
荒漠结皮层藓类植物死亡对表层土壤水分蒸发和入渗的影响
2021-10-09 01:18:42李继文尹本丰索菲娅周晓兵李永刚张元明
生态学报 2021年16期
李继文,尹本丰,索菲娅,周晓兵,陶 冶,张 静,李永刚,张元明,*
1 新疆大学生命科学与技术学院, 乌鲁木齐 830046 2 荒漠与绿洲生态国家重点实验室, 中国科学院新疆生态与地理研究所, 乌鲁木齐 830011
水是限制荒漠生态系统生产力和植被演替的关键因素,水分的入渗和蒸发作为荒漠地表水文循环的重要环节深刻影响着降水在土壤中的再分配过程[1]。由于水分的限制,荒漠生态系统很难形成大面积连续分布的维管植物,但以蓝藻、地衣、苔藓等隐花植物组成的生物土壤结皮却能发育成荒漠地表的重要覆被类型,其盖度在部分区域可达70%以上[2]。生物土壤结皮除显著影响土壤养分、土壤结构外[3- 4],在地表水分的入渗、蒸发等水文过程中也扮演重要角色[5- 6],但对其在水分入渗和蒸发方面的研究结论还存在较大争议,主要观点为结皮对水分的入渗和蒸发过程具有促进作用[7- 10]、抑制作用[10-12]及无作用[13-16]3种观点。
藓类结皮作为生物土壤结皮的高级阶段,是生物土壤结皮的重要组成部分和生物量的最主要贡献者。与维管束植物不同,藓类植物多不具有维管束,属于典型的变水植物,能够随环境水分的变化而变化[17],在截留降水、涵养水源等方面扮演重要角色[18-21]。尤其对于荒漠藓类植物而言,为应对干旱少雨的恶劣环境,荒漠藓类植物在个体水平和群体水平上均表现出较强的环境适应性[22-26]。研究发现,在干燥环境中齿肋赤藓(Syntrichiacaninervis)叶片紧贴于茎,通过减少暴露于空气的表面积来减少水分蒸发,而其毛尖结构能够收集空气中的水分,较无毛尖齿肋赤藓毛尖能够多收集10.26%的凝结水量[27]。在群体水平上,荒漠藓类植物密集丛生并呈现垫状分布,既提高了土壤毛细管系统的持水力,也在其表面形成了一个静止层,减弱了水分蒸发速率[28]。
荒漠藓类植物叶片多为单层细胞,对环境变化十分敏感,被誉为环境变化的指示剂。近几十年来诸如增温、氮沉降及极端天气等事件的发生致使荒漠藓类植物生存受到严重威胁。研究发现,荒漠藓类植物对环境变化的敏感性要远高于相同生境下的维管束植物,3 g N m-2a-1以上的氮沉降及夏季频繁的小降水事件均会导致荒漠藓类植物的死亡[29-31]。然而,荒漠藓类植物的死亡这一过程是否会影响表层土壤的水分蒸发和入渗目前尚不清楚,因此,我们提出科学问题:荒漠结皮层藓类植物死亡是否对表层土壤水分蒸发和入渗存在影响,若存在是如何影响的?为解答此科学问题,我们选取古尔班通古特沙漠优势藓类植物齿肋赤藓为研究对象,以裸沙为对照,通过对比活藓类结皮与藓类植物死亡结皮层对土壤水分入渗和蒸发的影响,以期为深入探讨荒漠藓类结皮对荒漠生态系统水文过程的影响提供理论依据。
1 材料与方法
1.1 研究区概况
古尔班通古特沙漠是我国最大的固定半固定沙漠,海拔300—600 m,面积为4.88 ×104km2。年平均气温7.19℃,极端最高气温41.5℃;多年平均降水量128.6 mm,年均蒸发量大于2000 mm。该沙漠冬季具有稳定的积雪,积雪厚度通常在20 cm左右,积雪覆盖持续100—160 d。该沙漠地表广泛发育着生物土壤结皮和草本植物层片,早春积雪融化为生物土壤结皮及草本植物的生长提供了充足的水分。同时,维管束植物与生物土壤结皮镶嵌分布是该沙漠主要地表景观之一,其中生物土壤结皮主要包括藻结皮、地衣结皮和藓类结皮3类,藻结皮多分布于沙垄的中上部,地衣多分布于沙陇中下部及丘间低地,而藓类结皮主要分布在丘间低地[32]。
1.2 样品采集及处理
在丘间低地随机选取发育良好且生长相对一致的以齿肋赤藓为优势种的藓类结皮,用PVC管(内径10 cm,高度20 cm)垂直插入沙土中,小心取得未扰动的藓类结皮样品10份,在裸沙处,以同样方法取得5份样品,封底后运回至实验室备用。为避免采样时对藓类结皮结构的扰动,采样前用喷壶将结皮表面喷湿。在实验室,采用65℃湿热处理4 h将藓类结皮灭活,设置5个重复。室温放置24 h后,复水30 min,利用Mini-PAM 2500 (Walz, Hernz, Germany) 测定其荧光活性,确认其完全处于死亡状态;存活结皮组与裸沙组施加等量的水分,各设置5个重复。将所有样品放至温度23℃、湿度16%的培养箱中自然蒸发15 d,以保证所有样品中具有相同的初始含水量。
1.3 水分入渗与蒸发测定
1.3.1藓类结皮水分入渗的测定
采用Mini-Disk Infiltrometer (Decagon公司,美国)便携式渗透计测定入渗速率,将渗透计底部多孔烧结钢盘平行放置于藓类结皮表面,选择适于藓类结皮的4 cm (相当于0.4 kpa)压头,记录储水室初始水量后,间隔30 s记录一次储水室的水量,以时间的平方根为x,累积入渗深度为y,然后将得到的数据拟合到Zhang[33]提出的描述圆盘式渗透计渗透情况公式中,得到C1和C2常数值。
y=C1x1/2+C2x
利用得到的C2计算水分入渗速率:
K=C2/A
式中,A为给定土壤类型的van Genuchten参数与吸力率和入渗仪圆盘半径之间的关系值。
累积入渗量=Vt-Vf(i)
式中,Vt为储水室初始水量,Vf(i)为第i次测量时储水室的水量。
1.3.2藓类结皮水分蒸发测定
研究发现齿肋赤藓在春季固定了其全年固碳量的50%以上[34],因此,本研究选择了冻融季节的20 mm降水量来模拟测定齿肋赤藓死亡对水分蒸发的影响[35]。将20 mm降水用小型喷壶均匀喷洒于蒸发仪[16]PVC管内的齿肋赤藓表面,称量其初始重量。将所有样品放置于室温为23℃的温室内培养,在192 h内每间隔8 h测定一次样品重量,192 h后间隔12 h测定一次,直至恒重,结合蒸发时间和蒸发量计算蒸发速率。
蒸发速率=(Wf(i)-Wf(i-1))/t
累积蒸发量=Wt-Wf(i)
式中,Wt为蒸发仪加水后的初始重量,Wf(i)为第(i)次测量时蒸发仪重量,Wf(i-1)为第(i-1)次测量时蒸发仪重量,t为第(i)次测量与第(i-1)次测量的时间间隔。
1.4 数据处理
用Excel对所得的数据进行常规处理。使用 SPSS 25.0软件分别对活藓类结皮与死亡藓类结皮及裸沙的水分蒸发、入渗数据进行正态性检验和方差齐性检验,随后对这3种地表类型间的水分蒸发、入渗数据进行单因素方差分析(one-way ANOVA)和多重比较(LSD)。采用Origin 9.0软件进行绘图。
2 实验结果
2.1 藓类结皮存活与否对土壤水分入渗速率的影响
3种不同地表类型在初始阶段入渗速率差异较大,裸沙入渗速率表现为由最大速率逐渐降低,活藓类结皮入渗速率表现为先增大后减小,而藓类植物死亡的结皮层入渗速率则表现为逐渐增大,活藓类结皮较藓类植物死亡的结皮层及裸沙更快进入稳定入渗阶段(图1;表1)。在初始入渗阶段和稳定入渗阶段,与裸沙相比活藓类结皮水分入渗速率均降低了近85%。同时,藓类植物死亡的结皮层进一步限制了水分入渗,初始阶段入渗速率较活藓类结皮降低了60.11%,即使在稳定入渗阶段,藓类植物死亡的结皮层水分入渗率也只达到活藓类结皮的85.91%。
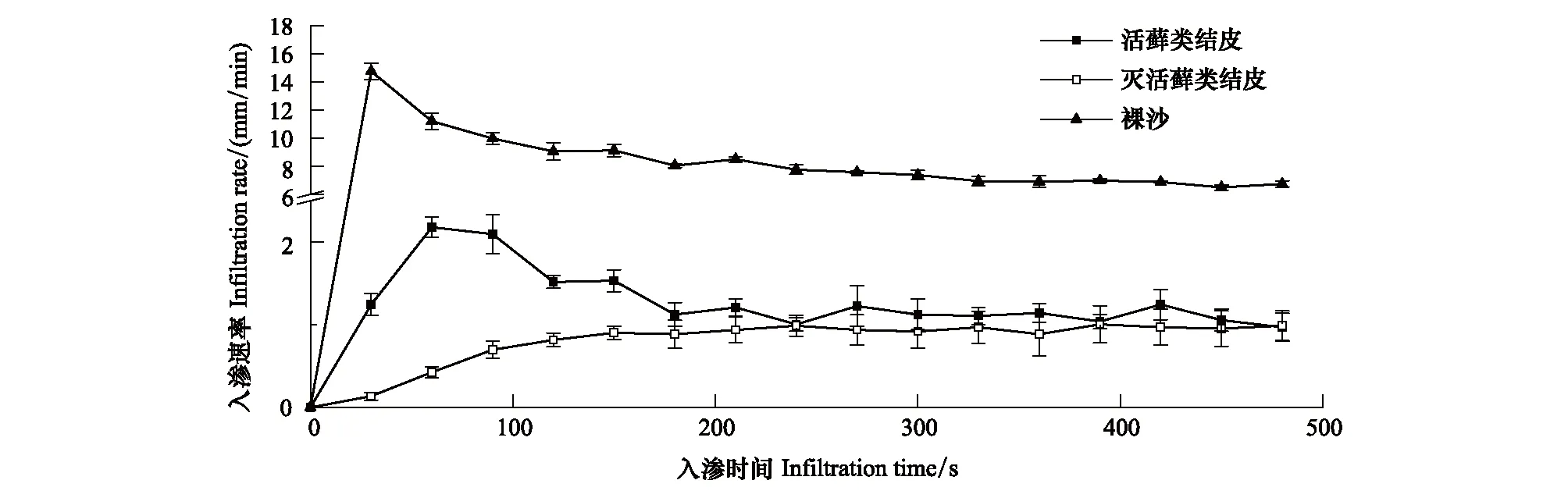
图1 3种地表类型的水分入渗速率Fig.1 Infiltration rate of three surface types

表1 3种地表类型入渗特征参数
2.2 藓类结皮存活与否对土壤水分累积入渗量的影响
与入渗速率结果一致,活藓类结皮的累积入渗量也显著低于裸沙,结皮层藓类植物死亡后其累计入渗量显著低于活藓类结皮,且其抑制作用最大,累积入渗量分别是活藓类结皮的64.48%,仅为裸沙的20.42%(图2,P<0.05)。

图2 3种地表类型的水分累积入渗量Fig.2 Cumulative water infiltration of three surface types
2.3 藓类结皮存活与否对土壤水分蒸发速率的影响
由图3可知,裸沙、活藓类结皮及藓类植物死亡的结皮层蒸发过程均经历了常速率、减速率、残余速率3个阶段,但3种不同地表覆盖类型在各蒸发阶段时长具有显著性差异。常速率阶段藓类植物死亡的结皮层蒸发时间(70 h)显著大于活藓类结皮(34 h)及裸沙(40.4 h),减速率阶段表现为活藓类结皮蒸发时间(141 h)显著大于藓类植物死亡的结皮层(96.8 h)及裸沙(84.8 h),残余速率阶段藓类植物死亡的结皮层(189.2 h)及裸沙(201.2 h)蒸发时间显著高于活藓类结皮(145 h)。在整个蒸发过程中,活藓类结皮在常速率阶段较裸沙平均蒸发速率降低31.03%,而残余速率阶段蒸发速率则加快30.59% (图4)。

图3 3种地表类型的水分蒸发速率Fig.3 Water evaporation rate of three surface types
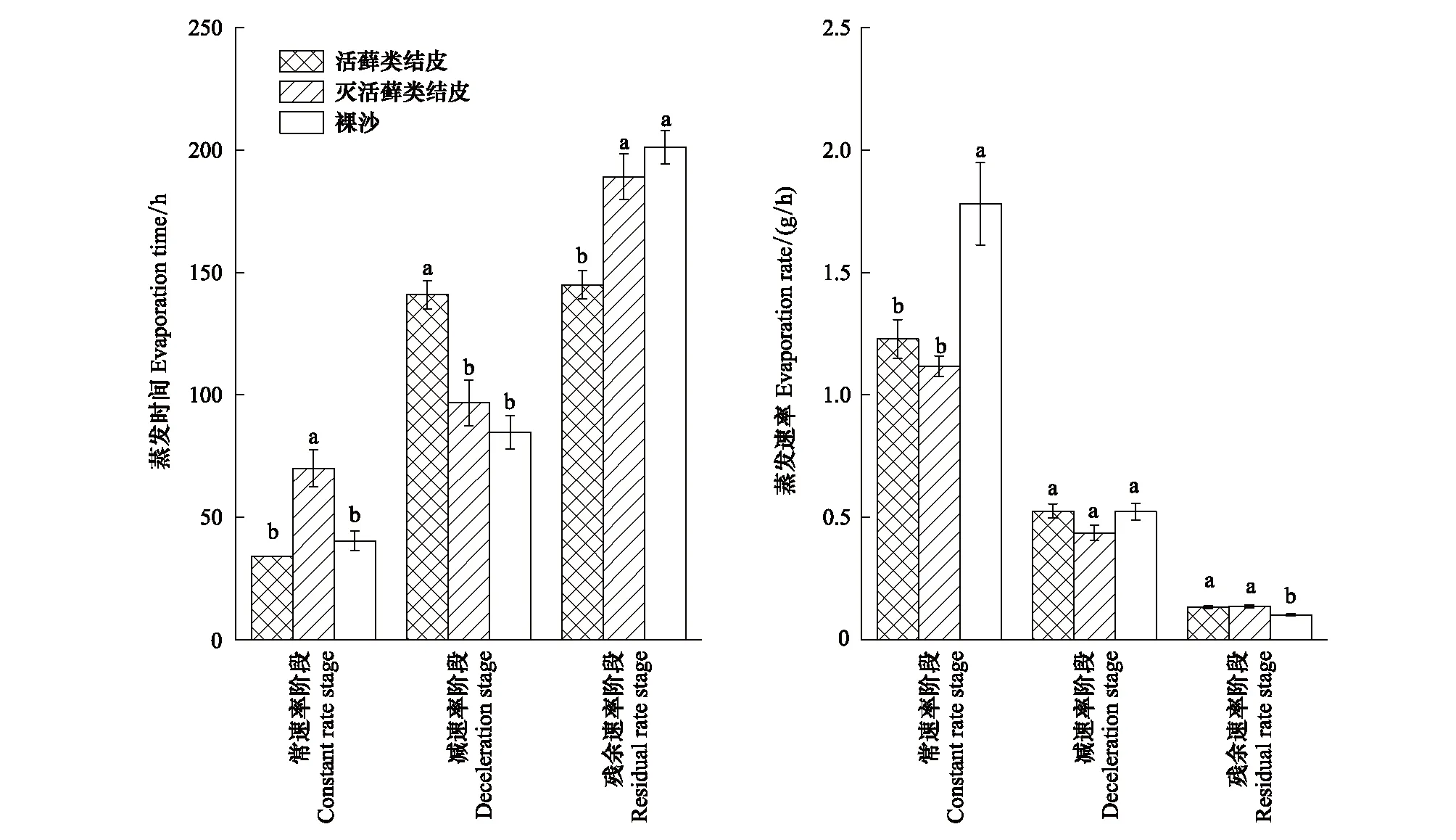
图4 3种地表类型蒸发过程中蒸发时间和速率的差异性Fig.4 Differences in duration and rate of evaporation among three surface types during evaporation processes不同字母表示同一速率下不同地表类型之间在0.05概率水平上存在显著差异(P<0.05)
2.4 藓类结皮存活与否对土壤水分累积蒸发量的影响
在累计蒸发量方面,藓类植物死亡的结皮层与活藓类结皮没有显著性差异 (图5)。与裸沙相比,藓类结皮在前期减少了土壤水分累积蒸发量,而在蒸发后期则呈现相反趋势,表现为藓类结皮显著大于裸沙,尤其是藓类植物死亡的结皮层累计蒸发量最大。
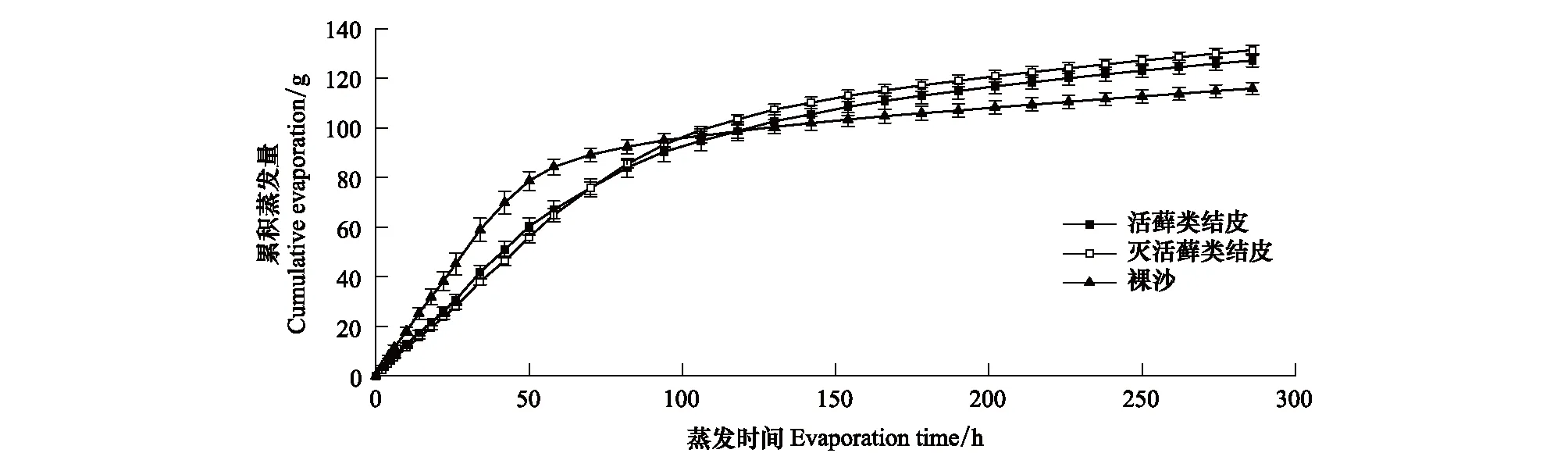
图5 3种地表类型的水分累积蒸发量Fig.5 Cumulative water evaporation of three surface types
3 讨论与结论
生物土壤结皮是荒漠地表的重要覆被类型,能够显著影响地表土壤水文过程[36-37]。以往在生物土壤结皮对土壤水文过程影响的研究中,尤其是在水分入渗及蒸发方面一直存在争议,主要观点为结皮对水分的入渗和蒸发过程具有促进作用[7- 10]、抑制作用[10-12]及无作用[13-16]3种观点。这可能由于研究地区、测定指标及所选生物土壤结皮类型不同导致。针对藓类结皮而言,主流观点认为藓类结皮的存在可造成土壤水分浅层化,且随着结皮盖度增加水分入渗深度越浅[12, 38- 39]。这在本研究中也得到了证实,即藓类结皮的存在显著阻碍了水分的入渗,其原因可能与藓类结皮层细沙含量较高有关[12, 40],藓类植株及其与微生物分泌的多糖物质通过吸水膨胀,堵塞土壤孔隙且增强覆盖土壤表面的能力[8, 41],大大提高了水分的滞留时间,这可以降低入渗速率,从而显著影响土壤的含水量。
在荒漠生态系统中,蒸散是水平衡的关键过程[42-43],超过90%的年降水量会因这一过程而损失[44]。研究发现,蒸发过程主要受地表覆盖类型的影响,例如植被类型和生物土壤结皮等[45]。本研究结果显示,在20 mm降雨量条件下,藓类结皮覆盖对水分蒸发的影响随不同蒸发阶段而异,这与前期诸多学者研究结果相似,认为生物土壤结皮对水分蒸发不是简单促进或抑制[16, 46-47]。在蒸发前期,齿肋赤藓表面蜡质结构、叶片毛尖及毛尖间形成的交叉网络降低了水分蒸发,且群体水平上垫状的分布使其表面形成了一个空气静止层,减弱了水分蒸发。而在蒸发后期,藓类结皮的存在却提高了蒸发速率,这可能由于齿肋赤藓已进入休眠状态,降低保水能力,且藓类植物的覆盖形成了毛细管作用,从而促进了土壤水分的蒸发,而裸沙表面形成的硬壳状土壤干燥层可能是造成蒸发后期裸沙蒸发速率较低的主要原因。值得注意的是,在蒸发测量结束后,藓类结皮土壤水分累积蒸发量显著大于裸沙,结合藓类结皮对水分入渗的抑制作用,可以得知藓类结皮的存在显著了降低土壤含水量。
长期增温和夏季频繁小降水均会造成藓类植物的死亡,进而影响荒漠地表的水分和养分。研究发现,不同时间序列的长期增温(6年和10年)均会显著降低藓类结皮对水分的截留率[5]。但在本研究中发现,活藓类结皮相比藓类植物死亡的结皮层显著降低水分的入渗速率和累计入渗量。这可能与藓类植物死亡时间长短造成的藓类结皮盖度差异有关,长期增温实验导致苔藓盖度明显降低,而本研究中采用的是即时死亡藓类植物,不存在盖度变化。但结皮层藓类植物死亡造成的水分入渗降低是否与藓类死亡后其表面疏水/吸水性物质变化、抑或藓类毛尖、茎叶微结构变化有关还需要进一步深入研究。
生物土壤结皮作为干旱半干旱区重要的地表覆被类型,其重要的作用之一是对荒漠土壤水文过程的影响,我们的研究结果也强化了这种观点。值得注意的是,尽管藓类植物的死亡显著降低了水分的入渗,但与活藓类植物相比其累计蒸发量无显著差异,甚至在蒸发后期略高于活藓类结皮。前期有研究发现,干旱、半干旱区生物土壤结皮会通过影响生态系统的水文过程而造成维管植物的退化[48],尤其是以苔藓为主的生物土壤结皮能显著降低土壤湿度而导致人工种植灌木的死亡[49]。综合考虑藓类植物死亡对水分入渗的抑制,而对蒸发的无影响、甚至略有促进的研究结果,可以推测齿肋赤藓的死亡在短期内会显著降低土壤含水量,进而影响与其共存维管植物的生存。全球气候的持续变暖已成为不争的事实,降水及其时空模式也随之出现不稳定的变化,研究预测显示在本世纪末土壤含水量将减少5%—15%,全球干旱区的干旱化程度不断加剧[50- 52]。以荒漠藓类植物为代表的生物土壤结皮对气候变化的响应要远大于维管束植物,在全球气候变化和土地利用方式的变化下,至2070年覆盖地球陆地面积约12%的生物土壤结皮将减少25—40%,显著影响荒漠地表的水文过程和养分循环[53]。因此,探究荒漠藓类植物死亡后的生态效应对预测荒漠植物变化趋势和保护荒漠植物物种多样性和地表稳定性具有重要生态价值。
致谢:陆永兴同学帮助写作,刘鑫、梅月磊和张庆同学帮助实验及数据处理,特此致谢。
猜你喜欢
求学·理科版(2023年6期)2023-04-12 18:35:48
新疆大学学报(自然科学版)(中英文)(2022年3期)2022-06-04 13:45:40
草业学报(2021年6期)2021-06-28 11:01:36
食品工业(2021年4期)2021-05-08 08:41:52
水土保持通报(2020年6期)2020-03-10 01:46:52
广西植物(2020年11期)2020-01-21 05:59:46
华东师范大学学报(自然科学版)(2019年3期)2019-06-24 05:29:09
科学Fans(2019年2期)2019-04-11 01:49:38
新疆大学学报(自然科学版)(中英文)(2018年4期)2018-02-10 00:09:15
淮阴师范学院学报(自然科学版)(2017年4期)2018-01-12 09:56:26
