候中心开发的BCC_CSM2_MR模型数据,探讨了2050年气候变暖背景下三尾凤蝶适生格局变化,发现气候变暖越明显,三尾凤蝶适生面积越小。这与前人得出的气候变暖会加剧物种适生生境丧失的结论基本一致(Thomasetal., 2004)。在空间尺度上来看,不同地区的三尾凤蝶对气候变化的响应机制有着明显的不同。对比当前,当采取低辐射强迫可持续发展路径时,2050年气温增加幅度较低,横断山地区三尾凤蝶适生区略有北扩现象,秦岭和滇西北地区三尾凤蝶适生区明显收缩,贵州地区三尾凤蝶适生区存在东延现象,而随着气候变暖及增温加速,以上地区均明显向高海拔地区收缩,部分区域适生生境甚至丧失。一般来说,在全球变暖的大背景下,物种会向高纬度及高海拔地区迁移(Parmesanetal., 2003),蝴蝶向高海拔地区迁移速率更为明显(赵彩云等,2010)。本文结果表明海拔是影响三尾凤蝶地理分布的主要环境因子之一,气温增加越明显,三尾凤蝶适生区内平均海拔越高。部分中海拔地区因没有足够的高山环境供物种迁移而逐渐丧失适生生境(陈建国等,2011)。此外,由于动物和植物对气候变化的响应速率和适应能力不同步(Daviesetal., 2006),幼虫寄主植物较单一等特性不可避免的会对三尾凤蝶种群规模造成扰动,部分区域三尾凤蝶种群或面临灭绝。
生境丧失和破碎化或是物种退化、生物多样性衰退的重要原因(蒋志刚和葛颂,2005; Jantkeetal., 2011),自然保护区建设是保护生物多样性最有效的途径(薛达元等,1995)。当前我国三尾凤蝶中-高度适生面积为3.85×105km2,5.03%的适生面积位于国家级自然保护区内,绝大多数三尾凤蝶适生区域未得到有效保护,秦岭东段南麓、滇东北等区域仍存在保护空缺。半数以上具有三尾凤蝶适生区的保护区只注重旗舰物种的保护及森林生态系统保护,针对蝶类等无脊椎动物关注较少,无法满足三尾凤蝶在植被郁闭度、寄主植物等方面的生境需求。此外,我国自然保护区多呈“生态孤岛”式存在(郭子良等,2013),栖息地相对孤立不利于生物种群间基因交流(易传辉等,2011),进而造成三尾凤蝶遗传多样性下降或种群分化,现有保护政策无法避免气候变化导致的三尾凤蝶生境破碎化进程。针对越来越严重的三尾凤蝶适生生境丧失和破碎化威胁,建议相关部门在自然保护地机构改革和自然保护地边界调整的大背景下,综合考虑濒危蝶类及其寄主植物、访花植物生物学特性,调整保护边界,并在保护地之间布设生态廊道和生境踏石,保障三尾凤蝶等濒危蝶类栖息地完整性和连续性。同时多渠道开展迁地保护、就地保护、保育等保护措施,减缓气候变化背景下三尾凤蝶等濒危蝶类的灭绝风险。
猜你喜欢
西部林业科学(2022年6期)2023-01-03
中国中医药信息杂志(2022年7期)2022-07-17
中国野生植物资源(2022年3期)2022-04-19
福建农林大学学报(自然科学版)(2022年1期)2022-01-18
中国农村水利水电(2021年11期)2021-11-29
小学生必读(低年级版)(2021年4期)2021-07-28
风景园林(2021年2期)2021-04-24
幼儿100(2021年7期)2021-04-10
落叶果树(2021年6期)2021-02-12
小学生必读(低年级版)(2020年11期)2020-03-03
