
川金丝猴家域的季节性差异
2021-10-12 05:30董鑫郎嘉钰楚原梦冉赵姗姗张晋东白文科
生态环境学报 2021年7期
董鑫 ,郎嘉钰,楚原梦冉,赵姗姗,张晋东,白文科,
1. 西华师范大学环境科学与工程学院,四川 南充 637009;2. 西南野生动植物资源保护教育部重点实验室,四川 南充 637009;3. 高黎贡山国家级自然保护区保山管护局,云南 保山 678000;4. 西华师范大学管理学院,四川 南充 637009;5. 西华师范大学生态研究院,四川 南充 637002
家域是指在一定时间内,特定动物出现的一定概率值(例如95%)的区域范围(Kernohan et al.,2001),是动物个体或者群体进行取食、交配和育幼所利用和活动的区域(Burt,1943)。由于家域分布是开展野生动物栖息地质量、负载量、食性选择、种群生存等野生动物生态学研究的重要基础参数,因此成为野生动物种群生态学的重要研究内容之一(李言阔等,2010)。常用的家域估算方法有最小凸多边形法(Minimum convex polygon,MCP)、核密度估计法(Kernel density estimate,KDE)等(Kernohan et al.,2001;Laver et al.,2008;张晋东等,2013;段延,2018;袁耀华等,2019)。其中,核密度估计法是基于动物活动点位,利用空间分布的非参数估计来估算家域的一种方法,能有效解决个别边缘点位对家域分布范围的影响(Kernohan et al.,2001;Laver et al.,2008);同时,核密度估计法能有效区分动物对不同空间资源的利用强度,如通过等值线方法确定家域利用强度的空间分布格局(白文科等,2017a),是目前应用较多的家域估算方法(Powell,2000;张晋东等,2013)。
川金丝猴(Rhinopithecus roxellana)属树栖灵长类,其分类地位以及种群结构都具有明显的自身特征,是森林物种的典型代表之一。由于其移动速度快,活动范围大,空间利用上具“分散—聚合”现象(胡锦矗等,1980)。此前已有学者对金丝猴家域的季节性变化进行了相关研究,指出了川金丝猴的活动范围分布具有季节性差异(李保国等,1994;李阳,2010;顾志宏等,2011;李沂韦,2016;郭家强,2018;唐书培,2019)。其中,季节性的食物资源分布被认为是影响川金丝猴家域分布与生境利用的一个主要因素。如,有学者对神农架川金丝猴栖息地的选择中的季节差异进行了讨论,指出季节性植物生长发育程度导致了不同季节川金丝猴家域利用强度差异(李阳,2010);而家域与食性选择的研究指出不同季节川金丝猴活动区域的季节性变化与食物的可获得性密切相关(李沂韦,2016)。
随着空间信息技术在景观生态学的应用,特别是ArcGIS与Fragstats综合运用的方法逐渐成熟,利用景观指数来反映区域景观格局的结构组成和空间配置特征得到广泛应用(邬建国,2007;王永等,2017;赵行双,2018;李江荣等,2021)。但有关物种家域的景观格局分析研究较少。因此,为了解川金丝猴的家域的季节变化,以及家域是否景观尺度上存在破碎化,本研究以四川省九寨沟县白河国家级自然保护区下坪地一带为研究区域,选取保护区下坪地一带的川金丝猴群(约250只)为研究对象,结合景观生态学方法和家域模型,探究川金丝猴家域空间分布、生境利用的季节性差异,以及家域的景观格局变化特征,为保护区针对性管理和保护提供科学参考。
1 研究方法
1.1 研究区域概况
白河国家级自然保护区始建于 1963年,位于四川省阿坝藏族羌族自治州九寨沟县境内(104°02′—104°12′E,33°10′—33°22′N),是以大熊猫、川金丝猴等珍稀野生动物及森林生态系统为主要保护对象的自然保护区(图1)。保护区南北长约18.5 km,东西宽约14.8 km,总面积约162.04 km2,其中林地覆盖面积约128.35 km2,占保护区总面积的79.2%。保护区内山高谷深,海拔高差约3000 m左右;植被丰富繁茂,受地形和气候变化形成了落叶阔叶林(<2700 m)、寒温性针叶林(2500—3600 m)、温性针叶林(2200—2600 m)、灌丛(>2700 m)、草甸(>3200 m)、高山流石滩稀疏植被(>3700 m)等6种主要植被类型(金贵祥等,2015;李艳忠等,2016;李瑜等,2020)。区内川金丝猴最多时超过1500只,至少有3群川金丝猴,以地名区分分别是下坪地猴群、油房沟猴群、薛家坪猴群,且三群猴群分布距离较远,是国内川金丝猴分布密度最大、最具有代表性的保护区之一(Chu et al.,2018;Dong et al.,2019;顾海军等,1998;李艳忠等,2016;金贵祥等,2020)。

图1 四川省白河国家级自然保护区地理位置及研究区域Fig. 1 Geographical location and study area of Baihe National Nature Reserve in Sichuan province
1.2 数据来源与处理
以四川省白河国家级自然保护区种群数量最多的下坪地川金丝猴群(约250只)作为研究对象,实地调查获取了2011年3月—2013年9月的川金丝猴GPS活动位点。从观测日清晨(06:30—07:00)从营地出发开始寻找猴群,发现猴群时一般进行全天观察,直到猴群在傍晚(18:30—19:30)进入夜宿地为止。通常在距猴群30—50 m处,每隔30 min用全球卫星定位系统(GPS)记录一次猴群活动的位置、海拔、坡向等信息。如果因天气或地形的影响,发现猴群距离较远(>100 m),无法记录猴群活动的准确位点时,尽量通过到与猴群活动区域的相同海拔高度处记录位点;或在地形图上标记猴群活动的大体位置后,条件允许时再回到标记位置补记位点的方法进行弥补。将获得的川金丝猴活动位点(n=568)和保护区边界导入地理信息系统 ArcGIS 10.5软件中,发现有部分金丝猴活动点超出了保护区范围,由此增加了自然保护区外围缓冲区(1 km)一并作为本文的研究区域(图1)。
选取保护区范围Landsat系列遥感影像,利用遥感软件ENVI 5.5对其进行预处理,进行监督分类获取保护区内覆被类型。数字高程模型(Digital elevation model,DEM)来源于地理空间数据云(http://www.gscloud.cn/)。国界、省界矢量数据来源于国家基础地理信息中心,全国地理信息资源目录服务系统公开发布的1꞉100万全国基础地理数据库(https://www.webmap.cn/commres.do?method=result 100W),保护区边界及其保护站点等数据来自白河国家级自然保护区管理局。
1.3 家域估算与利用强度等级划分
家域是野生动物空间分布及生境利用等生态学研究的基础概念,其估算模型已得到了长足的发展,但不同模型得出的结果差距较大。其中,最小凸多边形法(MCP)是最早应用的(Burt,1943;Kernohan et al.,2001;Seaman et al.,1999;Laver et al.,2008),依据外围位点生成家域,计算简单,能较直观地描述研究动物整个活动区域(Powell,2000;Kernohan et al.,2001),但其忽视了内部位点的空间信息,无法准确反应动物对不同空间资源利用的强度(Powell,2000;Kernohan et al.,2001;Laver et al.,2008)。栅格单元计数法由于不能有效计算家域面积,近年来应用相对较少(Harris et al.,1990)。核密度估计法(KDE)是一种概率模型方法,利用空间分布的非参数估计估算家域能较好地区分动物对不同空间的利用强度,可以通过等值线来确定家域大小和形状(Powell,2000;Kernohan et al.,2001;Laver et al.,2008),被一些学者认为是最合适的家域估算模型而广泛使用。本研究选择了核密度估计法(KDE)来估算川金丝猴不同季节的家域分布。模型建立过程中,选择合适的平滑指数(即h统计量)是KDE法估算家域的最关键步骤(Worton,1995)。如果高估h值会产生更大且紧密的家域闭合曲线,导致估算结果偏大;而h值过小又会产生很多小的、离散的家域闭合曲线(张晋东等,2013)。因此,在建立不同季节KDE模型时,我们对比了最优平滑指数(href)法(Worton,1995)、最小平方交叉验证法(LSCVh)和似然交叉验证法(CVh)(Horne et al.,2006)3种平滑指数的估算结果。根据川金丝猴日移动距离大、活动范围广、活动中心多、点位样本量及时间跨度大等特点,且避免出现更大且紧密的家域闭合曲线或过多碎小离散的闭合曲线,最后选取了LSCVh值为建模参数的估算结果来分析川金丝猴不同季节家域与生境利用强度。
基于ArcGIS 10.5软件平台,利用Home Range Tools(家域分析工具)建立KDE模型,以获取调查区域川金丝猴群的家域分布(Rodgers et al.,2015)。通过模型参数对比,选择了最小平方交叉验证法(LSCVh)值为家域模型建立的平滑指数(Horne et al.,2006),对不同季节川金丝猴家域进行了分别估算。其中,季节划分为春季:3—5月、夏季:6—8月、秋季9—11月、冬季12至次年2月。
在模型中设置25%、50%、75%、95%的利用强度等值区间,将得到的不同季节家域模拟结果,设定 0—25%区域为核心生境(S1),25%—50%区域为次核心生境(S2),50%—75%区域为一般利用生境(S3),75%—95%为边缘生境(S4),0—95%区域为家域总面积(S0)。
1.4 家域的空间分布特征
以不同季节川金丝猴的家域分布情况为分析对象,在季节家域(S0)和利用强度生境(S1,S2,S3,S4)两个水平上,利用景观分析软件 Fragstats 4.2对川金丝猴家域的面积指标:斑块类型面积(CA)、斑块数(NP);形状指标:面积加权平均形状指数(AWMSI);聚散性指标:蔓延度指数(CONTAG)、景观分割指数(DIVISION)、分离度指数(SPLIT)景观格局指数进行计算分析。指标的计算公式及其生态学意义见表1(McGarigal et al.,1995;Wu et al.,2002)。

表1 选取的景观格局指数及其生态学意义Table 1 Landscape pattern indexes and ecological meanings used in the study
1.5 生境因子利用特征
将得到的不同季节川金丝猴家域图层在ArcGIS 10.5软件支持下,和海拔、坡度、坡向以及植被类型4个生境因子图层相叠加(武瑞东,2006;白文科等;2017b),得到白河自然保护区川金丝猴不同季节家域的生境因子分布图;再通过空间分析工具统计不同季节家域在各生境因子分布区间的面积。同时,获取各季节川金丝猴活动位点的生境因子特征值(海拔、坡度、坡向),利用SPSS 20.0软件进行不同季节生境因子特征值的单因素方差分析,以检验川金丝猴季节生境利用的是否存在差异(Zhang et al.,2006);再对生境因子特征值均数进行多重比较(Tukey)分析,以比较川金丝猴不同季节之间的生境利用差异特征。其中,显著性水平均设置为0.05。
2 结果与分析
2.1 不同季节的家域分布
由图2知,该川金丝猴群的整体利用区域较为一致,主要集中在保护区东南的下坪地区域。不同季节家域虽在空间分布上有差异,但不存在季节间的整体迁移现象,表明白河自然保护区该川金丝猴群具有稳定的活动区域。另外,该川金丝猴家域分布的最东缘超出了保护区的边界范围(图2),需在保护工作中予以一定关注。

图2 白河国家级自然保护区下坪地川金丝猴各季节家域利用空间分布Fig. 2 The home range of golden monkeys in different seasons in Baihe National Nature Reserve
2.2 不同季节家域的空间分布格局
2.2.1 面积指标特征
在季节家域(S0)水平的斑块面积上,川金丝猴在春、夏、秋、冬4个季节的家域总面积分别为14.19、15.16、25.17、4.94 km2(表2)。其中,冬季的家域面积明显小于其他季节,随着春季气温回暖,家域范围开始逐渐扩大,到食物资源丰富的秋季,其家域面积也达到最大。斑块数量上,春季、夏季和冬季的斑块数分别为5个、5个和4个(表2,图 2),其家域分布具有一定的不连通性;而秋季的家域范围完全融合(图2),表明川金丝猴秋季家域利用的空间分布更为均衡,连通性更高。
在等值区间利用强度生境(S1,S2,S3,S4)水平上,随着家域生境利用强度等级的降低,各季节川金丝猴的家域面积均逐渐增大(表2)。秋季的家域在空间上较其他季节面积大,并有更多的核心区域(S1,S2),但斑块数目较少,表明川金丝猴的秋季家域破碎化程度较低。冬季核心家域面积最小,另外,夏季各利用强度生境的斑块数量均为最大值,其中S3水平上相互分离的斑块有16个,表明夏季家域具有最高的破碎化程度(表2,图2)。
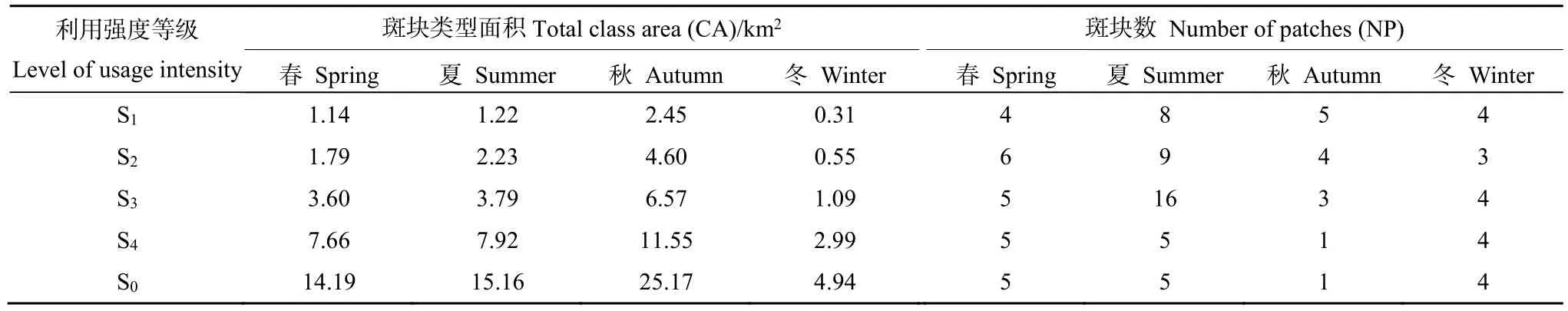
表2 川金丝猴不同季节家域面积和斑块数Table 2 The home range area of golden monkeys and patches number in different seasons
2.2.2 形状指标特征
在季节家域(S0)水平上,春、夏、秋、冬4个季节家域的面积加权平均形状指数(AWMSI)逐渐降低。其中,春、夏季较为接近,秋季家域形状趋于简单的原因是其斑块为相互连通的整体,而冬季家域面积明显小于其他季节且空间分离程度较大,导致其家域形状更趋于简单(图3)。
在等值区间利用强度生境(S1,S2,S3,S4)水平上,随着家域生境利用强度等级越低,面积加权平均形状指数(AWMSI)总体上逐渐变大(图3),这表明在川金丝猴家域的实际利用中不同季节的斑块的形状都是由核心区域向外呈现更复杂的趋势,具有明显的边缘效应。
2.2.3 聚散性指标特征
由于研究对象是一群川金丝猴,且其日移动距离远、活动范围较大,使得春、夏、秋季的蔓延度指数值相近,而只略大于冬季(表3),表明其冬季家域利用的空间分布较其他季节稍有分散。

表3 川金丝猴家域聚散性变化特征Table 3 Aggregation variation characteristics of Sichuan golden monkeys in different seasons
景观分割指数(DIVISION)和分离度指数(SPLIT)一定程度反映着不同季节川金丝猴家域斑块间的空间连通性和破碎化程度。夏季川金丝猴家域的景观分割指数和分离度指数较春季有所增大,推断其原因是随着气温回升,而可利用食物资源分布不均匀,导致川金丝猴家域在空间分布出现了一定分离。冬季,由于气温低及食物资源匮乏使川金丝猴的移动强度明显降低,导致其家域利用的空间分布也最为分散。
2.3 不同季节家域的生境因子特征
2.3.1 海拔因子
不同季节家域海拔区间分布差异显著(P<0.05)。由表4可知,不同季节川金丝猴家域分布的主要海拔区间为 2000—3200 m,除冬季外,所占家域总面积均在90%以上(图4)。其中,秋季家域的海拔区间最为集中,虽其家域总面积最大(25.17 km2),但2000 m以下和3200 m以上的家域面积均为 4个季节的最小值,仅为 0.14 km2和0.24 km2;而冬季家域的海拔区间分布最分散,2000 m以下和3200 m以上区域的面积达到季节家域面积的14.54%(表4)。

图4 川金丝猴各季节家域海拔区间分布图Fig. 4 Elevation distribution map of Sichuan golden monkeys’s home range in different seasons

表4 金丝猴不同季节家域海拔区间的面积分布Table 4 Elevation distribution of Sichuan golden monkeys in different seasons
2.3.2 坡度因子
由表5可知,不同季节川金丝猴家域在所有坡度区间均有分布,且季节间各坡度区间家域面积所占比例基本一致,除春季和秋季之间(P<0.05),其他季节坡度因子的利用均无显著差异(P>0.05),这主要是由该川金丝猴群所利用区域的地形地貌条件决定的。该区域川金丝猴各季节家域分布所集中的坡度区间为10°—50°,面积占比均大于90%,而家域利用分布在 10°以下和 50°以上坡度的区域面积较小(表5)。

表5 白河保护区川金丝猴不同季节家域不同坡度区间的面积分布Table 5 Slope distribution of Sichuan golden monkey’s home range in different seasons in Baihe National Nature Reserve
2.3.3 坡向因子
由表6可知,不同季节川金丝猴家域在所有坡向区间均有分布,且季节间各坡向区间家域面积所占比例基本一致,不同季节坡向因子的利用均无显著差异(P>0.05)。各季节家域利用所集中的坡向区间为 0—60°、180°—360°,合计面积占比约 90%。其中,秋季家域的坡区区间分布较其他季节更为平均,其原因是秋季食物资源的可获得性较强,川金丝猴选择的随机性更大;而冬季家域的坡向区间选择差异最大,推断其原因是冬季气温较低,川金丝猴家域明显缩小,坡向区间的面积分布受其范围内地形条件的影响。

表6 川金丝猴不同季节家域不同坡向区间的面积分布Table 6 Aspect distribution of Sichuan golden monkey’s home range in different seasons
2.3.4 植被类型
由图5可以看出,不同季节川金丝猴的家域分布主要集中在林区,植被类型以针阔混交林和针叶林为主,合计面积占比约99%;而落叶阔叶林、灌草丛、裸岩等不适合川金丝猴生存的植被类型面积极小。究其原因,这主要是由研究区植被的海拔垂直地带性分布决定的,同时也受到气温及阳光照射的影响,如冬季家域分布中的落叶阔叶林和裸岩的占比较其他季节更高(图5)。

图5 川金丝猴各季节家域植被类型分布图Fig. 5 Vegetation types distribution map of Sichuan golden monkey’s home range in different seasons
3 讨论
白河国家级自然保护区川金丝猴种群密度大,分布有大面积的针阔混交林,食物资源丰富,大面积的适宜栖息地为川金丝猴的生存及种群增长提供了有利条件。基于 3S技术的生境质量评价研究显示,保护区川金丝猴种群的生境质量经历了由较好转变为退化,然后又逐渐转好的过程(李艳忠等,2016)。而近年来,随着保护力度的加大,人为干扰逐步减少,保护区内川金丝猴数量持续增加,成为了川金丝猴生存的理想区域(金贵祥等,2020)。随着保护区内川金丝猴等野生动物栖息地的逐步转好,使得保护区内更多同域分布的野生动物种群提供了生存空间,生物多样性得到了较好的保护。然而,对该保护区内川金丝猴家域的季节性变化少有报道,尚需进一步研究。故本研究基于KDE模型,结合地面调查数据,通过模型及参数的对比,有效提高了川金丝猴家域分布与生境利用季节性差异分析的科学性。为白河自然保护区川金丝猴种群保护提供参考的同时,也实践了多种指标及分析手段在野生动物家域及生境利用分析中的应用。
本研究中,基于 KDE模型的不同季节家域空间分布变化显示了川金丝猴群的季节性“分离—聚合”现象,主要表现为不同季节间川金丝猴的核心生境斑块的数量变化和空间迁移,与诸多学者的研究结果较为一致(胡锦矗等,1980;金贵祥等,2020)。下坪地川金丝猴群在不同季节家域的分布范围虽然有差异,但季节间的整体家域分布相互重叠,表明该川金丝猴群家域的整体分布稳定,并不存在季节间的整体迁移现象。然而,由于全球气候变化及人为干扰将会使川金丝猴的栖息地区域破碎化,种群间连通性下降,进而威胁其正常的生存繁衍。一般来说,稳定的生存繁衍区域是其种群增长的基础,在持续的跟踪调查中,下坪地川金丝猴群的估计数量也的确有所增加(李沂韦,2016;金贵祥等,2020)。包括本文的诸多研究均揭示了下坪地区域的生态环境可以满足川金丝猴不同季节的生境选择需求,也体现了保护区建立与野生动植物资源保护的重要作用。而本研究中不同季节间栖息地利用强度(S1,S2,S3,S4)空间面积及分布格局的差异明显,具体表现为具有多核心家域(聚合)和扩散边缘家域(分离)的种群分布现象,明确解释了川金丝猴家域分布及生境利用特征;也进一步表明,除受研究区域本身地形等条件限制外,其家域利用主要是受气温及食物资源分布影响,这也与其他学者的研究结果相一致(李沂韦,2016)。
本研究综合了家域内部的空间利用强度,基于景观格局指数分析了川金丝猴家域空间分布的季节性差异。研究结果显示,该川金丝猴群家域利用的季节性差异不仅表现在家域面积的大小上,其内部利用强度的空间格局差异也是川金丝猴家域季节性特征的主要表现形式。同时,通过不同季节生境利用特征的比较,揭示了川金丝猴群季节性“分离—聚合”过程中的栖息地选择规律,解释了不同季节川金丝猴家域分布与生境利用的驱动因子。研究结果证明了川金丝猴家域空间分布的景观格局指数变化趋势与其季节性生境选择偏好密切相关,如秋季气温高、食物资源丰富,川金丝猴的家域面积最大,空间分布的连通性也最高;而冬季气温低、食物资源匮乏导致其家域面积缩小、空间分布也更加分离。
存在的不足:由于保护区地形复杂,野外川金丝猴群跟踪难度大,本研究只跟踪了下坪地区域的一个川金丝猴群,忽略了邻近猴群空间竞争对其家域分布与生境利用的影响。另外,人类活动一直被认为是影响野生动物栖息地选择的主要原因之一,而白河自然保护区下坪地区域川金丝猴群距离村庄较近,由于缺少微生境的实地调查,未能获取人类活动,如采药、放牧等人为干扰因子信息,可能对其家域分布及生境利用特征的分析有一定影响。
4 结论
本研究基于 KDE家域分析模型和景观格局分析工具,构建了四川省白河国家级自然保护区下坪地川金丝猴群在不同季节的家域模型和生境利用空间格局。得出的主要结论如下:
(1)研究区川金丝猴家域(90%以上)主要分布在海拔2000—3200 m,坡度10°—50°,坡向区间为0—60°、180°—360°的针叶林和针阔混交林中,但各季节生境利用面积占比却有所不同。
(2)冬季的川金丝猴的家域面积明显小于其他季节,由于气温低及食物资源匮乏使川金丝猴的移动强度明显降低,导致其家域利用的空间分布也最为分散。随着春季气温回暖,家域范围开始逐渐扩大,至秋季达到最大家域面积。同时,秋季家域破碎化程度较低,生境利用的空间分布融合,连通性最高;夏季家域破碎化程度高,夏季与冬季家域的连通性较低。
(3)该川金丝猴群的整体生境利用区域较为一致,主要集中在保护区东南的下坪地区域。不同季节家域虽在空间分布上有差异,但不存在季节间的整体迁移现象,表明白河自然保护区该川金丝猴群具有稳定的活动区域。川金丝猴家域分布的最东缘超出了保护区的边界范围,需在保护工作中予以一定关注。
猜你喜欢
品牌研究(2021年7期)2021-11-28
云南画报(2021年10期)2021-11-24
东方少年·布老虎画刊(2021年1期)2021-06-22
落叶果树(2021年6期)2021-02-12
小哥白尼(野生动物)(2020年4期)2020-07-27
建材发展导向(2019年11期)2019-08-24
学生天地(2018年36期)2019-01-28
现代园艺(2018年2期)2018-03-15
现代园艺(2018年1期)2018-03-15
体育科技(2016年2期)2016-02-28
