
黄土高原不同发育阶段生物结皮的导水和持水特征
2021-06-28 11:01:36孙福海肖波李胜龙王芳芳
草业学报 2021年6期
孙福海,肖波,2*,李胜龙,王芳芳
(1.中国农业大学土地科学与技术学院,北京100193;2.中国科学院水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西杨凌712100)
在干旱和半干旱地区,土壤中的水分是制约植物生长的主要生态因子,也是植被群落结构和生态系统功能的重要限制因素[1-2]。土壤导水是指水分渗出土壤的过程,土壤持水是指土壤所能保持的含水量,分别常用饱和导水率和田间持水量进行表征。土壤导水和持水性不仅决定着土壤中的水分状况,而且对物质循环、植物发育、地表径流和侵蚀过程有重要影响[3]。作为计算水量平衡、评估作物生长和模拟溶质运移等过程的重要参数,土壤导水和持水性的大小受土壤质地、结构、孔隙度等土壤性质的综合影响[4]。
生物结皮是生长在土壤表面及以下的藻类、地衣、苔藓、真菌和细菌等生物同土壤颗粒相互作用形成的复合层,是草地生态系统的“工程师”[5-6]。生物结皮作为一种特殊的下垫面,显著改善土壤理化性质,增加土壤有机物质,为沙地或荒漠植被演替提供养分[7]。国内外学者对生物结皮渗透特性展开了较多研究,受结皮特殊属性(质地、孔隙、结构等)的影响,研究结果差异较大[8-9]。一方面,生物结皮在形成过程中使表层孔隙不同程度地被细颗粒物质填充,增加了土壤粉粒和黏粒含量,减少了土壤中非毛管孔隙度,从而降低土壤入渗能力[10-11];另一方面,生物结皮通过假根和菌丝增加表层土壤大孔隙数量,其表层植物体间的间隙捕获了更多的径流,为水流下渗提供了良好通道,提高了土壤导水率[12];除此之外,Wang等[13]对比灌木群落下生长的生物结皮破坏前后的土壤入渗速率的差异,发现两者无显著差异。Williams等[14]研究表明,生物结皮破坏前后的土壤入渗速率差异不显著。分析以上研究结果可知,由于前人研究所选土壤质地、生物结皮类型、测定方法以及对照类型均存在差异,导致所得结果不尽相同。因此,不能将以上研究结论应用于其他研究区域的生物结皮水分研究中。
生物结皮的发育可以改善土壤的持水性[15]。王新平等[16]认为生物结皮的持水能力为沙丘沙的3~9倍,显著改善了土壤水分的有效性。Gypser等[17]研究表明,随着绿藻的生长,生物结皮的发育显著提高了土壤的饱和含水量。此外,结皮的发育阶段是影响土壤水分状况的重要因素[15]。而生物结皮的导水和持水性决定着其对干旱和半干旱地区浅层土壤水分有效性的影响,进而影响土壤微生境。
黄土高原地处我国半湿润向干旱荒漠的过渡地带,水资源匮乏,土壤侵蚀严重,生态系统脆弱,为有效控制水土流失,黄土高原实施了退耕还林(草)工程,随着该工程措施的大力推广,生物结皮成为该区最具特色的微自然景观[7]。目前,关于黄土高原地区生物结皮导水和持水特征随结皮演替变化的研究较少,其变化机理尚不明确。基于此,为探究黄土高原地区生物结皮对土壤导水和持水性的影响,对比不同发育阶段生物结皮覆盖下土壤导水和持水性的差异,进而揭示生物结皮对区域水分的影响。本研究针对黄土高原风沙土上不同发育阶段生物结皮(藻结皮、藻-藓混生结皮、藓结皮),通过定水头法和威尔科克斯法分别测定生物结皮的饱和导水率、水分入渗过程、田间持水量等指标,比较不同发育阶段生物结皮和裸沙水分渗透参数和持水参数的差异,分析不同发育阶段生物结皮对水分入渗过程的影响,以期完善生物结皮在土壤水分效应方面的研究,为进一步明确生物结皮在减缓黄土区水土流失中的作用提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于陕西省神木市六道沟小流域(110°21′-110°23′E,38°46′-38°51′N),流域面积为6.89 km2,海拔高度为1094.0~1273.9 m。该地区位于晋陕蒙三省接壤区,地理位置十分特殊,既属于黄土高原向毛乌素沙漠过渡、森林草原向典型干旱草原的过渡地带,又是流水侵蚀的黄土丘陵区向干燥剥蚀作用的鄂尔多斯高原过渡的水蚀风蚀交错带,同时也是典型的农牧交错带和生态环境脆弱区。该区属于中温带半干旱气候,年均气温8.4℃,年均降水量408.5 mm,降水多集中在6-9月。整个小流域的地面组成物质主要为第四纪黄土沉积物,西侧以砂土为主,占流域总面积的13.5%;东侧以沙壤土为主,占流域总面积的86.5%[18]。生物结皮是该地区最常见的地表微景观,不同土地利用类型下(草地、林地和农地等)都可以发现大量的生物结皮。研究区内代表性植物群落有苜蓿(Medicago sativa)、柠条(Caragana korshinskii)、长芒草(Stipa bungeana)、达乌里胡枝子(Lespedeza davurica)、茭蒿(Artemisia giraldii)和阿尔泰狗娃花(Heteropappus altaicus)等。
1.2 试验设计
在野外调查的基础上,于2019年7-8月进行野外样品采集和室内试验。生物结皮按照其演替阶段可以大致分为物理结皮、藻结皮、藻-藓混生结皮和藓结皮,本研究区以藻结皮、藻-藓混生结皮和藓结皮为主。选取风沙土上不同发育阶段生物结皮,并以相邻的裸沙(原状土)作为对照(距离在10 m以内),共4个处理[裸沙(CK)、藻结皮(A)、藻-藓混生结皮(B)、藓结皮(C)]。
1.3 样品采集与基本理化性质测定
采用标准环刀(高5 cm,体积100 cm3)对上述4种处理的0~5 cm和5~10 cm土层进行取样,用以测定饱和导水率(saturated hydraulic conductivity,Ks)和田间持水量(field capacity,Fc),每个土层3个重复;采用有机玻璃管(内径5 cm、高30 cm)对4种处理取样后带回室内测定入渗过程,所取结皮表层均完整无裂隙,每种处理3个重复。此外,在采样点附近取相应扰动土壤,带回实验室后用于测定土壤基本理化性质。
采用烘干法测定生物量[19];采用游标卡尺测量生物结皮的厚度[20];采用环刀法测定土壤容重(ρb),按照(1-ρb/2.65)×100%计算土壤总孔隙度[20];采用马尔文激光粒度仪(Mastersizer 2000,Malvern Instruments Ltd.,UK)测定土壤机械组成[21],并按美国制进行分级(砂粒粒径0.05~2 mm、粉粒粒径0.002~0.05 mm和黏粒粒径0~0.002 mm)。
1.4 室内入渗与持水性试验
采用环刀定水头法[22]测定土壤Ks,计算方法见公式(1):

式中:Q为出流量(mm3);L为土壤厚度(mm);S为出流横截面积(mm2);Δt为出流时间(min);h为水层高度(mm);T为试验过程中的平均水温(℃)。
采用土柱定水头法测定原状土壤入渗性能。测定过程中,利用马氏瓶加水维持土面3 cm的积水深度,分时间段记录马氏瓶加入水量。入渗0~5 min每隔30 s记录1次,入渗5~30 min每隔1 min记录1次,入渗30 min后每隔5 min记录1次,当至少3次相同时间内马氏瓶供水量恒定时可视为达到稳定,停止试验,测定入渗时间不少于70 min。入渗速率计算方法见公式(2):

式中:i(t)为入渗速率(mm·min-1);ΔV为某一时段马氏瓶加入水量(mm3);S′为土样横截面积(mm2),Δt为时段(min),T为试验过程中的平均水温(℃)。
本研究采用初始入渗速率、稳定入渗速率、平均入渗速率和累积入渗量4个指标用来分析土壤入渗性能。其中初始入渗速率指入渗前3 min的入渗速率;稳定入渗速率为单位时间内的入渗量趋于稳定时的入渗速率;平均入渗速率为达到稳渗时的累积入渗量与达到稳渗所用时间的比值;累积入渗量为一定时间内的累积入渗总量,本试验取60 min内的入渗总量为累积入渗量[23]。
采用威尔科克斯法测定Fc,重点研究野外田间土壤在饱和后的释水过程。采用2个标准环刀(高5 cm,体积100 cm3)对田间土壤进行取样,其中一个环刀土样饱和24 h后作为上环刀,另一个环刀土样自然风干后作为下环刀。上环刀取底盖放置滤纸后与下环刀对接。每隔4 h称重1次并计算2次含水量的差值,待差值小于1%时停止试验。本研究取饱和后24 h和饱和后释水8 h的土壤含水量分别作为饱和含水量(saturated water content,θs)和Fc。
1.5 数据处理
使用Microsoft Excel 2016对数据进行处理与分析,采用IBM SPSS 22.0进行单因素方差分析和Pearson相关分析,通过Origin Pro 2019作图。
2 结果与分析
2.1 不同发育阶段生物结皮对土壤理化性质的影响
如表1所示,研究区藻、藻-藓混生、藓3种类型生物结皮样地的土壤机械组成均以砂粒为主,随着生物结皮的发育和演替,生物结皮中砂粒含量不断降低,而粉黏粒含量、结皮厚度和生物量不断增加。其中,与藓结皮相比,0~5 cm土层的藻结皮和藻-藓混生结皮的砂粒含量分别增加了2.8%和0.7%,黏粒含量分别降低了81.0%和72.4%。
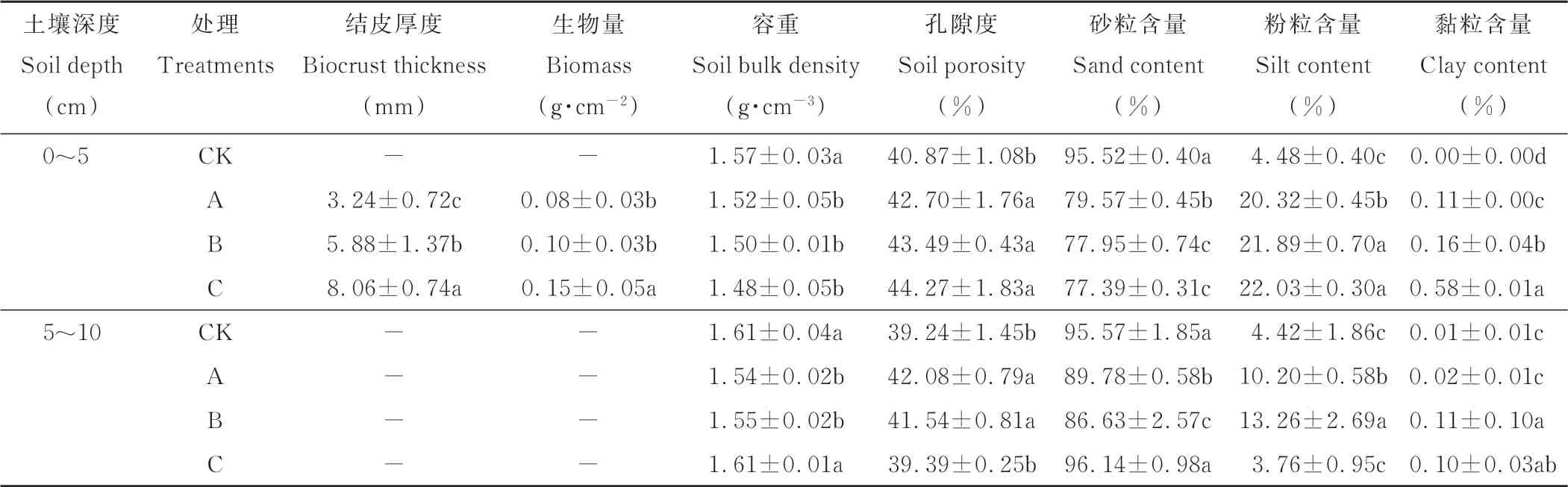
表1 不同发育阶段生物结皮和无结皮的土壤样品理化性质Table 1 Soil physicochemical proper ties of the biocrusts in differ ent developmental stages and bare soil
与裸沙相比,生物结皮的发育显著增加了表层生物量和黏粒含量,降低了砂粒含量。其中,在0~5 cm土层,与裸沙相比,藻结皮、藻-藓混生结皮和藓结皮的容重分别降低了3.2%、4.5%和5.7%,粉粒含量分别增加了3.53、3.89和3.92倍;在5~10 cm土层中,与裸沙相比,不同发育阶段生物结皮覆盖下土壤孔隙度增加了0.4%~7.2%,黏粒含量增加了1.0~10.0倍。此外,与0~5 cm土层相比,生物结皮对5~10 cm土层理化性质的影响较小。
2.2 不同发育阶段生物结皮对土壤Ks的影响
由图1可知,生物结皮(0~10 cm)的Ks较裸沙相比平均降低了49.9%,但不同发育阶段生物结皮的影响程度不同。其中,0~5 cm土层,藻结皮、藻-藓混生结皮和藓结皮的Ks分别为0.92、0.88和1.55 mm·min-1,与裸沙相比分别降低了62.1%、63.8%和36.3%;藻结皮、藻-藓混生结皮和藓结皮覆盖下5~10 cm土层的Ks分别为1.31、1.15和2.44 mm·min-1,较裸沙分别下降了56.4%、62.0%和18.8%。除藻结皮与藻-藓混生结皮间差异不显著,在其余处理两者之间差异均显著(P<0.05),且生物结皮对表层土壤(0~5 cm)Ks的影响大于下层土壤(5~10 cm)。
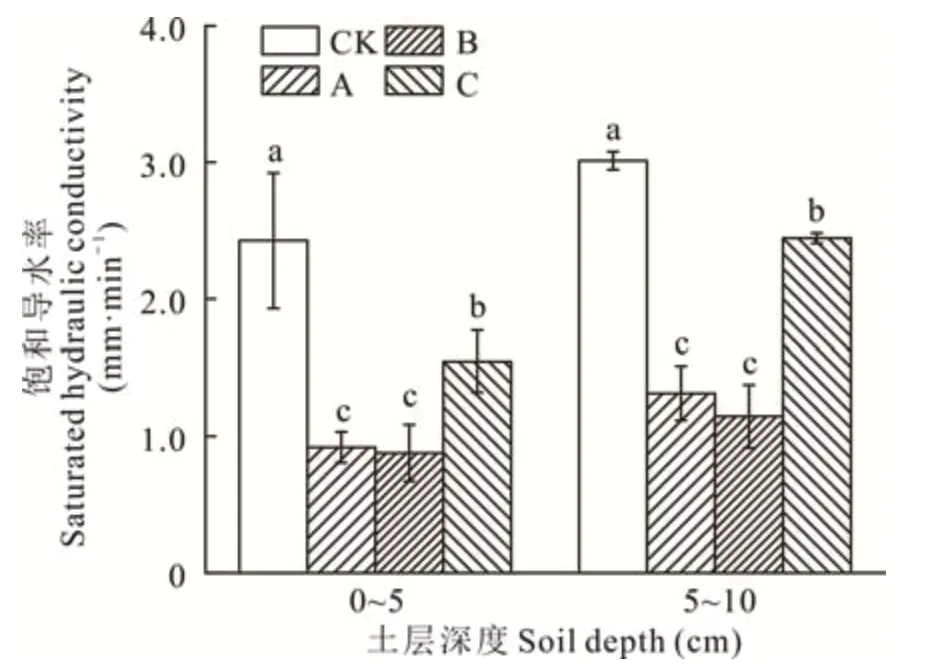
图1 不同发育阶段生物结皮和无结皮覆盖下不同深度土层的饱和导水率Fig.1 Saturated hydraulic conductivity of the biocrusts in different developmental stages and bare soil at different depths
2.3 不同发育阶段生物结皮对水分入渗过程影响
由图2可知,不同发育阶段生物结皮的入渗速率曲线相似,均呈现“L”型。0~5 min入渗速率在初期均较高,随着入渗时间的推移入渗速率急剧下降;5~20 min,入渗速率缓慢下降;20~70 min入渗速率基本趋于稳定。表2显示,不同发育阶段生物结皮均会阻碍土壤的水分入渗过程,但影响程度不同。与裸沙相比,藻结皮的初始入渗速率、稳定入渗速率、平均入渗速率和累积入渗量分别降低了43.9%、35.8%、37.3%和33.8%,平均降低了37.7%;藻-藓混生结皮的初始入渗速率、稳定入渗速率、平均入渗速率和累积入渗量较裸沙分别降低了32.3%、66.4%、62.0%和56.6%,平均降低了54.3%;藓结皮的初始入渗速率、稳定入渗速率、平均入渗速率和累积入渗量较裸沙分别降低了22.0%、23.1%、14.6%和13.9%,平均降低了18.4%。综上,不同发育阶段生物结皮的渗透性随着结皮演替呈先降低后增加的变化规律,即藓结皮>藻结皮>藻-藓混生结皮。
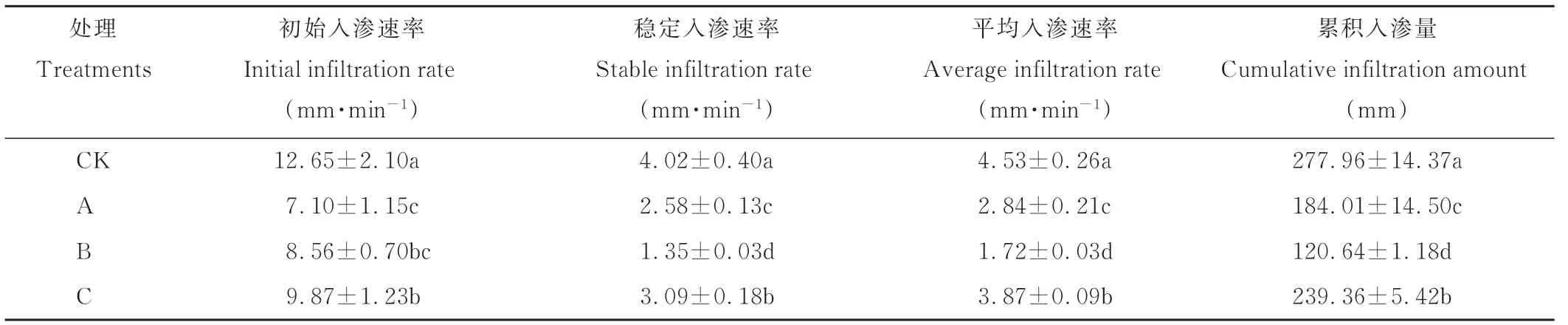
表2 不同发育阶段生物结皮覆盖土壤和无结皮的水分入渗参数Table 2 Infiltration parameters of the biocrusts in different developmental stages and bare soil
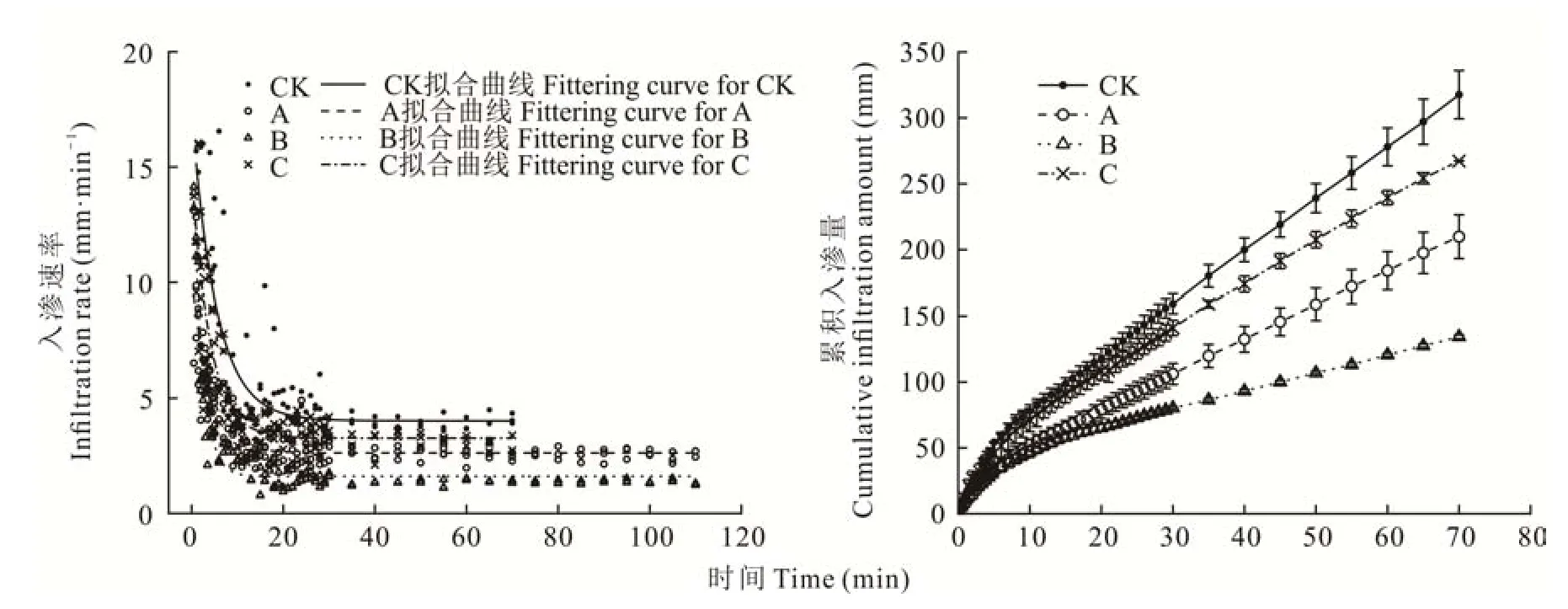
图2 不同发育阶段生物结皮和无结皮覆盖土壤的水分入渗过程与累积入渗量Fig.2 Infiltration process and cumulative infiltration amount of the biocrusts in different developmental stages and bare soil
2.4 不同发育阶段生物结皮对土壤持水性的影响
不同发育阶段生物结皮覆盖下0~10 cm土层的退水过程曲线表现为土壤含水量均在0~4 h急剧下降,4~72 h变化较为平缓,72 h后基本趋于稳定(图3)。在退水过程中,与裸沙相比,不同发育阶段结皮覆盖下0~5 cm土层均显著提高了土壤的持水量(P<0.01),但不同发育阶段生物结皮的影响略有差异。其中,藻结皮、藻-藓混生结皮和藓结皮72 h的体积含水量分别为0.27、0.29和0.22 cm3·cm-3,较裸沙相比分别增加了1.45、1.67和0.99倍,平均增加了1.37倍。在5~10 cm土层中,藻结皮、藻-藓混生结皮和藓结皮72 h的体积含水量较裸沙相比分别增加了1.26、1.74和0.61倍。此外,在整个退水过程中(0~120 h),藻结皮、藻-藓混生结皮和藓结皮(0~10 cm)的平均土壤含水量较裸沙分别增加了1.14、1.40和0.74倍。
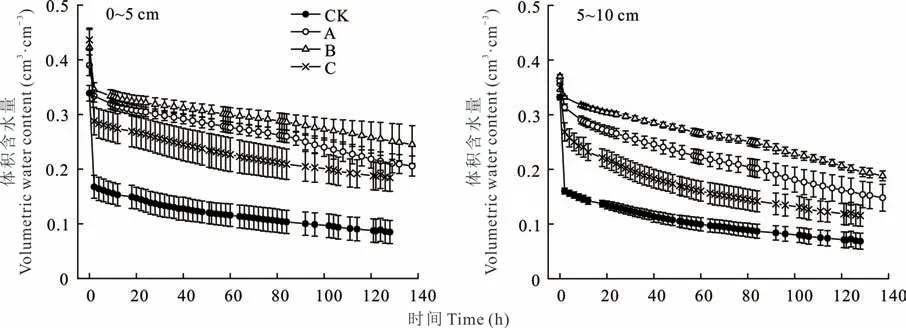
图3 不同发育阶段生物结皮和无结皮的退水曲线Fig.3 Change of soil water content of biocrusts in different developmental stages and bare soil
由图4可知,不同发育阶段生物结皮均可显著提高土壤的θs和F(cP<0.047)。在0~5 cm土层中,藻结皮、藻-藓混生结皮和藓结皮的θs分别为0.39、0.42和0.44 cm3·cm-3,与裸沙相比分别增加了15.1%、24.9%和28.8%。而5~10 cm土层的θs分别增加了9.0%、11.5%、5.4%;在0~5 cm土层中,与裸沙相比,藻结皮、藻-藓混生结皮和藓结皮的Fc分别增加了1.03、1.12和0.77倍。5~10 cm土层中的Fc分别增加了0.92、1.09和0.64倍。综上,不同发育阶段生物结皮的持水性表现为藻-藓混生结皮>藻结皮>藓结皮。
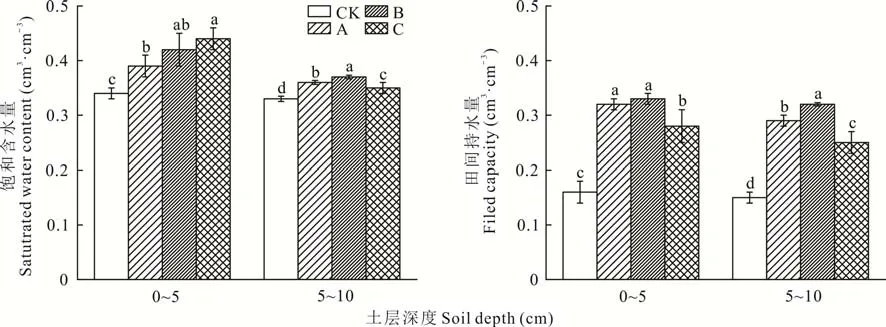
图4 不同发育阶段生物结皮和无结皮覆盖下0~10 cm土层的饱和含水量和田间持水量Fig.4 Saturated water content and field capacity of the biocrusts in different developmental stages and bare soil at 0-10 cm depth
3 讨论
生物结皮作为广泛分布于干旱和半干旱地区土壤表层的特殊复合体,其从形成到稳定的演替呈现物理结皮-藻结皮-藻-藓混生结皮-藓结皮的发育规律[24]。本研究表明随着生物结皮演替,生物结皮的砂粒含量逐渐降低,粉、黏粒含量增加,其可能主要与生物结皮中的藻丝体有关,藻丝体与沙土颗粒直接接触,进而束缚沙土颗粒形成团状物[20],同时,它们也向胞外分泌并积累多糖聚合物(extracellular polymeric substance,EPS)[25],从而进一步胶结沙土颗粒,形成更稳定的团聚体,进而改善土壤机械组成,使得黏粒和有机质含量增加,进而影响土壤的导水和持水特征[26-27]。
本研究中,与裸沙相比,不同发育阶段生物结皮均阻碍了土壤水分渗透。这与其他研究结果有相似之处。例如,Xiao等[10]研究表明,生物结皮在积水入渗条件下的稳定入渗速率和Ks均显著低于无结皮土壤,并认为土壤颗粒分布和结皮厚度是影响入渗的关键因素;李渊博等[20]研究发现,与裸地相比,风沙土和黄绵土上发育的藓结皮降低了土壤Ks,并认为容重与有机质含量为关键影响因素。这些研究均一致表明:生物结皮的发育降低了土壤渗透性。这一方面可能与生物结皮的物理特性有关,首先,生物结皮具有一定的成土作用,促进土壤团聚体的形成,改善土壤结构,从而影响水分在土壤中的渗透速率[7];其次,生物结皮发育后使土壤表层孔隙被细颗粒物质填充,在地表形成致密层,阻碍水分入渗[28-29]。另一方面,该结果可能与生物结皮的生物特性相关,生物结皮层的细菌可以产生大量的胞外聚合物[30],其吸水膨胀后会形成不透水层,进而堵塞地表孔隙,减少水分在孔隙中的流通,最终降低水分的下渗[10]。本研究中,不同发育阶段的生物结皮对导水性的影响不尽相同,生物结皮的渗透性随着其发育演替呈先降低后增加的变化趋势。其中,藻结皮与藻-藓混生结皮减渗作用更显著,其主要原因可能在于其表面结构的差异,相对于藓结皮,藻结皮与藻-藓混生结皮的表层结构更封闭,结皮层越封闭,降低入渗的作用更加明显。而藓结皮表层以单株藓类植物体密集丛生为主,其株间间隙为水流进入下层土壤提供了良好通道[24]。同时,藓结皮遇水发生膨胀后可进一步增加株间间距,导致减渗作用较弱。另一方面,生物结皮斥水性随着藻结皮的发育逐渐增加,而藓结皮发育后斥水性逐渐降低,从而可能间接导致渗透性呈先降低后增加的趋势[10,17]。
本研究表明,黄土高原不同发育阶段生物结皮均提高了土壤持水性,具体表现为显著增加了表层土壤(0~5 cm)的θs和Fc,由于结皮层自身较薄,其对5~10 cm土层的影响有限。这主要是由于生物结皮形成后改变了表层土壤的物理性质,首先,生物结皮发育后增加了土壤粗糙度[31],有利于土壤表面获得更多的降尘,在生物结皮的遮蔽保护下,表层土壤中的细粒物质得以保存和恢复,使土壤结构细化[7],进而增加土壤的持水性[32]。其次,生物结皮发育后降低了土壤容重,改善了土壤孔隙状况,进而间接影响土壤持水性[33]。结皮的发育程度对土壤持水性产生重要影响,本研究表明生物结皮持水性随生物结皮的演替呈先增加后降低的变化趋势。其主要原因可能与生物结皮层中的生物群落结构变化有关[34],藻结皮作为结皮演替的初期,以蓝藻为优势群落(如具鞘微鞘藻),具鞘微鞘藻有数十甚至上百条藻丝紧紧地缠绕,外面加上厚厚的胶鞘,并且鞘束之间相互交织,对砂粒起着明显的捆绑作用[34],同时分泌大量的EPS黏结砂粒,形成更稳定的团聚体,进而改善土壤孔隙条件[26-27],加之大气降尘填充在砂粒间隙,形成平滑致密的结皮,进而稳定土壤表面,增加土壤持水性和抗侵蚀能力[32,35]。当藻结皮发育至藻-藓混生结皮阶段时,藻类数量明显增多,丝状藻类主要集中在藻-藓混生结皮的上层,其与真菌菌丝共同起作用,对砂粒起着缠绕和捆绑作用,使结皮厚度和强度明显增加,进而提高土壤持水性[16,36]。当生物结皮演替为藓结皮,随着苔藓数量的增加,它们与藻类竞争土壤营养物质,致使藻类生物量下降[37],其中藻丝体和EPS含量低于前期发育阶段,致使其土壤持水性逐渐降低。此外,本研究中,随着生物结皮的演替,其导水和持水特性与土壤机械组成的相关性不显著,这说明不同发育阶段生物结皮土壤导水和持水性能的变化可能受其他因素(如生物结皮中非维管束植物和微生物的群落结构)的影响更大。
本研究表明,与裸沙相比,生物结皮形成后改善了表土理化性质,提高了表土持水能力,降低了土壤水分的下渗。从藻结皮-藻-藓混生结皮-藓结皮,其持水能力和阻碍水分下渗的能力均呈现先增加后下降的变化趋势,这可在一定程度上为当前生物结皮对土壤水分状况的影响提供参考。
4 结论
黄土高原生物结皮的发育显著增加了表土生物量、粉粒和黏粒含量,降低了砂粒含量和土壤容重,进而影响土壤导水和持水特性。与裸沙相比,藻结皮、藻-藓混生结皮和藓结皮的水分入渗参数分别平均降低了37.7%、54.3%和18.4%;田间持水量分别增加了0.97、1.10和0.70倍;整个退水过程(0~120 h)的平均土壤体积含水量分别增加了1.14、1.40和0.74倍。同时,藻-藓混生结皮对土壤导水和持水性能的影响大于藻结皮和藓结皮。与藻-藓混生结皮相比,藻结皮和藓结皮的水分入渗参数分别增加了36.5%和86.9%,田间持水量分别降低了3.0%和15.1%。即随着藻结皮向藓结皮发育演替,土壤导水性呈现先降低后增加的趋势,而土壤持水性表现为先增加后降低的变化规律。
猜你喜欢
新疆大学学报(自然科学版)(中英文)(2022年3期)2022-06-04 13:45:40
作物学报(2022年7期)2022-05-11 13:30:26
土壤(2021年5期)2021-11-17 06:19:44
水土保持通报(2020年6期)2020-03-10 01:46:52
新疆大学学报(自然科学版)(中英文)(2018年4期)2018-02-10 00:09:15
科学与财富(2017年26期)2017-09-23 09:56:05
纺织科技进展(2015年1期)2015-11-28 05:56:29
江西煤炭科技(2015年2期)2015-11-07 03:10:22
江苏农业科学(2014年9期)2014-11-15 14:11:31
果树学报(2013年2期)2013-05-07 03:14:06
