
新疆博格达峰南坡岩面生地衣群落特征的研究∗
2021-10-10 00:45:46买吾拉江衣沙克艾尼瓦尔吐米尔
新疆大学学报(自然科学版)(中英文) 2021年5期
买吾拉江·衣沙克,艾尼瓦尔·吐米尔
(新疆大学 生命科学与技术学院,新疆 乌鲁木齐 830046)
0 引言
地衣是由真菌和藻类或者蓝细菌共生而形成的一种特殊的生物类群,但近年来有关地衣共生体的研究取得了进展,研究人员发现地衣是一类包括多种微小生物种类的微生态系统,地衣体内除了藻类和真菌外还包括很多种微小的生物类群[1,2].地衣体内的这些特殊的微小生物类群作为地衣产生次生代谢物的主要来源,使地衣能够广泛的分布和适应各种极端栖息地环境[1,2].据报道地衣对环境的适应性极强,一般其它植物种类处于生理极限的高山、北极、南极和沙漠环境中地衣都能够生长,据预测地球表面超过10%的陆地生态系统已被地衣覆盖[3].岩面生地衣(saxicolous lichen)是指能够生长在岩石环境,地衣体紧贴于岩石表面的壳状、枝状和叶状地衣种类.在干旱地区广泛的分布在光照强烈、高山低温等条件较差的裸露的岩石表面,主要以壳状地衣种类为主形成地衣群落.分布在岩石上面的地衣种类通过分泌地衣酸等次生代谢物质腐蚀岩石、改进栖息地微环境、促进土壤的形成,为其它植物种类的侵入、定居以及形成植物群落提供条件,促进植物群落的原生演替速度,与此同时为多种昆虫提供食物和栖息环境[4,5].
国外学者从16世纪开始在地衣分类学、区系地理、地衣生态学等方面开展相关研究.自20世纪以来在北美和欧洲,研究人员对树附生、岩面生、地面生地衣群落的结构、物种分布与环境间因子间的关系、地衣群落动态、地衣群落构建机制以及对全球变化的响应等方面进行了深入和系统的研究[6,7].在地衣群落结构及群落构建机制方面,Koch等研究了附生地衣功能群多样性和性状聚集格局对热带雨林演替阶段的指示作用,并探讨了附生地衣群落的构建[8].Prieto等采用群落谱系和功能性状相结合的方法,对山毛榉森林中的附生地衣群落构建机制进行了对比研究.研究发现,分布在北部的样方中附生地衣群落谱系和功能性状表现出发散,群落的构建主要与种间关系的限制有关,相反在南部的样方中群落的构建表现出谱系和功能性状的聚集,主要是与环境的过滤作用有关.因此,他们认为群落谱系和功能性状相结合的研究方法在地衣群落构建机制中具有潜在的研究意义[9].Benitez等采用地衣功能性状,对厄瓜多尔热带山地森林砍伐后的附生地衣群落的构建机制进行了研究,认为森林砍伐后环境条件的变化导致了地衣功能性状的变化,从而影响了附生地衣群落构建及其结构[10].Degtjarenko等在爱沙尼亚北部研究了碱尘污染地区附生地衣功能性状的影响和群落物种分布格局[11].在岩面生地衣群落方面的研究发现岩面生地衣种类的分布与岩石的种类、岩石的pH、坡度、坡向、栖息地的湿度、光照强度、人为干扰和空气污染等多种因素有关[12−15].Purvis等报道岩面生地衣群落的种类多样性受到它们分布的岩石理化性质的限制[16].Puy Alquiza等研究墨西哥Guanajuato市两座历史建筑表面的岩面生地衣与岩石关系时,发现岩面生地衣的假根穿透岩石表面0.5 μm至50 μm深处,导致岩石表面矿物质的分解,并破坏岩石表面微环境而影响岩面生地衣群落的种类组成[17].
国内已有学者对树附生、地面生、朽木生地衣群落进行系统的研究.李苏等持续对云南哀牢山区的原始森林及次生林开展研究,采用定量研究方法对附生地衣物种多样性与分布特征;附生大型地衣的物种组成、生物量和物种多样性;附生地衣盖度、多样性和群落特征与中国西南地区亚热带森林的演替和环境变化间的关系;附生地衣与森林类型和宿主树木特征的关系;亚热带常绿阔叶林附生地衣凋落物的物种多样性和生物量的边缘效应;干热胁迫对蓝藻地衣固氮酶活性的影响;中国西南地区亚热带森林中非优势树种对附生地衣物种丰富度的增强作用等进行了大量的研究[18−23].王玉良等采用主成分分析法对鹞落坪国家自然保护区树生地衣和地面生地衣进行了群落排序,应用Spearman秩相关检验对地衣进行了种间关联分析,以地衣多样性、相似性和均匀度等指数,对各种地衣群落进行了比较[24,25].李英英等对沿海热带常绿季雨矮林中树干附生地衣的分布规律进行了研究,采集并记录季雨矮林中群落重要值前20的树种及树干附生地衣的相关数据,对地衣与宿主胸径、种类间的关系,地衣在宿主上的分布高度、方位朝向及相关环境因子间的关系进行分析[26].柳帅等研究了亚热带山地森林附生地衣的移植生长及其对环境变化的响应[27].但有关岩面生地衣群落方面研究报道较少,我们近年来在乌鲁木齐南部山区、阿尔泰山两河源自然保护区、托木尔峰国家自然保护区、博格达山北坡等地区对岩面生地衣群落进行了定量研究,积累了一定的野外研究经验,并初步查明了影响岩生地衣物种分布的因素[28−33].博格达峰南坡的自然环境与北坡具有明显的差异,南坡较干旱,植被覆盖率较低,自然环境比较脆弱.岩面生地衣群落是干旱环境的优势群落,具有高度的环境适应能力,研究并查明该群落的物种多样性,找出影响地衣群落种类组成及物种分布的因素,有效保护地衣资源,对评价干旱区自然生态环境稳定性方面具有重要的意义.
1 研究区概况和研究方法
1.1 研究区概况
博格达峰南坡地区气候属于中温带大陆性干旱性气候.全年平均气温7°C,冬夏冷热悬殊,昼夜温差大.年平均降雨量71.7 mm.蒸发强烈,干燥少雨是主要的气候特征.该区地形地貌复杂,既有高低落差大而陡峭的丘陵、洪积扇,也有狭长平坦的河谷、冰峰与湖泊.博格达峰南坡的植被带类型从低到高可以分为:海拔1 100~1 600 m的山地荒漠带,海拔1 800~2 400 m的山地草原带,海拔2 400~2 900 m的亚高山草甸带和海拔2 900~3 400 m阴坡的高山草甸带[34,35].
1.2 研究方法
在博格达峰南坡的达坂城区西沟乡、东沟乡、阿克苏乡(黑沟村、黑沟冰湖、高崖子牧场)等地的不同海拔选择6个具有代表性的样地.在6个样地中设立20 m×20 m的样点20个,每个样点的四边和中间设置5 m×5 m的样方5个,采集样方中的岩面生地衣(采集岩面生地衣标本时,在样方内尽量选择表面积大于1 m×1 m的岩石)并计算地衣的盖度.计算地衣盖度时,在样方中直径大于1 m的每一岩面上设立面积为50 cm×50 cm的铁筛置于小样方上,铁筛用细丝划分成2.5 cm×2.5 cm的小格400个,调查时计测每种地衣在铁筛网格线交叉处出现的次数,估测岩面生地衣的盖度[28,31].同时用数码相机拍下每个样点中的地衣外部形态,记录每个样点的海拔高度、经纬度、湿度、光照强度、坡向、人为干扰程度等信息,并且带回基物(岩石,土壤)样品,以便测量理化性质.20个样点的环境因子原始数据都经过最大值标准化处理,并分为5个等级(表1).
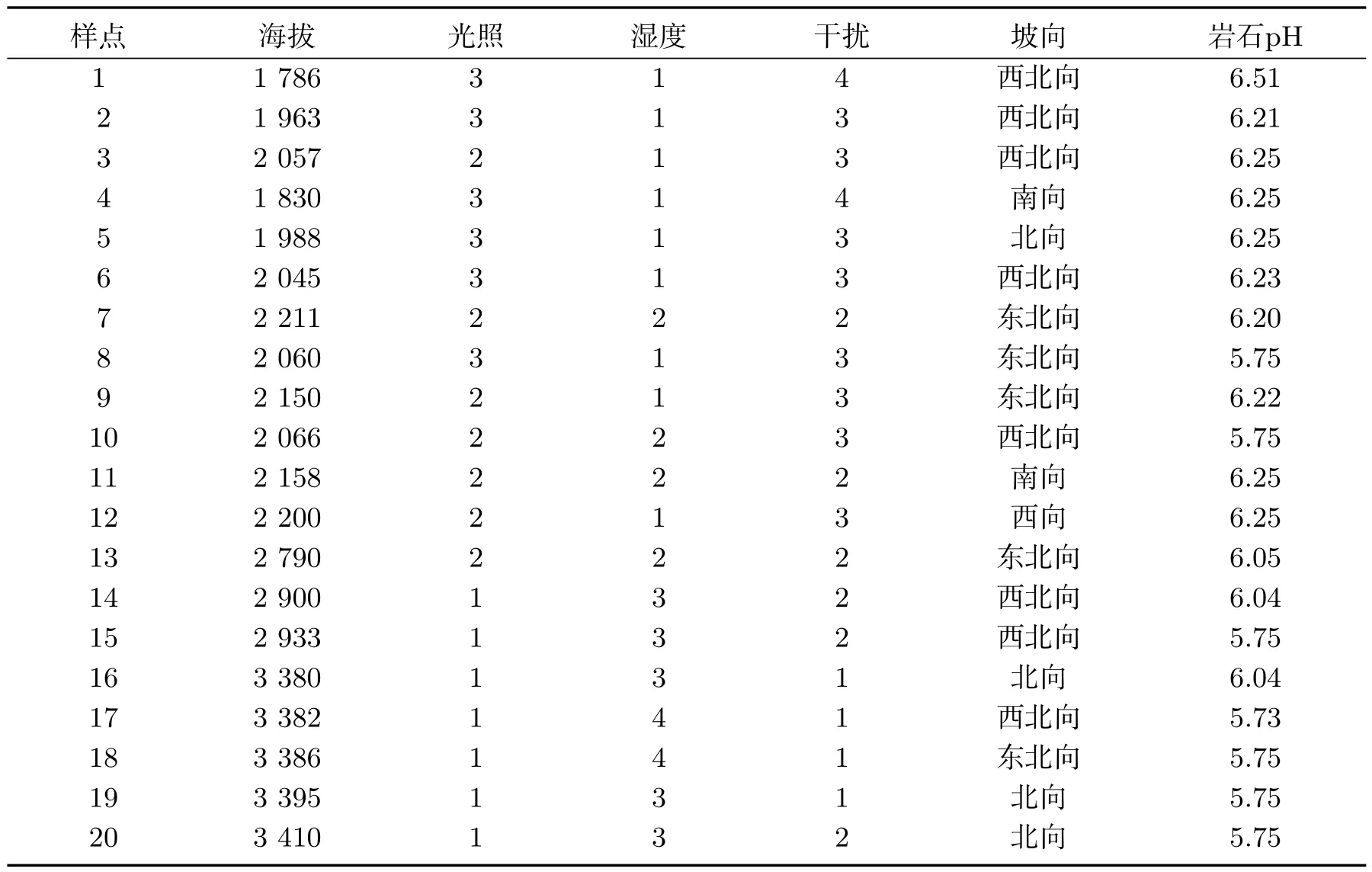
表1 20个样点6种环境因子Tab 1 Six environmental factors in 20 sites
采集的地衣标本在实验室采用形态解剖观察方法结合化学显色反应法进行物种鉴定[36].用舜宇SZM45体视显微镜(上海光学仪器一厂)观察地衣体的皮层、髓层和子囊盘部分.将地衣标本制作成切片,用Nikon ECLIPSE E100光学显微镜观察子囊和子囊抱子等形态特征.
1.3 数据分析
以岩面生地衣盖度为指标,样点为对象用PCORD 5软件进行聚类分析(CA)和除趋势对应分析(DCA),根据多元分析结果对岩面生地衣群落进行了数值分类,结合优势种特征对地衣群落进行命名[37,38].用CANOCO 4.5 for Windows软件进行典范对应分析(CCA),分析了环境因素对岩面生地衣种类分布的影响.CCA分析时由除趋势对应分析(DCA)得出地衣种的单峰响应值(梯度长度SD),当SD>2 时即可进行CCA分析.并用层次分割法校正单环境因子对物种组成的贡献率,解释的显著性用蒙特卡罗置换检验(Monte Carlo permutation tests)[37,38].根据各样点岩面生地衣的盖度,计算各群落的物种丰富度、多样性和相似性指数.
Patrick丰富度指数Dp=s,式中s为群落内物种数.Margalef丰富度指数D=(S−1)/lnN,Menhinnick丰富度指数D=lnS/lnN,式中S为物种数;N为全部种的盖度总数.Shannon-Wiener多样性指数[37,38]式中H为群落的多样性指数;Pi为第i个物种相对盖度的百分比;s为总种数.均匀度指数E=H/Hmax,H为实测多样性值.Hmax为理论上群落多样性的最大值,Hmax=lns,其中s为总种数.Simpson多样性指数[37,38]D=,式中s为物种数量;pi为第i物种相对盖度的百分比.Whittaker相似性指数[37,38]式中I 为相似性指数;ai为第i个物种的盖度在群落a中的比例;bi为第i个物种的盖度在群落b中的比例;S为a,b群落中相应的种数.Jaccard’s相似性指数式中a为两个群落共有的种数;b和c分别为仅存在于不同两个群落的种数.
2 结果与分析
2.1 岩面生地衣群落种类组成
本研究共鉴定分布在新疆博格达峰南坡的岩面生地衣68个种,隶属于10目17科26属(表2).在科属种类组成上茶渍目地衣占优势,有4科8属17种,分别占该地区地衣科数种类总数量的23.53%、30.76%和20.58%;其次为鸡皮衣目共有1科3属14个种;再次为微孢衣目有1科2属11个种.瓶口衣目、无座盘菌目、地图衣目、地卷目、石耳目和黄茶渍目等6个目的种数都少于5个种,说明博格达峰南坡的干旱环境有利于壳状地衣种类的分布,限制枝状和叶状地衣种类的分布(表3).

表2 博格达峰南坡岩面生地衣群落的种类组成及盖度Tab 2 Species composition and coverage of saxicolous lichen communities on the southern slope of Bogda Peak

续表2
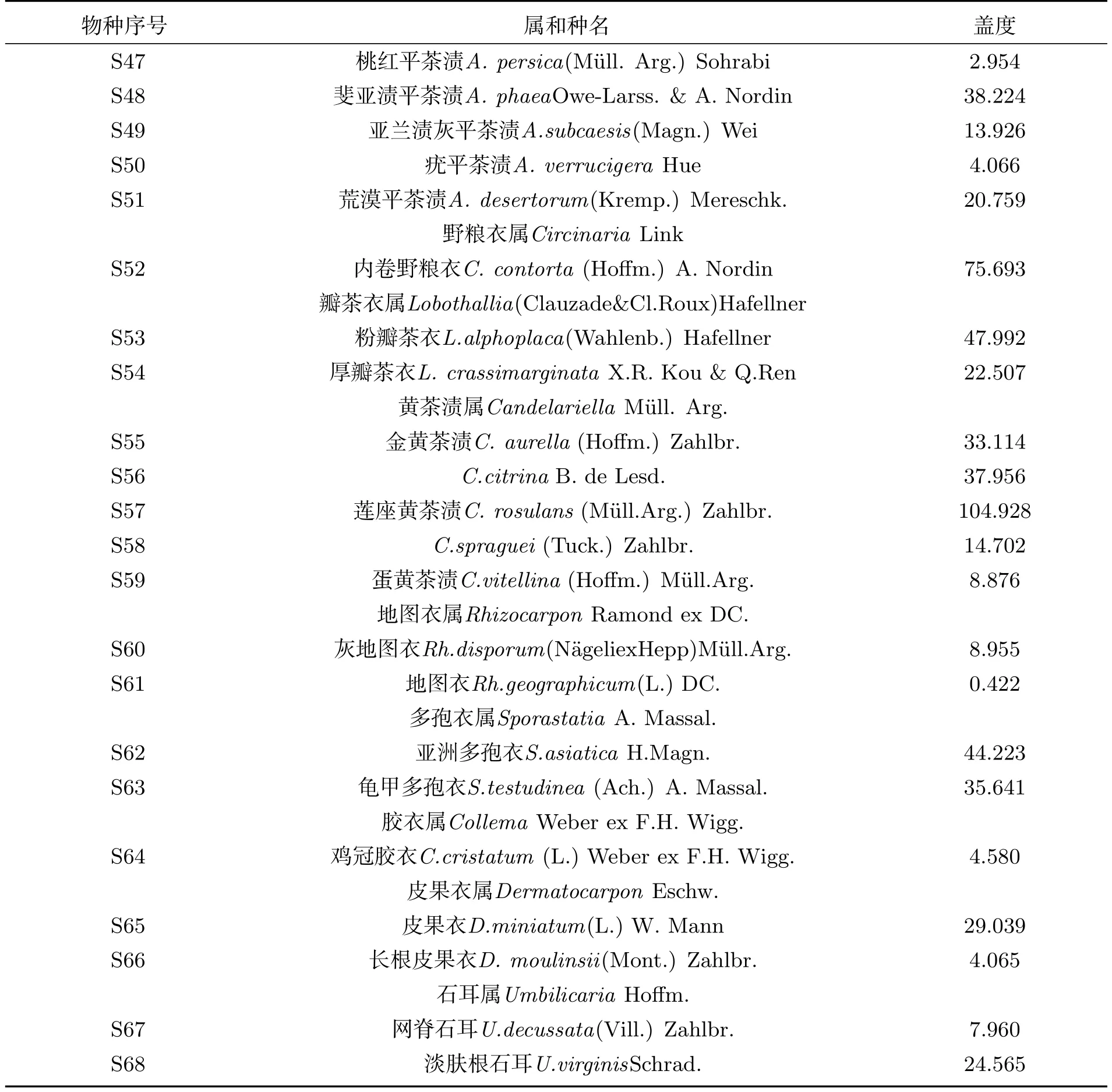
续表2

表3 博格达峰南坡岩面生地衣数量统计Tab 3 Statistics of saxicolous lichen on the southern slope of Bogda Peak
2.2 岩面生地衣群落分类及群落参数
聚类分析和除趋势对应分析的结果见图1和图2.因样点17中的地衣种类较少,覆盖度较低,所以本研究中只分析了19个样点的数据.分析结果表明,19个样点可分为以下4个样点组,我们根据各样点组的优势地衣种类的盖度,采用优势种命名法对岩面生地衣群落进行命名[28,31].

图1 19个样点的聚类分析树状图Fig 1 Cluster analysis dendrogram of 19 sampling sites
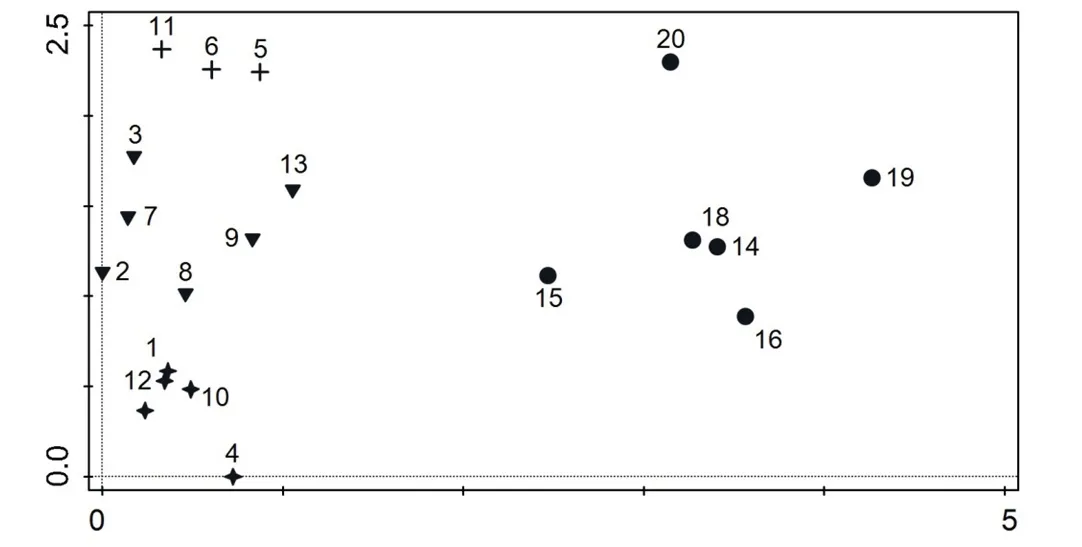
图2 19个样点的除趋势对应分析排序图Fig 2 DCA ordination diagram of 19 sampling sites
样点组1:包括样点1、4、10和12.分布在该样点组的地衣种类有丽石黄衣Xanthoria elegans (Link) Th.Fr.、碎茶渍Lecanora argopholis (Ach.) Ach.、淡腹黄梅Xanthoparmelia mexicana (Gyeln.) Hale、朝鲜黄梅X.coreana(Gyeln.) Hale、杯状平茶渍Aspicilia cupulifera (H.Magn.) Oxner、蛋黄茶渍Candelariella vitellina (Hoffm.)M¨ull.Arg.等20种地衣.地衣平均覆盖度为63.14%,其中覆盖度大于1%的地衣种类有12种,被命名为丽石黄衣+碎茶渍群丛.
样点组2:包括样点5、6和11.主要的岩面生地衣有茸刺山褐梅Momtanelia tominii (Oxner) Divakar et al.、粉芽石黄衣Xanthoria sorediata (Vain.) Poelt、丽石黄衣X.elegans(Link) Th.Fr.、碎茶渍Lecanora argopholis(Ach.) Ach.、刺平茶渍Aspicilia confusa Owe-Larss.&A.Nordin、网脊石耳Umbilicaria decussata (Vill.) Zahlbr.、包氏微孢衣Acarospora bohlinii H.Magn.、盾脐鳞Rhizoplaca peltata (Ramond) Leuckert &Poelt等21个种.地衣平均覆盖度为77.72%,其中覆盖度大于1%的地衣种类有16种,被命名为茸刺山褐梅+粉芽石黄衣群丛.
样点组3:包括样点2、3、7、8、9和13.分布在这些样点的岩面生地衣种类有莲座黄茶渍Candelariella rosulans(M¨ull.Arg.) Zahlbr.、厚瓣茶衣Lobothallia crassimarginata X.R.Kou&Q.Ren、粉瓣茶衣L.alphoplaca (Wahlenb.)Hafellner、拟菊叶黄梅Xanthoparmelia taractica (Kremp.) Hale、汇合网衣Lecidea confluens (Weber) Ach.、杯状平茶渍Aspicilia cupulifera (H.Magn.) Oxner、青蓝色平茶渍A.cyanescensOwe-Larss.&A.Nordin等34个种.地衣平均覆盖度为88.39%,其中覆盖度大于1%的地衣种类有23种,被命名为莲座黄茶渍+厚瓣茶衣群落.
样点组4:包括样点14、15、16、18、19和20.分布于这些样点的岩面生地衣种类有科罗拉多黄梅Xanthoparmelia coloradoensis (Gyeln.) Hale、韦伯黄梅X.weberi (Hale) Hale、金黄茶渍Candelariella aurella(Hoffm.) Zahlbr.、斐亚渍平茶渍Aspicilia phaea Owe-Larss.&A.Nordin、亚兰渍灰平茶渍A.subcaesis (Magn.) Wei、方斑网衣Lecidea tessellata Fl¨orke、破小网衣Lecidellacarpathica K¨orb.等41个种.地衣平均覆盖度为66.56%,其中覆盖度大于1%的地衣种类有18种.被命名为科罗拉多黄梅+韦伯黄梅群落.
为了进一步找出各群丛间的差异,对岩面生地衣群丛进行Shannon-Wiener和Simpson多样性指数、Patrick丰富度指数、Margalef丰富度指数和Menhinnick丰富度指数、Pielou均匀度指数的分析(表4).
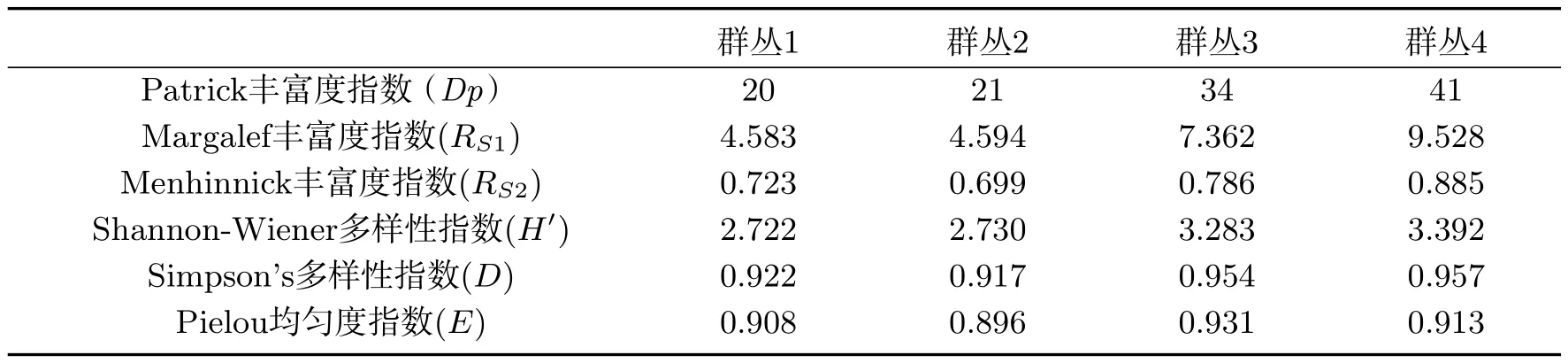
表4 岩面生地衣群落物种丰富度、多样性及均匀度指数Tab 4 Species richness,diversity and evenness index of saxicolous lichen community
由表4可知,群丛4的物种丰富度指数和物种多样性指数最高.组成该群丛的6个样点均分布在海拔2 400 m~3 400 m间的亚高山和高山草甸带,环境比较潮湿,降水量较丰富,光照较弱,几乎不受人为干扰的影响,因此岩面生地衣的物种种数最多分布广泛.组成群丛3的样点分布在海拔1 800~2 400 m之间山地草原带,这里的光照强烈,较干燥,有少量放牧等人为干扰,所以地衣物种多样性比高海拔地区的群丛4小.群丛1和群丛2的样点分布在中低海拔地带,人为干扰强度较大、空气干燥,因此地衣物种多样性较低,种类数量较少.
用Whittaker相似性指数和Jaccard相似性指数比较各群丛的相似性程度(表5).结果显示,Whittaker相似性指数的变化范围在0.272~0.536之间;Jaccard相似性指数的变化范围在0.114~0.211之间.群丛1和群丛2的相似性最高分别为I=0.536,Sj=0.211;其次为群丛3和群丛4的相似性分别为I=0.455,Sj=0.193.群丛2和群丛4的相似性最小为I=0.272,Sj=0.114.进一步说明各群丛的相似性受到群丛间共有地衣种类的数量和群丛分布区环境因素的影响.
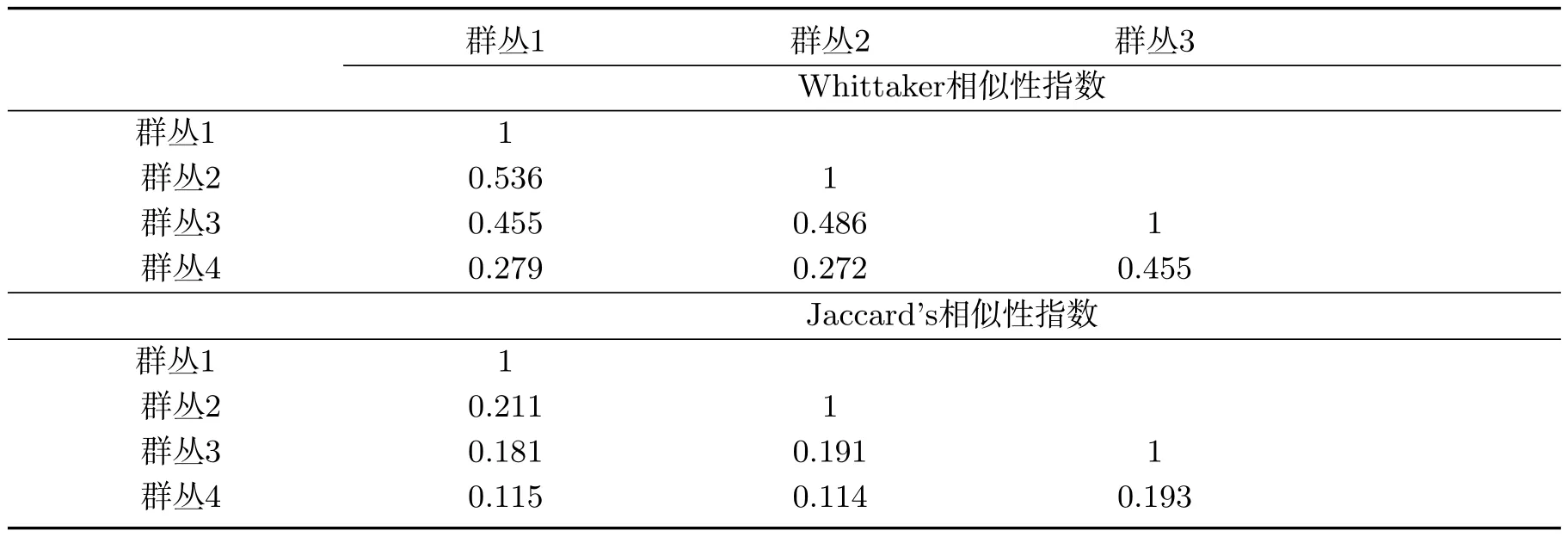
表5 岩面生地衣群落相似性Tab 5 Similarity of saxicolous lichen community
2.3 群落物种分布与环境因子的相关性分析
CCA分析时,首先用除趋势对应分析得出岩面生地衣种分布的单峰响应值(梯度长度SD值).结果表明,其梯度长度SD值均大于2(DCA1=5.261、DCA2=3.098、DCA3=2.567、DCA4=2.716),物种分布变化较大,符合进行CCA排序.对19个样点68种岩面生地衣与6个环境因子间的CCA排序结果显示,前四个轴的特征值分别为0.585 1、0.501 9、0.437 3、0.394 6,前四个轴对于物种分布解释率有47.76%,解释了一部分的物种分布情况;加入环境样方矩阵后,前四个轴的特征值分别为0.698 9、0.298 3、0.315 8、0.295 8,方差累计百分比为89.95%,更好地拟合了物种分布与环境之间的关系,说明典范对应分析结果可靠.
对6个环境因子(海拔、光照、湿度、干扰、坡向、岩石pH)与第一排序轴和第二排序轴的关系进行了蒙特卡罗置换检验,结果表明,海拔和湿度与第一排序轴极显著正相关,因此可以定义第一排序轴为海拔与湿度;而第二排序轴与光照强度和pH正相关,但未达到显著水平(表6).
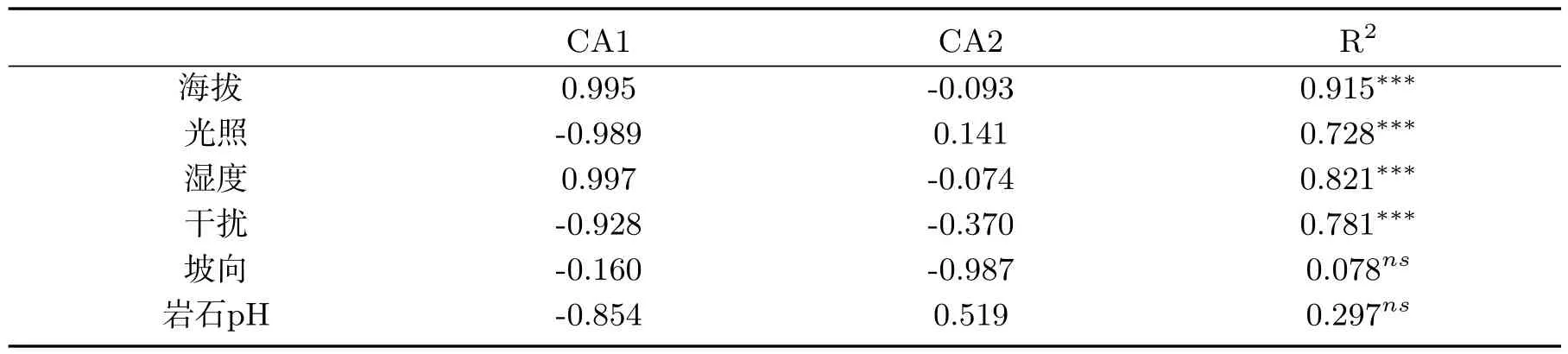
表6 排序轴环境因子的相关性置换检验结果Tab 6 Permutation test results of correlation between sorting axis and environmental factors
在约束排序分析中,虽然获得了各环境因子的贡献率,但由于解释变量间的相关性较显著(表7),置换检验后导致影响显著的环境因子占优势,因此排序结果不是该环境因子真正的解释量.基于层次分割理论[39]计算获得的解释变量真实贡献率的结果显示,坡向对岩面生地衣的影响最大,其贡献率为6.9%,干扰的影响最小,其贡献率为4.1%,6个环境因子的总解释率为33.4%,均未达到显著水平(表8).

表7 环境因子的相关性分析Tab 7 Correlation analysis of environmental factors

表8 对层次分割结果的置换检验Tab 8 Permutation test on Hierarchical Segmentation Results
CCA排序图3和图4综合反映了19个样点和68种岩面生地衣种分布与6个不同环境因子间的关系.

图3 样点分布与环境因子关系的CCA排序图Fig 3 CCA revealing the relationships between sampling sites and environmental factors

图4 岩面生地衣分布与环境因子关系的CCA排序图Fig 4 CCA revealing the relationships of the saxicolous lichens species with environmental factors
从CCA排序图3和图4得知,第一象限包括分布在样点13、18和20的16个地衣种.其中拟菊叶黄梅Xanthoparmelia taractica (Kremp.) Hale、齿裂黄梅X.protomatrae(Gyeln.) Hale、菊叶黄梅X.stenophylla(Ach.) Ahti &D.Hawksw.、亚兰渍灰平茶渍Aspicilia subcaesis (Magn.) Wei、亚洲多孢衣Sporastatia asiatica H.Magn.等种类的分布及盖度出现随着海拔变化的现象,说明海拔是影响这些种类分布的因素.内卷野粮衣Circinaria contorta(Hoffm.) A.Nordin出现在干扰较小的高海拔地带,它的分布受到坡向的影响,岩石不同坡向该地衣的盖度和频度具有差异.第二象限包括分布在中低海拔地带的样点7至样点9.其中爱琴海微孢衣Acarospora aeginaica H.Magn.、间隔微孢衣A.interrupta (Ehrenb.ex Nyl.) Vain.、软骨鳞茶渍Squamarina cartilaginea (With.) P.James、杯状平茶渍Aspicilia cupulifera (H.Magn.) Oxner和龟甲多孢衣Sporastatia testudinea (Ach.) A.Massal.等种类分布在干扰较小的岩石北面.被膜微孢衣Acarospora molybdina Trevis.、斯塔妃微孢衣A.stapfiana(M¨ull.Arg.) Hue、汇合网衣Lecidea confluens (Weber) Ach.和长根皮果衣Dermatocarpon moulinsii (Mont.)Zahlbr.分布在较干燥的岩石南面.第三象限包含的样点1、2、4、6和12的地衣种类的分布与岩石的酸碱度有关.样点3、5和10的地衣种类分布在光照较弱,干扰小的地带,对岩石酸碱度的要求不高.例如珠光网衣Lecidea perlatolica Hertel &Leuckert、杜金平茶渍Aspicilia dudinensis (H.Magn.) Oxner、桃红平茶渍A.persica(M¨ull.Arg.) Sohrabi和Candelariella citrina B.de Lesd.的种类的分布与岩石酸碱度有着高度的相关性.Anamylopsora sp、Xanthomendoza trachxphylla (Tuck.) Fr¨od´en、灰地图衣Rhizocarpon disporum (N¨ageli ex Hepp) M¨ull.Arg.等种类中低海拔光照强度较高的地带.第四象限包括高海拔地带的样点14、15、16和19.分布在该样点的朝鲜黄梅Xanthoparmelia coreana (Gyeln.) Hale、韦伯黄梅X.weberi (Hale) Hale、散茶渍Lecanora dispersa (Pers.) R¨ohl.和粉芽石黄衣Xonthoria sorediata (Vain.) Poelt等种类分布在较潮湿的高海拔地带.美国平茶渍Aspicilia americana B.de Lesd.、杯状平茶渍A.cupulifera(H.Magn.) Oxner、荒漠平茶渍A.desertorum(Kremp.) Mereschk.、蛋黄茶渍Candelariella vitellina(Hoffm.) M¨ull.Arg.、糙聚盘衣Glypholecia scabra (Pers.)M¨ull.Arg.、破小网衣Lecidella carpathica K¨orb.和盾脐鳞Rhizoplaca peltata (Ramond)Leuckert&Poelt等种类广泛的分布在不同海拔地带,对环境的适应性较强.
3 讨论
研究群落中各不同种类的分布与环境因子之间关系,找出影响物种分布格局和群落结构特征的因素一直以来是植物群落生态学中的热点研究领域,也是地衣生态学家研究地衣群落构建机制的核心[35].对于岩面生地衣群落而言,影响其物种分布的不仅是地衣种类生长和分布的岩石的理化性质(元素种类、pH等),而且岩石坡度、方向、温度、湿度、海拔、干扰等因素对岩面生地衣种类的分布以及地衣群落构建等方面起到决定作用[40,41].例如Foder在罗马尼亚东南部的Macin山国家公园研究以花岗岩为主的酸性岩石上岩面生地衣群落的丰富度、群落结构以及群落构建机制.他发现研究区的岩面生地衣是由不同的局部地衣群落组成的集合群落(metacommunity),各局部群落的种类组成随着岩石的性质、光照强度、其它地衣种类或植物间的竞争、湿度和富营养化等各种人为干扰而变化[42].
本研究在博格达峰南坡选择不同海拔地带的20个样点,对岩面生地衣群落的物种组成和群落结构进行了生态学定量研究.研究结果显示,在博格达峰南坡随着海拔和坡向等因素的不同各样点的湿度和光照强度发生相应的变化,这种变化最终导致了不同海拔岩面生地衣群落种类组成和分布间的差异.CCA分析结果表明,博格达峰南坡岩面生地衣群落地衣种类的分布主要受海拔和坡向的影响,其余光照强度、干扰和岩石理化性质的影响不大.所调查的68种岩面生地衣中,平茶渍属(Aspicilia A.Massal.)地衣在中海拔至高海拔的丰富度较高,同时也对各种干扰有一定的耐受能力;茶渍属(Lecanora Ach.)、小网衣属(Lecidella K¨orb.)、脐鳞属(Rhizoplaca Zopf)地衣适应中海拔,无干扰的样点;黄茶渍属(Candelariella M¨ull.Arg.)、皮果衣属(Dermatocarpon Eschw)地衣对干扰的抵抗力最佳.稀疏化分析的结果表明,设置在高海拔的样点18、样点19和样点20中岩面生地衣群落物种多样性最高.在海拔2 700 m以上的样点岩面生地衣群落的物种多样性比低海拔样点的岩面生地衣群落的物种多样性高,我们认为这是由于研究区高海拔样点的降水量较充足、栖息环境较稳定等因素有关.高海拔地区各样点受到的干扰类型和强度较小,因为干扰强度的升高会直接限制岩面生地衣群落物种多样性和种类的组成,导致不同海拔岩面生地衣群落结构间的差异,这与Shukla等的研究结果一致[15].
Monte研究环境因素对玄武岩、钙质岩、花岗岩、粗面岩、凝灰岩等不同类型岩石表面岩面生地衣的繁殖和分布时发现,地衣体内的共生藻为橘色藻属的种类分布在岩石北面的光照强度较低,较潮湿的地方;而在比较干燥的环境下带有蓝藻的地衣占优势,于此同时微化境对地衣繁殖策略产生选择性压力[43].本研究结果显示,在博格达峰南坡岩面生地衣种类组成及群落物种分布格局的差异不仅受环境因素的影响,还与不同地衣种类的适应能力和功能性状有关.例如.嗜氮的丽石黄衣Xanthoria elegans (Link) Th.Fr.与嗜氮和嗜酸的地图衣Rhizocarpon geographicum (L.) DC.等种类广泛的分布在不同海拔的岩石表面,进一步体现了壳状地衣对干旱环境的较高的适应能力.另外,地衣体内有蓝藻的鸡冠胶衣Collema cristatum (L.) Weber ex F.H.Wigg.、坚韧胶衣C tenax (Sw.) Ach、地卷Peltigera rufescens (Weiss) Humb.种类分布在比较潮湿的环境,地生同层枝衣Peccania terricola H.Magn.、黑小极衣Lichinella nigritella (Lettau)P.P.Moreno&Egea、黑鳞壁衣Psorotichia nigra H.Magn.分布光照较强的中等潮湿的环境,皮果衣Dermatocarpon miniatum (L.) W.Mann、长根皮果衣Dmoulinsii(Mont.) Zahlbr.、盾鳞衣Placidium squamulosum (Ach.) Breuss和粉芽盾衣Peltula euploca (Ach.)Poelt ex Ozenda &Clauzade等分布在干旱的岩石表面,我们在博格达峰北坡研究时也发现同样的现象[31].其余地衣体内有共生绿球藻或球形绿藻的种类广泛的分布在不同海拔较好的适应栖息地和环境.在岩面生地衣的繁殖策略方面也形成了对干旱环境的进一步适应,例如具有粉芽的种类分布在岩石的裂缝、侧下面来保护粉芽不受雨水和大风的破坏,具有裂芽和子囊盘的种类分布在岩石表面和侧面的不同部位,这个结果与博格达峰北坡岩面生地衣群落的分布相似[31].综上所述,新疆博格达峰南坡的环境比北坡干旱,岩面生地衣种类主要以适应干旱环境的壳状地衣为主.通过研究发现,海拔、干扰强度、光照强度、湿度、岩石pH等因素直接和间接的影响岩面生地衣种类的分布,其中海拔和坡度的影响最大.与此同时不同岩面生地衣种类的功能形状、对环境的适应能力之间的差异也影响岩生地衣群落结构和群落构建.
猜你喜欢
中国水土保持科学(2023年6期)2024-01-05 05:53:12
水土保持研究(2022年1期)2022-12-27 07:35:24
湖北植保(2022年4期)2022-08-23 10:51:52
土壤(2021年1期)2021-03-23 07:29:06
收藏界(2018年1期)2018-10-10 05:23:08
创新作文(小学版)(2018年31期)2018-05-16 03:00:30
土壤学报(2017年5期)2017-11-01 09:21:27
摄影之友(影像视觉)(2017年1期)2017-07-18 11:12:16
小学阅读指南·低年级版(2016年6期)2016-05-14 21:39:20
山西大学学报(自然科学版)(2014年2期)2014-05-10 06:55:16
