
芥子酸改善高脂饮食模式下叙利亚仓鼠脂代谢与氧化应激水平
2021-10-09 05:19梁婵华王可盈曹文瀞罗高何雯曾榛宋家乐
现代食品科技 2021年9期
梁婵华,王可盈,曹文瀞,罗高,何雯,曾榛,3*,宋家乐,4*
(1.桂林医学院公共卫生学院,广西桂林 541101)(2.桂林医学院学生工作(部)处,广西桂林 541101)(3.中南大学湘雅公共卫生学院,湖南长沙 410000)(4.桂林医学院第二附属医院临床营养科,广西桂林 541109)
肥胖是由于机体长期处于能量摄入与消耗失衡状态而引起的一种慢性代谢性疾病[1],主要以体内脂肪积聚过多所致脂肪层过厚和脂肪积蓄过剩为特征[2]。随着我国经济发展和居民生活水平的不断提高,西方饮食文化(高脂肪、高热量饮食模式)对中国传统饮食文化冲击所造成的饮食行为改变和结构不均衡,以及不良生活习惯、环境和遗传等因素[3],使得肥胖成为中国乃至全球日益瞩目的公共健康问题之一。高脂饮食(High fat diet,HFD)诱导的肥胖不仅会引起机体氧化应激、炎症和肠道菌群失调[4],从而导致代谢综合征的发生,增加心血管疾病、胰岛素抵抗、糖尿病、非酒精性脂肪肝病、癌症,甚至哮喘的发病风险[5-7],而且还影响着社会与经济发展。目前,全球的成人肥胖发生率约为13%[8]。此外,儿童超重与肥胖的发生率也迅速增加,2016 年全球5~19 岁超重或肥胖的儿童已超过3.4 亿人[9]。因此,寻找有效预防和治疗肥胖的方法迫在眉睫。
芥子酸(Sinapic acid,SA)是广泛分布于水果、蔬菜、油料作物和谷物中的一类具有较强生物活性的天然酚酸类化合物,属于羟基肉桂酸的一种[10]。研究发现,其在菜籽、紫山药、柠檬、柑橘类等蔬果植物中含量很高[11,12]。近年来的研究结果也提示,芥子酸具有抗结肠炎[13,14]、抗氧化[15]、抗癌[16]、促进脂肪分解[17]以及调控游离脂肪酸、磷脂、三酰甘油和氧磷脂等脂类间的转化代谢[14]等生理作用。
叙利亚黄金仓鼠具有与人类相似的脂类代谢通路,是目前较为公认的用于代谢综合征研究的动物模型之一[18]。目前,芥子酸对高脂肪膳食模式下仓鼠脂类代谢影响及其作用机制的报道较少。因此,本研究以膳食高脂肪饲料(60%脂肪供能)诱导的叙利亚黄金仓鼠建立肥胖模型,同时观察实验饲料(高脂饲料中添加0.03%芥子酸)对高脂膳食模式下仓鼠体重改善、脂质代谢及氧化应激发生的影响效果,从而探讨芥子酸抗肥胖的作用机制,为进一步将芥子酸开发成为更安全、更有效的抗肥胖膳食补充剂提供一定的科学理论依据。
1 材料与方法
1.1 材料与试剂
芥子酸(纯度大于98%,S106903),购自上海阿拉丁生物科技有限公司;AIN-93G 普通饲料、对照组高脂饲料(60%脂肪供能)、实验组高脂饲料(含60%脂肪,0.03%芥子酸),购于常州鼠一鼠二生物科技有限公司;总胆固醇(Total cholesterol,TC)、甘油三酯(Triglyceride,TG)、低密度脂蛋白胆固醇(Low-density lipoprotein cholesterol,LDL-C)、高密度脂蛋白胆固醇(High-density lipoprotein cholesterol,HDL-C)、游离脂肪酸(Nonesterified free fatty acid,NEFA)、丙二醛(Malonic dialdehyde,MDA)、总抗氧化能力(Total antioxidation capability,T-AOC)试剂盒,南京建成生物技术有限公司;BCA 蛋白浓度试剂盒,肉毒碱棕榈酰转移酶1(CPT-1)、过氧化物酶体增殖物激活受体-γ(PPAR-γ)、胆固醇7α-羟化酶(CYP7A1)、羟甲基戊二酸单酰辅酶A 还原酶(HMGCR)、乙酰辅酶A 羧化酶1(ACC1)抗体,上海碧云天生物技术有限公司;脂肪酸合成酶(FAS),武汉塞维尔生物科技有限公司;胆固醇合成转录因子SREBP2,北京索莱宝科技有限公司。
1.2 仪器与设备
FA2004 电子天平,上海舜宇恒平科学仪器有限公司;Eppendorf 5424R 型冷冻离心机,德国Eppendorf公司;ELx808 酶标仪,美国BioTek 公司;FluorChem M 超灵敏全自动化学发光成像分析系统,美国Protein Simple 公司;KZ-II 高速组织研磨仪,武汉塞维尔生物科技有限公司。
1.3 实验动物
雄性叙利亚黄金仓鼠(共21 只,体重110~140 g)购自北京维通利华实验动物技术有限公司,饲养于桂林医学院公共卫生学院SPF 级动物房(许可证号:SYXK(桂)2020-0005)。饲养环境为标准光照/黑暗(12 h/12 h)循环饲养,自由摄食及清洁饮水。
1.4 实验方法
1.4.1 高脂模型建立及实验分组方法
所有仓鼠经适应性饲养1 周后,依照完全随机数字表法分为3 组(7 只/组)进行后续研究。仓鼠高脂肥胖模型以Dan 等[19]的文献进行操作。其中,正常组饲喂标准饲料、模型组饲喂高脂饲料、芥子酸组饲喂实验高脂饲料(含0.03%芥子酸)。总实验周期为12 周,每3 d 进行体质量监测,每日进行饮水量和摄食量的监测。本动物实验方案已通过桂林医学院动物伦理委员会审查批准(GLMC-201806003)。
1.4.2 组织取材
实验第12 周干预结束后,各组仓鼠禁食不禁水12 h,予腹腔注射1%戊巴比妥钠(0.1 mL/10 g)麻醉,腹主动脉取血,常温下分离血清后存放与-80℃冰箱。仓鼠置于冰上开腹后,快速分离仓鼠附睾、肾周、腹股沟等出脂肪组织及肝脏组织并迅速放入-80℃冰箱保存备用。
或疑月中有兔形,……予以为月无光,而溯日为明,世所知也。天有十二辰,列于东方者。有神司其位。日出在东,其对在酉,酉为鸡,日光含景,则鸡在日中。及运而西。则对在卯,卯为兔,月光含景,则兔在月中。月有兔形,何足异哉?人知日中为乌,而不知为鸡。知月中有兔,不知兔自日以传形也。或曰段成式言月中有桂,仙人吴刚斫其根。曰:不然,日行于西,与扶桑对,则陊景日中,月望之明,景亦随之。[注](明)周婴:《巵林》,新文丰出版社,1984年,第98-99页。
1.4.3 血清生化指标检测
按照各试剂盒检测操作要求,分别检测血清中TG、TC、LDL-c、HDL-c、NEFA 的浓度。
1.4.4 肝脏内TC 与TG 水平分析
称取一定质量的肝脏组织,冰生理盐水去除血污后,加入2 mL 的无水乙醇与丙酮(V/V,1:1)混合液,冰上制备10%的肝组织匀浆。经3000 r/min 离心15 min 后,取上清液用试剂盒检测TC、TG 的浓度。
1.4.5 血清和肝脏组织中T-AOC和MDA水平检测
取冻存的血清和肝脏组织匀浆液,严格按照试剂盒说明操作,分别检测血清中及肝脏中MDA 和T-AOC 的含量水平。
1.4.6 粪便中TC 的检测
1.4.7 Western Bolt 法检测肝脏组织中相关蛋白的表达水平
称取一定质量肝脏组织(20~35 mg),冰生理盐水清洗血污,按比例加入蛋白裂解液(含PMSF)提取组织总蛋白,12000 r/min,4℃条件下离心10 min,收集上清采用BCA 法测定蛋白浓度。配制SDS-聚丙烯酰胺凝胶电泳(SDS-PAGE)胶,加样后调整电压,浓缩胶80 V,分离胶120 V,电泳120 min。湿法转膜,条件为恒定电流240 mA。转膜1 h 后,将NC 膜放于5%的脱脂奶粉中封闭4 h。孵育一抗过夜,TBST 洗膜后,孵育二抗1 h,TBST 洗膜3 次后,加入特超敏ECL化学发光试剂,自动化学发光成像仪检测相关蛋白的表达情况。
1.5 数据处理与统计分析
所有实验均重复3 次,结果以均值(means)±标准偏差(SD)表示。所得实验数据运用SPSS 24.0 统计软件进行单因素方差分析与统计处理,p<0.05 为具有统计差异。
2 结果与讨论
2.1 芥子酸对高脂饮食诱导的仓鼠体质量和器官质量的影响
由表1 可知,正常组、高脂肪组及芥子酸组的初始体重均无显著性差异(p>0.05)。喂养12 周后,与正常组相比,高脂肪组仓鼠最终体重增加1.77 倍(p<0.05)。芥子酸处理组仓鼠最终体重和体重增加量均显著低于高脂肪组(p<0.05),分别减少了11.15%和23.86%。结果表明,芥子酸有显著下调仓鼠体重的作用。但Chen 等[3]的研究结果也提示,给予芥子酸干预并不能够改善高脂饲料(45%脂肪供能)喂养后大鼠体重的增加。
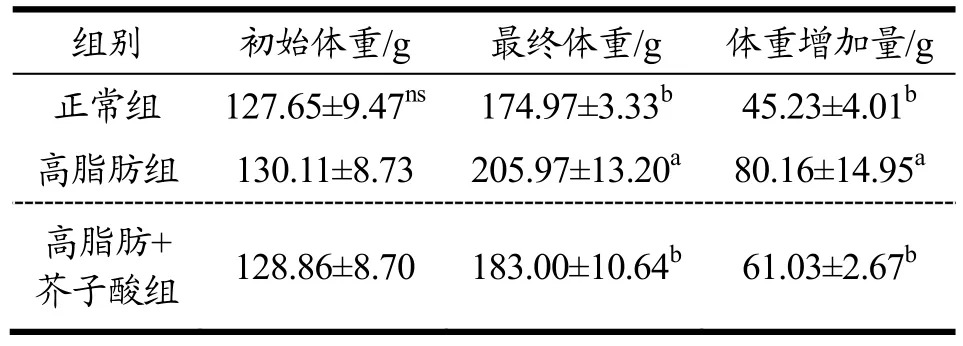
表1 芥子酸对饲喂高脂饮食后仓鼠体重的影响 Table 1 Effects of sinapic aicdon body weight in high fat diet fed hamsters
如图1 所示,相比正常组,高脂肪组仓鼠的肝脏、附睾脂肪和腹股沟脂肪组织的重量极显著增加(p<0.01)。而饲喂高脂肪饲料虽较正常组能够促进仓鼠肾周脂肪的堆积(即肾周脂肪脏器指数增加趋势),但两者相比无统计学意义(p>0.05)。与高脂肪组相比,芥子酸干预对仓鼠肝脏、附睾脂肪、肾周脂肪和腹股沟脂肪的脏器指数分别减少13.05%、20.38%、27.91%和12.24%。这一结果提示,芥子酸可能通过缓解脂肪组织堆积抑制高脂饮食诱导的仓鼠体重的增加。另外,芥子酸组对正常组附睾脂肪和肾周脂肪组织的重量均无显著性影响(p>0.05)。

图1 芥子酸对饲喂高脂饮食后仓鼠肝脏与各脂肪的脏器指数的影响 Fig.1 Effects of sinapic aicdon liver and obesity tissue index in high fat diet fed hamsters
2.2 芥子酸对饲喂高脂饮食后仓鼠血清脂代谢水平的影响
从图2 可知,与正常组相比,高脂肪组仓鼠血清中TG、TC、LDL-c 和NEFA 水平显著增加(p<0.05),而HDL-c 水平降低,但无统计显著差异(p>0.05)。与高脂模型组相比,芥子酸干预能够显著降低高脂肪膳食所引发的仓鼠血清中TC、TG、NEFA 和LDL-c的水平(p<0.05),分别减少了36.54%、26.54%、44.13%和72.40%,而HDL-c 水平则升高了1.25 倍。Chen 等[3]研究提示,芥子酸干预能够有效降低肥胖大鼠血清TG 水平,同时提升HDL-c 水平。同时,Wu 等[13]的研究也提示芥子酸干预能够减少高脂饮食下C57BL/6J 背景的ApoE−/−小鼠血清游离脂肪酸和总TG 水平。而相比正常组,芥子酸干预能够显著降低仓鼠血清LDL-c 的水平至48.74%(p<0.01),HDL-c增加1.17 倍(p<0.01),而对TG 和TC 的水平无明显改变。

图2 芥子酸对饲喂高脂饮食后仓鼠血清脂代谢水平的影响 Fig.2 Effects of sinapic aicdon serum lipid level in high fat diet fed hamsters
2.3 芥子酸对饲喂高脂饮食后仓鼠肝脏脂代谢水平的影响
如图3 所示,高脂膳食能够显著诱发仓鼠肝脏TG和TC 水平升高。与正常组相比,高脂膳食组仓鼠肝脏中TG 和TC 水平分别升高了约1.90 和1.47 倍。而芥子酸干预却能降低肝脏中TG 和TC 的水平。与高脂膳食组相比,芥子酸组的肝脏TC 水平降低了34.58%(p<0.05),TG 水平降低了28.19%,但无显著性差异(p>0.05)。然而,肝脏TC 与TG 水平在正常组和芥子酸组之间没有显著差异。
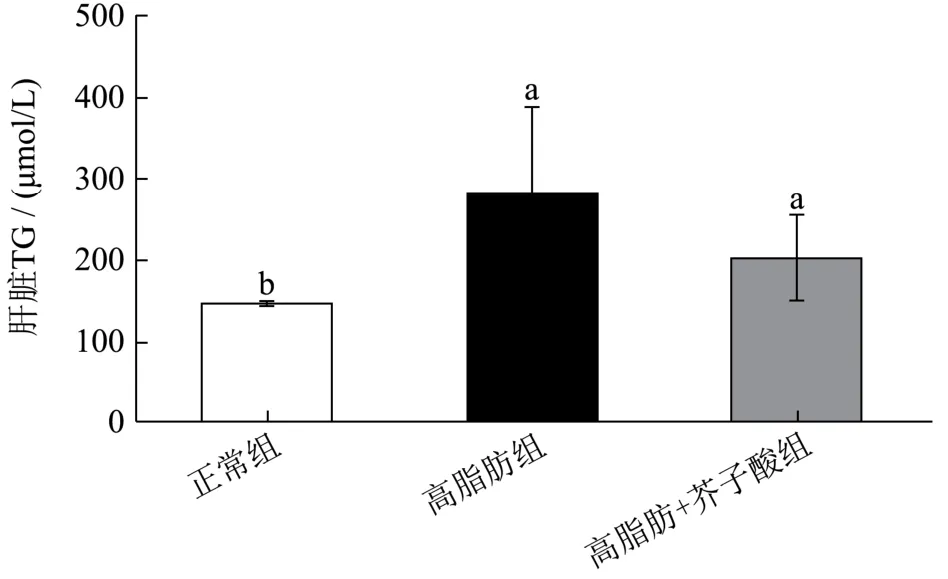
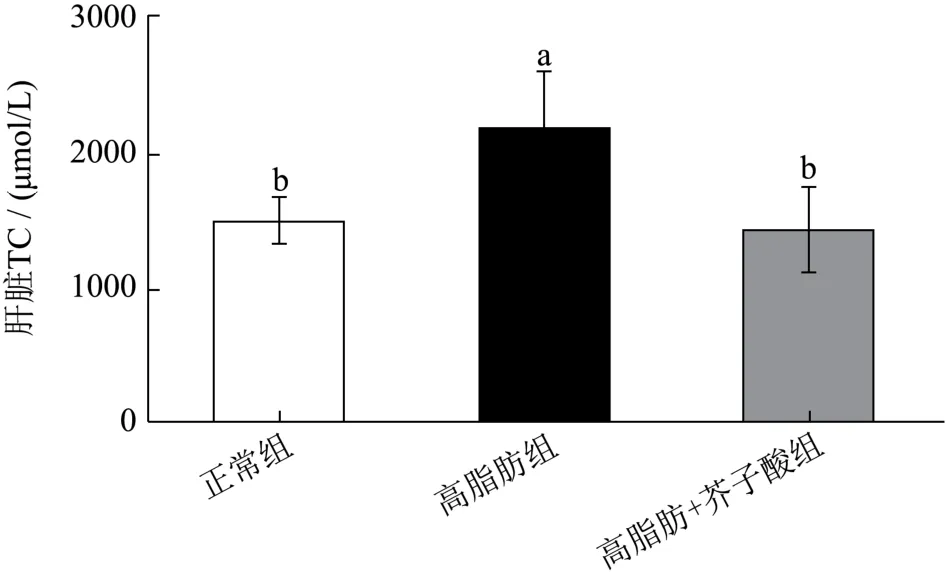
图3 芥子酸对饲喂高脂饮食后仓鼠肝脏中TG、TC 水平的影响 Fig.3 Effects of sinapic aicdon hepatic TG and TC level in high fat diet fed hamsters
2.4 芥子酸对饲喂高脂饮食后仓鼠粪便中TC水平的影响
粪便排出的胆固醇含量与血脂水平有密切关系,当机体脂代谢紊乱时其会大幅度下调[19]。由图4 可知,芥子酸干预组粪便中TC 含量(1650.00 μmol/L)显著高于正常组(1177.50 μmol/L)和高脂膳食组(920.00 μmol/L)(p<0.05),这可能是由于芥子酸干预后,降低机体对胆固醇的吸收,并加速胆固醇的代谢,从而促使胆固醇排出体外,该结果与芥子酸干预后的仓鼠外周血脂水平变化相一致。
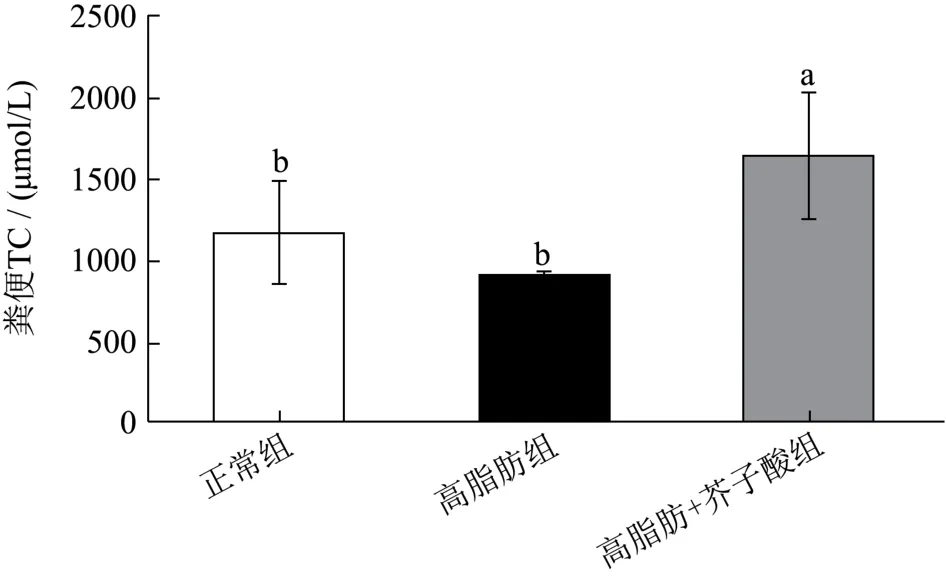
图4 芥子酸对饲喂高脂饮食后仓鼠粪便中TC 水平的影响 Fig.4 Effects of sinapic aicdon fecal TC level in high fat diet fed hamsters
2.5 芥子酸对饲喂高脂饮食后仓鼠血清和肝脏氧化应激水平的影响
在脂肪堆积的同时氧化应激反应的升高是肥胖等相关代谢综合征的重要致病机制[21]。MDA 是生物膜多不饱和脂肪酸过氧化过程中产生的一种主要反应醛类物质,其水平的高低通常可以反映机体内脂质过氧化程度的强弱[22,23]。由表2 可知,12 周的高脂饮食导致血清和肝脏中氧化应激的发展,高脂膳食组仓鼠的MDA 水平显著高于正常组(p<0.05)。芥子酸干预后,血清和肝脏组织中MDA 水平分别降低10.18%和16.23%,但均无显著差异(p>0.05)。芥子酸对血清T-AOC 水平有改善作用,但对肝脏组织内T-AOC 水平无明显影响,这一实验结果与Chen 等[3]的研究结果较为接近。上述结果提示芥子酸可能具有缓解脂质过氧化反应的作用,从而纠正脂代谢紊乱。

表2 芥子酸对饲喂高脂饮食后仓鼠血清和肝脏氧化应激水平的影响 Table 2 Effects of sinapic aicdon serum and liver oxidative stress level in high fat diet fed hamsters
2.5 芥子酸对饲喂高脂饮食后仓鼠肝脏脂肪酸合成的影响
FAS 和ACC1 是合成脂肪酸的关键酶,其蛋白水平的高低可间接反映肝脏组织脂肪的合成情况[24,25]。异常活化的FAS 和ACC1 可破坏脂代谢平衡,使得体内脂肪合成速度快于脂肪分解,从而诱导脂肪的过度堆积,加速肥胖发生的进程[26,27]。高脂饮食喂养能够显著上调仓鼠肝脏组织中FAS和ACC1的蛋白表达水平(图5)。与正常组相比,高脂膳食组仓鼠肝脏中FAS 表达水平上升约6.17 倍,ACC1 表达水平升高2.35 倍。而经芥子酸干预后,相比高脂肪膳食组,仓鼠肝脏中FAS 和ACC1 的蛋白表达水平分别下降34.38%和52.72%(p<0.05)。


图5 芥子酸对饲喂高脂饮食后仓鼠肝脏中FAS 和ACC1 的蛋白表达影响 Fig.5 Effects of sinapic aicd on hepatic protein expression of FAS and ACC1 in high fat diet fed hamsters
2.6 芥子酸对喂高脂饮食后仓鼠肝脏脂肪酸β氧化的影响
PPAR-γ是核受体超家族成员,主要参与肝脏脂类代谢和脂肪细胞的合成、分化过程[28]。PPAR-γ和CPT-1 是控制脂肪酸β氧化的关键蛋白,其中CPT-1是脂肪酸β氧化的限速酶[29,30]。在本研究中,12 周高脂膳食导致仓鼠肝脏组织中PPAR-γ和CPT-1 蛋白水平均显著低于正常组仓鼠,且两种蛋白的表达水平分别较正常组降低了41.60%和31.87%(图6)。与高脂组相比,芥子酸干预能上调仓鼠肝脏组织中PPAR-γ和CPT-1 的蛋白表达水平,分别增加了2.00 倍和2.61倍。而文献报道指出,激活PPAR-γ和CPT-1 能够促进脂肪细胞分化,减少脂肪细胞数量,控制脂质过氧化水平,并利于调节脂质代谢[31,32]。

图6 芥子酸对饲喂高脂饮食后仓鼠肝脏中PPAR-γ 和CPT-1 的蛋白表达影响 Fig.6 Effects of sinapic aicdon hepatic protein expression of PPAR-γ and CPT-1 in high fat diet fed hamsters.
2.7 芥子酸对喂高脂饮食后仓鼠肝脏胆固醇代谢的影响
SREBP2 主要参与机体内胆固醇代谢过程,通过激活胆固醇合成途径中多个因子的表达(如HMGCR)来调节胆固醇的生成、吸收、排泄等生理进程,过度活化的SREBP2 和HMGCR 在代谢综合征病理生理过程中发挥着重要作用[33]。如图7 所示,芥子酸处理显著抑制高脂膳食所造成仓鼠肝脏中SREBP2(53.49%)和HMGCR(32.68%)的表达(p<0.05)。而与正常组相比,芥子酸干预后的肥胖仓鼠肝脏内SREBP2 和HMGCR 蛋白表达的水平降低64.69%和45.46%(p<0.05)。CYP7A1 是胆固醇从体内排出的重要转运蛋白,促进肝脏内胆固醇代谢正常运行[34]。与高脂膳食组仓鼠相比,芥子酸干预对肥胖仓鼠肝脏中CYP7A1 的蛋白水平显著升高1.65 倍(p<0.05)。以上结果提示芥子酸能够抑制胆固醇的合成,并通过促进胆固醇从体内排出达到改善胆固醇代谢异常的作用。

图7 芥子酸对饲喂高脂饮食后仓鼠肝脏中SREBP2、HMGCR 和CYP7A1 的蛋白表达影响 Fig.7 Effects of sinapic aicdon hepatic protein expression of SREBP2,HMGCR and CYP7A1 in high fat diet fed hamsters
3 结论
连续12 周的芥子酸干预能有效抑制高脂饮食所造成的仓鼠体重增加,减少其体内脂肪组织的质量;下调TG、TC、NEFA 和LDL-c 水平,同时升高HDL-c水平和粪便中TC 含量。芥子酸还能同时增强肝脏组织中PPAR-γ、CPT-1 和CYP7A1 的蛋白表达,并通过下调ACC1、FAS、HMGCR 及SREBP2 等脂质代谢相关因子来控制脂肪合成与胆固醇代谢来调控肥胖发生进程中的脂类代谢异常。此外,芥子酸还能增强血清和肝脏组织中T-AOC 水平,降低血清与肝脏组织中MDA 水平,从而减缓脂质氧化应激的发生。综上所述,芥子酸对高脂饮食引起的仓鼠肥胖和脂代谢异常有明显的改善作用,该作用可能与抑制脂质过氧化、减少脂肪合成和促进胆固醇代谢有关。
猜你喜欢
童话世界(2020年32期)2020-12-25
中华养生保健(2020年5期)2020-11-16
小学科学(学生版)(2020年1期)2020-01-19
故事会(2019年1期)2019-01-11
新世纪智能(语文备考)(2018年11期)2018-12-29
小学生必读(中年级版)(2018年5期)2018-08-15
金色少年(奇趣科普)(2017年8期)2017-09-07
宝藏(2017年2期)2017-03-20
中国运动医学杂志(2016年3期)2016-07-10
东方艺术·国画(2015年3期)2015-08-20
