
菊苣EST-SSR分子标记开发及通用性分析
2021-10-19 13:51梁小玉胡远彬白史且张新全
草地学报 2021年9期
梁小玉, 季 杨, 胡远彬, 易 军, 白史且, 张 靓, 张新全
(1.四川省畜牧科学研究院, 四川 成都 610066; 2. 四川农业大学动物科技学院, 四川 成都 611130;3. 四川省林业和草原局, 四川 成都 610082)
菊苣(Cichoriumintybus)为菊科多年生宿根草本植物,在畜牧业、食品业、医药业以及工业上广泛应用,是当今世界上极具潜力的经济作物之一。目前,国内外对菊苣的研究热点集中在繁育、品种培育、食品营养、化学成分、药理以及牧草等方面[1],种质资源评价与新品种选育对开发利用菊苣资源越来越重要。但是,缺乏相关菊苣基因组信息,使我们对其遗传信息了解较少,目前所使用的相关序列扩增多态性(Sequence related amplified polymorphism,SRAP),随机扩增多态性DNA(Random amplified polymorphic DNA,RAPD),扩增片段长度多态性(Amplified fragment length polymorphism,AFLP)等分子标记也难以满足种质资源评价、后代精准鉴定辅助育种等工作的需要,在一定程度上限制了菊苣种质资源的开发利用。因此,大量开发新的分子标记对于菊苣种质资源辅助育种具有重要的意义。
SSR标记具有共显性遗传、多态性高、易检测、特异性好、通用性高及重复性好等优点,是目前动、植物遗传育种中应用最多的一种分子标记[2-5],SSR按来源可分为基因组SSR和表达序列标签SSR (EST-SSR)。EST-SSR具有基因组SSR的特征,能够直接获得基因表达的信息,为功能基因提供“绝对”的标记,其多态性可能与基因功能直接相关[6-9],在近缘物种甚至远缘物种间通用性更好,且开发成本较低[10-13]。本研究基于美国国家生物技术信息中心(National center for biotechnology information,NCBI)数据库公布的菊苣EST序列分析其所含微卫星重复序列的组成和特征,开发菊苣EST-SSR引物,并验证菊苣EST-SSR引物有效性及其在莴苣属(LactucaL.)和苦荬菜属(IxerisCass.)中的通用性,以期为菊苣及其近缘属种的亲缘关系鉴定、系统进化、遗传变异、遗传图谱构建和功能基因鉴定与克隆等研究提供重要的工具和有价值的信息。
1 材料与方法
1.1 植物材料与DNA提取
引物有效性验证采用的是来源于美国国家基因库的5份菊苣材料,来源及形态差异均较大,通用性评价采用的是31 份莴苣亚族(LactucinaeLess.)材料,其中,7份苦荬菜属和1份莴苣属材料(序号20)来源于中国国家牧草种质基因库,其余23份莴苣属材料来源于四川农业大学草学系(见表1)。5~6叶期时,每份种质选择表型一致的20 个单株的幼嫩叶片等量混合成100 mg,采用新型植物基因组DNA提取试剂盒(天根,北京)提取DNA(每份材料3个重复)。利用0.8%琼脂糖凝胶电泳和紫外分光光度计检测DNA浓度和纯度,合格的DNA稀释成10 ng·μL-1置于-20 ℃冰箱内保存备用。
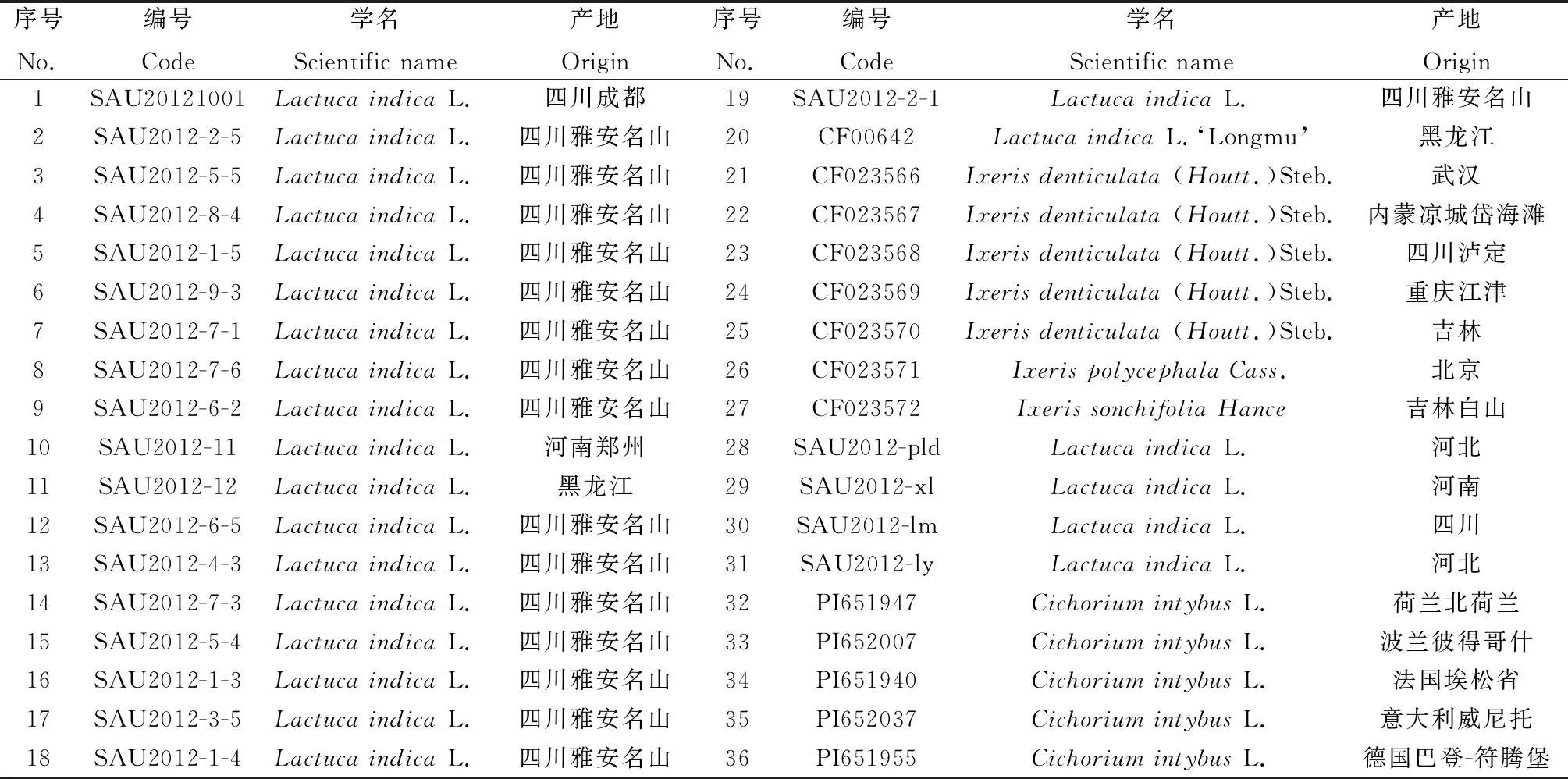
表1 供试材料来源
1.2 菊苣EST-SSR引物设计
1.2.1菊苣EST-SSR序列下载与SSR位点搜索 利用DNAstar软件对来源于NCBI数据库的53 975条菊苣EST序列(截止2020年12月)进行拼接和聚类。应用简单序列重复识别工具(Simple sequence repeat identification tool,SSRIT)在线查找菊苣无冗余EST序列中的SSR位点。搜索标准为SSR长度≥18 bp,二、三、四、五、六核苷酸的序列数目分别至少重复9,6,5,4,4次。
1.2.2SSR引物设计 根据SSR侧翼的保守区域,应用Primer premier 5.0 软件设计引物。引物设计参数为:熔解温度 50℃~60℃,最适复性温度55℃,上下游引物间温度差异≤3℃;预期扩增产物大小90~500 bp;引物长度18~22 bp,预期扩增率≥80%;引物GC含量40%~60%,最适GC含量50%,上下游引物间GC含量差异≤5%;尽量避免错配、发卡结构、引物二聚体出现。引物由北京六合华大基因科技股份有限公司合成。
1.3 PCR扩增
聚合酶链反应(Polymerase chain reaction,PCR) 扩增体系为15 μL,包括1 μL DNA,0.3 μL酶,引物各1 μL,混合液7.5 μL,水4.2 μL。反应程序:94℃5 min;94℃30 s,熔解温度40 s(每循环降低1℃),72℃1 min,5个循环;94℃30 s,Tm -5℃30 s,72℃1 min,35 个循环;72℃延伸10 min后4℃保存。PCR扩增产物经6.0%变性聚丙烯酰胺凝胶电泳(电泳缓冲液1×TBE,电压200 V,时间4 h)。电泳完成后银染,最后用数码相机拍照保存。
1.4 统计方法
SSR发生频率为含SSR的Uni-EST数量与无冗余总EST数量之比,SSR出现频率为SSR数量与无冗余总EST数量的比值,SSR平均分布距离为无冗余EST数量的总碱基数与SSR数量的比值。根据PCR产物扩增情况,选取清晰易辨的条带进行统计分析,有带记为1,无带记为0,构建0/1二元数据矩阵,计算多态性位点百分率、引物多态性信息含量(Polymorphism information content,PIC)[14]。利用NTsys-pc 2.10e软件中的similarity程序计算材料间遗传相似性系数(Genetic similarity,GS),基于clustering程序中的SHAN进行聚类分析,再根据Graphics程序中的tree plot绘制亲缘关系树形图。
2 结果与分析
2.1 菊苣EST-SSR的信息分析
2.1.1菊苣EST中SSR的数量与分布特点 由表2可知,53 975 条菊苣EST拼接和聚类后获得40 188 条Unigenes,序列总长30 723.73 kb,从中共搜索到648 个SSR位点。SSR发生频率为1.54%,出现频率为1.61%,平均每47.41 kb出现1个SSR。菊苣转录组中SSR类型丰富,且各类型出现数量和分布距离的差异较大,其中,三核苷酸、二核苷酸和六核苷酸重复类型占主导地位,分别占SSR位点总量的46.45%,30.56%和19.13%。SSR平均长度24.15 bp,为中等偏长的序列,其中,长度18~20 bp和21~25 bp的SSR在总SSR中占比较高,分别达到43.84%和36.64%。
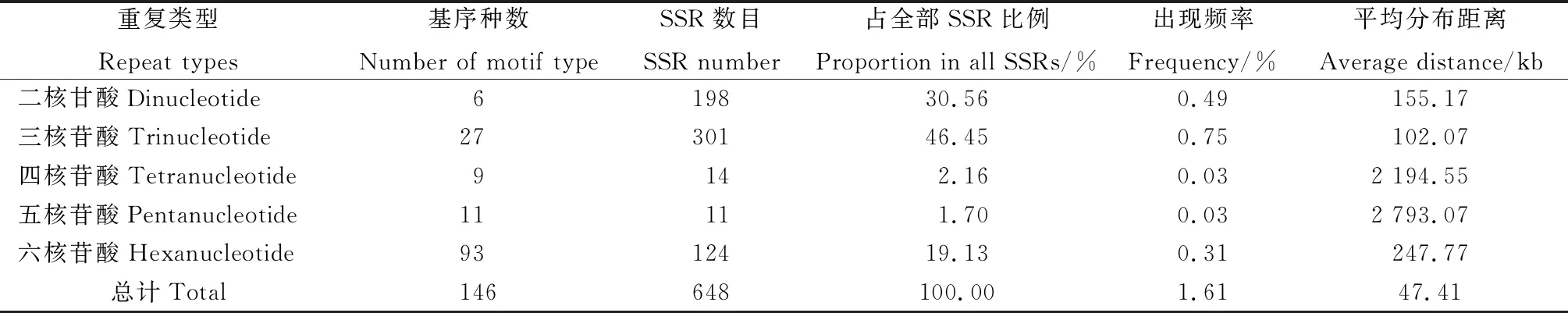
表2 基于重复基元大小的 EST-SSR 分布
2.1.2菊苣EST-SSR基序重复类型和特征 菊苣EST-SSR核苷酸基序类型较丰富,648 个SSR中共搜索到146 种重复基序,六核苷酸类型中的重复基序种类最多,达到93 种,占基序类型总数的63.70%(表2)。出现频率最高的6种基序类型主要存在于二核苷酸和三核苷酸(表3),分别为:TC/GA,AG/CT,TTC/GAA,ATC/GAT,ATG/CAT和TGA/TCA,六核苷酸每种基序类型出现频率均很低。不同重复类型SSR基序的重复次数差异很大,范围在4~29 次间,其中,二核苷酸基序重复次数跨度最大,在9~29 次均有分布,三核苷酸基序有10 种重复次数,主要分布在6~20 次。重复次数基本上随重复基序的核苷酸数目的增多而减少,且不同核苷酸重复基序的主导重复次数不同,三核苷酸重复6次的占主导,达26.397%,六核苷酸重复4次的占主导,为16.36%,二核苷酸重复7次的占主导,为11.42%(图1)。
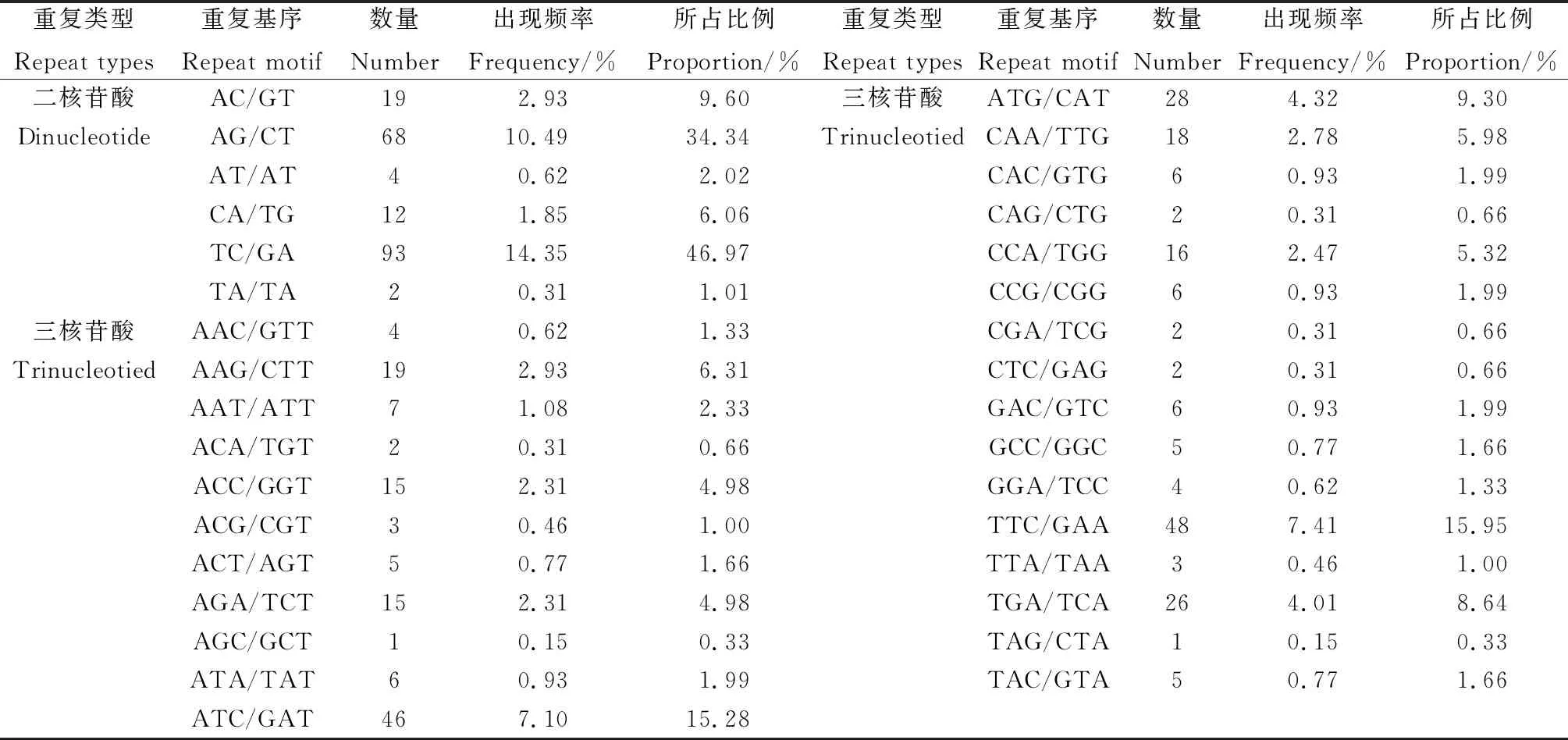
表3 菊苣EST中二核苷酸和三核苷酸重复基序特点
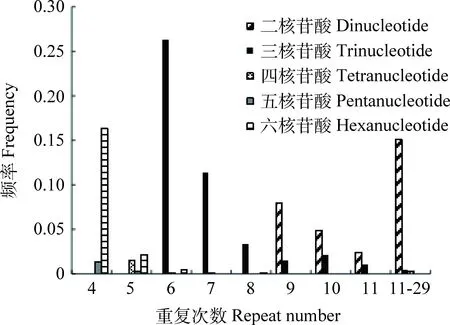
图1 菊苣EST中不同SSR重复基序的重复次数分布
2.2 菊苣EST-SSR引物筛选与检测
为了初步验证设计的菊苣EST-SSR引物的有效性,从345 对引物中选择152 对评分95 分以上的引物,以5份菊苣材料(表1)的基因组DNA为模板进行PCR扩增。152 对引物中有119 对引物能够有效扩增,实际扩增率78.29%,与预期值接近。其中,87 对引物扩增片段长度与预期接近,90 对引物能扩增出清晰条带,87 对引物在5个材料间检测出多态性,多态率为57.24%,多态性引物信息见表4。已合成的119 对引物中,含有二、三、五和六核苷酸重复的引物分别有20,37,4和58 对,含六核苷酸重复类型的引物最多,没有四核苷酸重复类型。
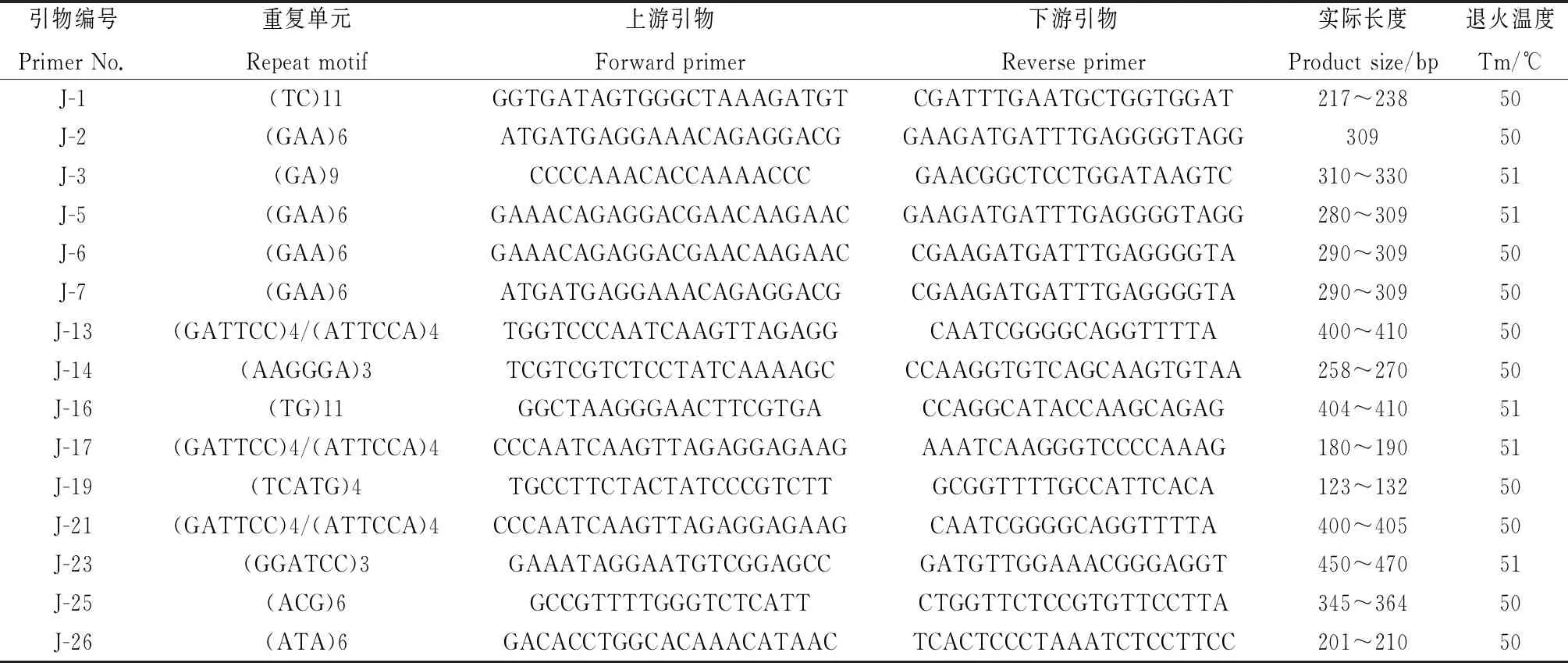
表4 引物序列及信息
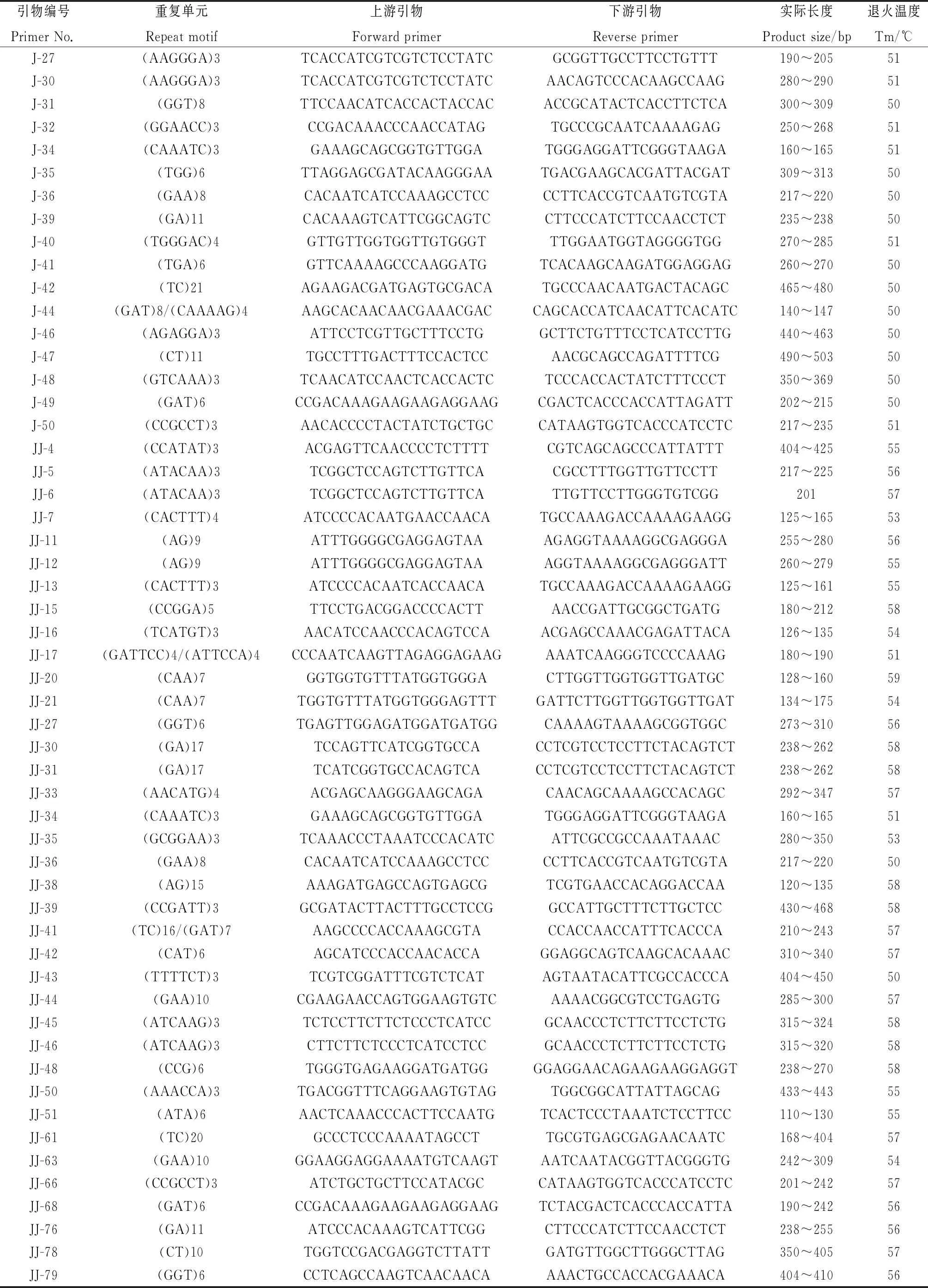
续表4
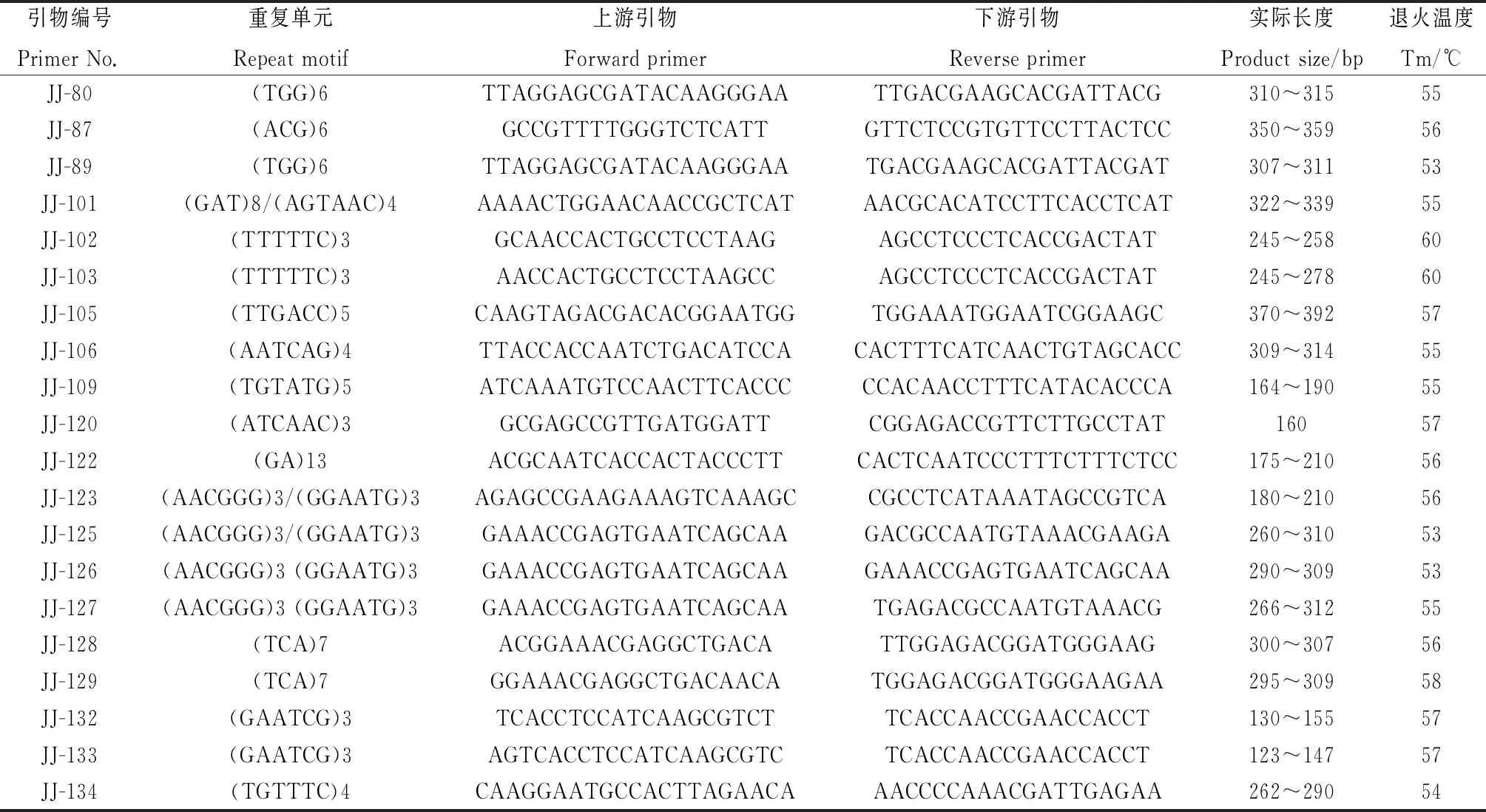
续表4
2.3 菊苣EST-SSR引物通用性评价
2.3.1菊苣EST-SSR对近缘属材料扩增产物多态性分析 在87 对具有多态性的菊苣EST-SSR引物中随机挑选出43 对引物,以31 份莴苣亚族材料(表1)的基因组DNA为模板进行PCR扩增,37 对引物可以有效扩增,转移扩增效率达到86.05%。挑选扩增出清晰条带的33 对引物进行统计分析,由表5可知,共扩增出599 条清晰条带,每对引物扩增条带数范围在8~26 条,平均每对引物扩增18.2 条条带,引物JJ4和JJ134(见图2)扩增条带数最多,均为26 条。其中,多态性条带584 条,平均每对引物扩增出17.7 条多态性条带,多态位点在85.7%~100%间,平均97.1%。引物扩增PIC值在0.12~0.34间,平均0.21,PIC值最高的为J19。整体看,33 对引物对供试材料有较高扩增效率,参试材料种质间变异较大,存在较丰富的遗传多样性。

图2 菊苣EST-SSR引物JJ-134的扩增图谱
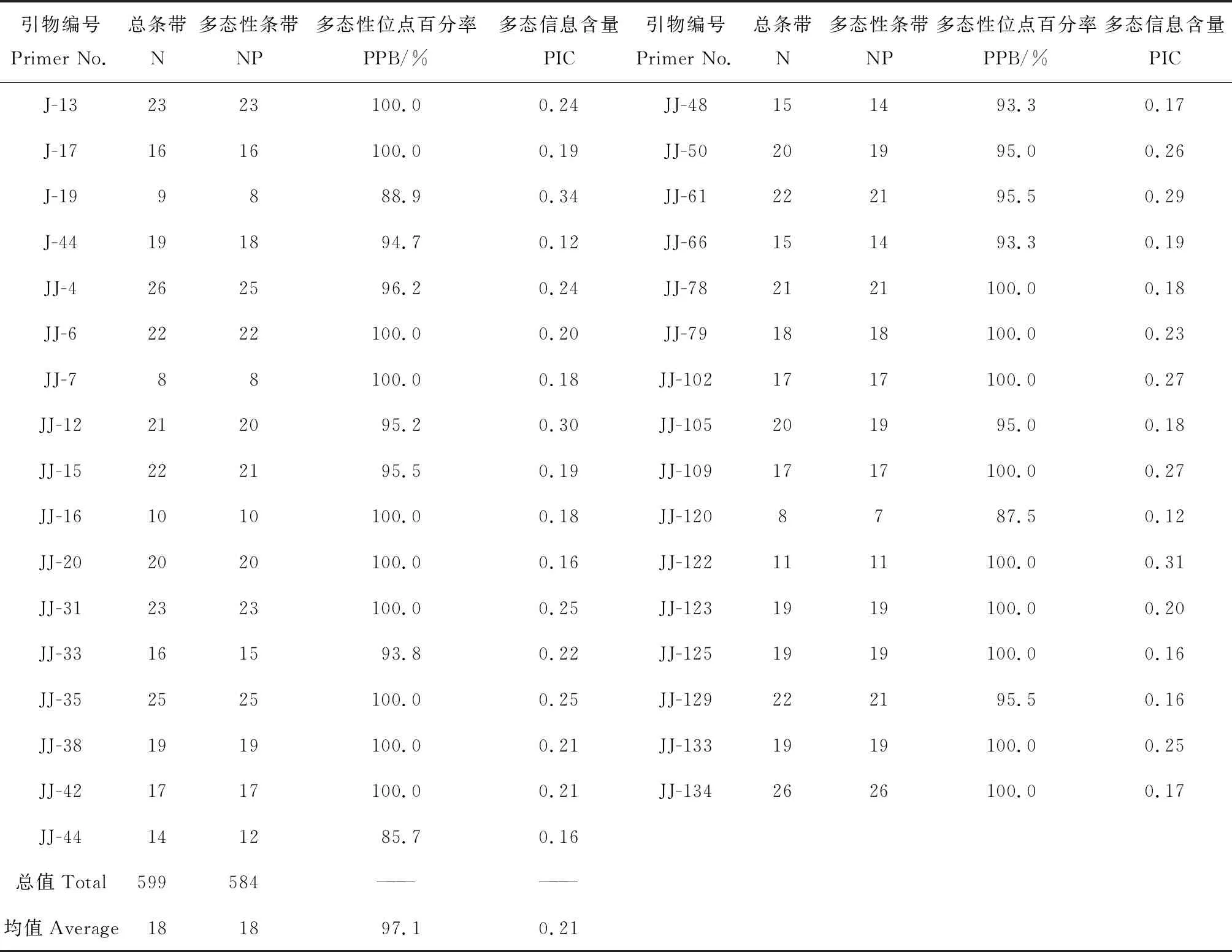
表5 菊苣EST-SSR引物组合对苦荬菜和莴苣扩增结果及多态性
2.3.2基于菊苣EST-SSR聚类分析 遗传相似系数(GS)结果显示,供试材料之间差异明显,具有较为丰富的遗传多样性。31 份近缘种质间的成对GS值在0.472~0.938间,平均GS值为0.76,变幅为0.466,其中,来自四川雅安的2份材料SAU2012-2-5和SAU2012-5-5间GS值最大为0.94,其遗传距离最小为0.472,表明其亲缘关系最近,来源于四川雅安的SAU2012-2-5和内蒙的CF023567间GS值最小,其遗传距离最大,表明其亲缘关系最远。7份苦荬菜属种质间的成对GS值在0.49~0.874,平均GS值为0.618,23 份莴苣属种质间的成对GS值在0.760~0.938,平均GS值为0.861,表明参试苦荬菜属的遗传多样性高于莴苣属的。在遗传相似系数分析的基础上,应用非加权类平均聚类方法对31 份材料进行聚类分析,构建亲缘关系树状图(图3)。31 份供试材料在遗传相似系数0.822处可以划分为6个组群。其中,24 份莴苣属材料全部聚在第Ⅰ组,第Ⅰ组群又包含2个亚组,第1亚组包括17 份四川的、2份黑龙江的以及1份河南的,而序号为28~31的4份莴苣属材料聚在第2亚组。7份苦荬菜属材料分别聚在5个组群内,第II组包含2份苦荬菜属材料,即CF023568和CF023569,第III组、第IV组和第VI组均只有1份苦荬菜属材料,分别为:CF023571,CF023566和CF023572。第V组也包含2份苦荬菜属材料,分别为内蒙的CF023567和吉林的CF023570。
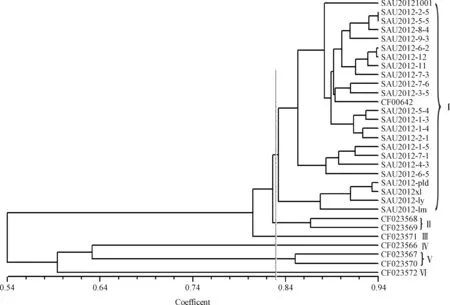
图3 苦荬菜和莴苣的UPGMA聚类图
3 讨论
随着测序技术的飞速发展,大量快速增长的EST数据已成为EST-SSR标记开发的重要来源,Cordeir等[15]推测,EST文库中大约2%~11%的EST序列中含有SSR,而不同植物物种的EST中有1%~5%能发展成SSR标记[16]。截止2020年12月,NCBI数据库中公布的菊苣EST序列为53 975条,与水稻(Oryzasativa)(1 369 506条)、小麦(Triticumaestivum)(1 358 140条)、大麦(HordeumvulgareL.)(537 546条)等禾本科(Poaceae)作物相比,菊苣EST序列数据明显不足[17]。本研究中菊苣SSR出现频率为1.61%,略高于玉米(ZeamaysL.)的1.5%[16],但远低于沙打旺(Astragalusadsurgens)的25.85%[18]、苏丹草(Sorghumsudanense)的 21.75%[19]、不结球白菜(Brassicarapassp.chinensis)的34.84%[20]和水稻的27%[21]等。这种差异与物种、SSR开发工具、搜索标准及EST数量、质量、来源等因素密切相关[22]。如开发水稻EST-SSR时,搜索SSR的长度标准由12 bp增加到30 bp时,SSR的出现频率从50%减少到1%[23],而在本研究中,SSR长度标准≥18 bp,搜索标准较为严格。
大量研究结果表明,大多数植物的EST-SSR以二、三核苷酸重复类型为主[24-25],如沙打旺[18]、藜麦(ChenopodiumquinoaWilld.)[25]、猕猴桃(Actinidia)[26]等以二核苷酸重复类型为主,本研究中,菊苣与燕麦(AvenasativaL.)、苏丹草、甘蔗(Saccharumofficinarum)等一样,以三核苷酸重复类型为主[17,19]。Gao等[27]发现,许多三核苷酸与具有重要功能的基因相关。四、五核苷酸重复类型很少,与大多数研究一致。值得关注的是,本研究中六核苷酸重复类型所占比率较高,达到19.13%,表现出明显的偏倚性,这在所报道的植物种类中极少见,如枣(Ziziphusjujuba)[24]、苏丹草[19]、不结球白菜[20]等多在2%以下,但与李新凤等[28]的研究结果相似,他们认为这种现象可能与编码区域由框移突变引起的非三核苷酸SSR受抑制有关。
此外,主导重复在不同物种间也存在差异[7],Rota等[23]对水稻、小麦、大麦基因组和EST序列中SSR的分布和频率研究发现,二、三、四、五核苷酸重复基序类型分别有4,10,33和102 种。本研究中,菊苣二核苷酸至五核苷酸重复基序类型分别有6,27,9和11 种,优势重复基序类型为TC/GA (14.35%),与孙清明等[29]研究结果类似。二核苷酸重复基序出现的重复次数类型多、跨度大,如AG/CT和TC/GA分别有27种、23种,重复次数类型跨度从重复9次到27 次、29 次,Cho等[30]认为,SSR的重复次数与其变异呈正相关。菊苣重复基序出现情况及其频率高低均表现出明显的偏倚性,可能与不同物种进化水平、基序的基因表达程度或突变频率等因素的差异有关[31],也可能与菊苣现有的EST数较少、覆盖度不足有关。
利用EST设计引物的有效扩增率一般在60%~90%[32]。本研究中菊苣EST-SSR引物有效扩增率为78.29%,表明通过菊苣EST开发SSR的效率较高,开发的引物具有良好的特异性。部分引物未能扩增可能是所设计引物跨越mRNA剪切位点以及扩增产物包含的内含子太长所致。用于苦荬菜属和莴苣属材料通用性检测分析的菊苣EST-SSR引物扩增结果显示,PIC值均低于0.5,按照Botstein[33]对多态性的定义,均为低度多态性标记。但是,班骞等[34]采用本研究开发的菊苣EST-SSR引物构建苦荬菜指纹图谱时,PIC值均高于0.5,平均达到0.834。这种差异与不同参试材料及来源差异、引物扩增情况等有关,也与引物序列的选择密切相关,如引物核苷酸重复类型与次数、基序类型以及SSR的长度,当SSR长度≥20 bp时多态性较高[35],而且低级基序SSR多态性普遍比高级基序的高[36]。本研究中筛选合成引物标准为评分95 分以上的,基序为六核苷酸的引物则接近总数的50%,随机挑选用于分析的33 对引物中,基序为六核苷酸的占比达55%,而班骞挑选引物时并未考虑综合评分,所用引物以二、三核苷酸为主,且基序重复次数较高,增加重复次数,也相应的增加了多态性的概率[31,34-35]。
大量研究表明,EST-SSR侧翼序列在物种之间高度保守,具有较好的科、属、种间通用性,亲缘关系越近,扩增的谱带越清晰,而且转移性比基因组SSR更高[34,36-37]。本研究获得类似结果,菊苣EST-SSR稳定性和重复性好,扩增的谱带清晰,转移率较高,达到86.05%,表明菊苣EST-SSR在莴苣亚族中的通用性较强。31 份近缘种质间的遗传相似系数在0.472~0.938间,表明参试种质资源蕴含着较丰富的遗传多样性。聚类分析显示,开发的菊苣EST-SSR能够有效鉴别莴苣属和苦荬菜属,参试材料地理来源与聚类的一致性较为明显,表现出较明显的地域分布规律。部分地域差异较大的材料聚在同一组,可能发生了人为或者天然基因交流。CF023571,CF023566及CF023572等3份材料分别聚在第III组、第IV组和第VI组,与3份材料来源地域差异较大,且为不同种的野生型苦荬菜密切相关,表明它们不仅与莴苣属材料间亲缘关系较远,而且种间亲缘关系也较远,有明显的种间遗传差异。通用性验证结果显示,新开发的菊苣EST-SSR标记通用性较强,可以对菊苣及其近缘属遗传多样性以及属、种间亲缘关系进行有效的鉴定。
4 结论
本研究基于53 975 条菊苣EST序列搜索到648 个SSR位点,成功设计了345 对引物,并筛选出119 对评分95 分以上的有效引物,其中37 对引物通用性较强,种间转移扩增率达86.05%。此外,苦荬菜属和莴苣属种质的遗传多样性初步分析显示,供试材料具有较丰富的遗传多样性,聚类划分与材料地理来源有相关性,不同种间存在明显的遗传分化。这为菊苣族分子标记开发、种质资源评价、系统进化研究及分子标记辅助育种奠定了重要基础。
猜你喜欢
南方医科大学学报(2022年3期)2022-04-13
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10
中国预防兽医学报(2020年8期)2020-11-05
三农资讯半月报(2020年15期)2020-08-25
儿童时代·幸福宝宝(2019年9期)2019-10-28
红领巾·萌芽(2017年2期)2017-03-09
云南畜牧兽医(2014年2期)2014-02-28
新疆农垦科技(2014年2期)2014-02-28
中国蔬菜(2012年1期)2012-01-27
健康之家(2009年5期)2009-05-26
