
PEDV E 蛋白对宿主细胞UPR 反应关键蛋白GRP78 表达的影响
2021-10-05 08:48王显赫徐梧皓张鑫林马大宇雷亚峰郑亮郭德轩吴志军张华曹宏伟
黑龙江八一农垦大学学报 2021年4期
王显赫,徐梧皓,张鑫林,马大宇,雷亚峰,郑亮,2,郭德轩,吴志军,2,3,张华,2,3,曹宏伟,2,3
(1.黑龙江八一农垦大学生命科学技术学院,大庆 163000;2.黑龙江八一农垦大学动物科技学院;3.黑龙江八一农垦大学生物技术中心)
猪流行性腹泻病毒(Porcine epidemic diarrhea virus,PEDV)是引起猪流行性腹泻(Porcine epidemic diarrhea,PED)的一种单股正链RNA 病毒,属于尼多病毒目(Nidovirales)冠状病毒科(Coronaviridae)α 冠状病毒属(Coronavirus)的成员之一[1]。该病对各年龄段的猪皆易感,并且能够引起仔猪的呕吐,腹泻,脱水而死,感染仔猪死亡率高达百分之九十[2]。虽然目前全球的科研工作者已经对PEDV 病毒有了较为广泛的认知与研究,但由于其RNA 病毒具有高变异性等特点,导致一直以来并没有特别有效的方法控制其增殖,任由其肆虐全球。
E 基因是PEDV 编码最小的一个结构蛋白基因,全长为231 bp,其编码蛋白大小约为7 kD,由76 个氨基酸组成,为II 型膜蛋白[3]。E 蛋白也是病毒粒子囊膜的组成成分之一,其C 端朝向病毒粒子的内部,N 端朝向病毒粒子的外部,并且包埋在脂质双分子层中[4]。与其他结构蛋白类似,E 蛋白也是一个多功能的蛋白,参与病毒囊膜的形成,在病毒复制和出芽过程中起重要作用[5]。
内质网(Endoplasmic reticulum,ER)是真核细胞的重要细胞器,主要负责蛋白质折叠和修饰、脂质生物合成和钙稳态[6-9]。当病原体入侵时,未折叠或错误折叠的蛋白质在内质网中积累,常常会导致内质网应激,内质网应激与宿主抗病毒天然免疫有着密切关系,内质网应激往往能够激活天然免疫信号通路来抑制病毒的复制[9-11]。在内质网应激下,能够激活一种叫做未折叠蛋白反应(Unfold protein response,UPR)的信号通路。UPR 通过减弱蛋白质合成、上调蛋白质折叠和转运机制以及激活ER 相关降解系统来恢复ER 稳态。当内质网应激时间过长且严重到内质网功能无法恢复时,UPR 会通过激活细胞凋亡来清除受损细胞[12-16]。研究以PEDV E 蛋白的真核表达载体感染宿主细胞,验证其正确表达和亚细胞定位,并通过利用免疫印迹技术检测UPR 反应的关键蛋白GRP78 表达水平,研究PEDV E 蛋白对内质网应激与未折叠蛋白反应的影响,为PEDV 与内质网应激之间的关系提供参考。
1 材料与方法
1.1 材料
1.1.1 细胞与质粒
人肾上皮细胞(HEK-293T)、猪肾上皮细胞(PK15)由实验室保存。细胞用Gibco 生物公司DMEM 培养基,加入10%胎牛血清以及100 μg·mL-1的青霉素与链霉素,在含5% CO2的37 ℃二氧化碳培养箱中培养。
pCMV-Myc 质粒和pCMV-Myc-E 质粒(PEDV E基因连接至pCMV-Myc 载体)由实验室构建保存。
1.1.2 抗体与试剂
鼠抗β-actin(TA-09)、山羊抗小鼠辣根酶标记IgG(ZB-2305)、山羊抗兔辣根酶标记IgG(ZB-2301)、鼠抗Myc 单克隆抗体(TA-01)、兔抗β-actin单克隆抗体(ZM-0001)、TRITC 标记的山羊抗小鼠IgG(ZF-0313)购自北京中杉金桥生物技术有限公司;兔抗GRP78 抗体(AF-0171)购自碧云天生物技术公司;转染试剂脂质体Lipo 2000(11668019)购自Invitrogen 公司;质粒小提试剂盒(D6943-02)和胶回收试剂盒(D2500-02)购自Omega 公司;毒胡萝卜素Thapsigargin(T9033)购自Sigma 公司。
1.2 实验方法
1.2.1 PEDV E 蛋白转染细胞
在转染质粒的前一天将HEK-293T 和PK15 细胞接种到24 孔板中,待细胞长满每孔的80%时,用PBS 缓冲液清洗细胞培养板,每孔加入400 μL 不含胎牛血清的DMEM 培养基,放入含5%CO2的37 ℃二氧化碳培养箱中培养30 min 。将对应的pCMV-Myc质粒和pCMV-Myc-E 质粒以800 ng 剂量与转染试剂脂质体Lipo 2000 混合,静置20 min 后,加入到相应的孔中分别培养12、24 h 和48 h 后收集细胞。
1.2.2 PEDV E 蛋白间接免疫荧光(IFA)鉴定
取出已经培养HEK-293T 和PK15 细胞24 h 后的细胞培养板,弃掉细胞培养液,用PBS 缓冲液清洗两次细胞培养孔,每孔500 μL 4%多聚甲醛固定细胞20 min,利用预冷PBS 清洗细胞孔3 次,每孔加入250 μL 浓度为0.1%的Triton X-100 溶液,透膜20 min,再次利用PBS 缓冲液清洗细胞孔3 次,每孔加入500 μL 现用现配的5%BSA 封闭2 h,再次利用PBS 缓冲液清洗细胞孔3 次,每孔加入250 μL 鼠抗Myc 单克隆抗体(1∶500)作为一抗室温孵育2 h,利用PBS 缓冲液清洗细胞孔3 次,每孔加入250 μL 的TRITC 标记的山羊抗小鼠IgG(1∶500)作为二抗室温避光孵育2 h,利用PBS 缓冲液清洗细胞孔3 次,DAPI 染色剂染色10 min 后用免疫荧光显微镜观察PEDV E 蛋白表达。
1.2.3 Western Blot 检测PEDV E 蛋白对内质网应激关键蛋白GRP78 的影响
将已经分别转染12、24 h 和48 h 后的细胞培养板取出,每孔用PBS 缓冲液清洗3 次,每孔加入35 μL 含1∶100 PMSF 的RIPA 裂解液在冰上裂解细胞30 min,用细胞刮刀将细胞刮下并且收集到1.5 ml EP 管中,4 ℃,12 000×g 离心10 min。取新的1.5 mL EP 管加入8.75 μL 的5×Loading Buffer,与上清混匀后沸水浴变性10 min,再次12 000×g 离心10 min。配置SDS-PAGE 凝胶,每孔上样10 μL,浓缩胶和分离胶分别用80 V 和120 V 电压进行电泳,对应Marker 剪下条带(GRP78:72 KDa;β-actin:42 KDa;Myc-E:15 KDa)。恒定电流0.2 A 转膜150 min,用5%脱脂奶粉封闭2 h。分别以兔抗GRP78 抗体(1∶1 000),鼠抗β-actin(1∶1 000)和鼠抗Myc(1∶1 000)为一抗孵育2 h,用TBST 缓冲液清洗8 次,每次15 min;山羊抗兔辣根酶标记IgG(1∶3 000)和山羊抗小鼠辣根酶标记IgG 为二抗孵育2 h,用TBST 缓冲液清洗8 次,每次15 min,最后曝光得出结果。进行Western Blot 检测PEDV E 蛋白时间和剂量梯度对UPR 反应关键蛋白GRP78 表达的影响。
2 结果与分析
2.1 PEDV E 蛋白Western Blot 检测表达情况
将pCMV-Myc-E 质粒和空白组分别转染HEK-293T 和PK15 细胞系24 h 后收样,通过Western Blot检测PEDV E 蛋白表达情况,结果显示,转染pCMVMyc-E 质粒的实验组检测到了PEDV E 蛋白的条带,而转染pCMV-Myc 质粒以及空白对照实验组没有检测到PEDV E 蛋白的表达(图1),以上结果说明pCMV-Myc-E 质粒能够在HEK-293T、PK15 细胞系内表达E 蛋白。
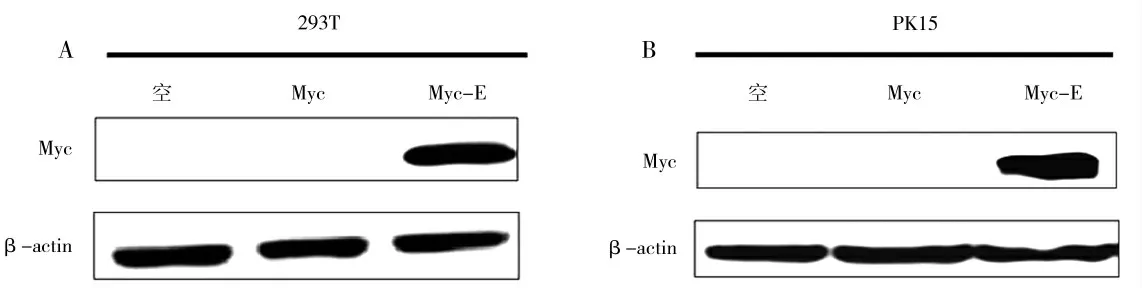
图1 Western Blot 检测证实PEDV E 蛋白表达Fig.1 Western Blot test confirmed PEDV E protein expression
2.2 PEDV E 蛋白间接免疫荧光(IFA)检测表达情况
利用pCMV-Myc-E 质粒分别转染HEK-293T、PK15 细胞系24 h 后收样,运用IFA 方法检测PEDVE 蛋白,结果显示,在HEK-293T 细胞系内,转染pCMV-Myc-E 质粒的实验组检测到了定位在细胞质上的PEDV E 蛋白,而转染pCMV-Myc 质粒以及空白对照实验组并没有检测到PEDV E 蛋白的表达,在PK15 细胞系内有同样的结果(图2),以上结果同样说明pCMV-Myc-E 质粒能够在HEK-293T、PK15 细胞系内表达E 蛋白。

图2 间接免疫荧光(IFA)检测PEDV E 蛋白表达Fig.2 Indirect immunofluorescence detection of PEDV E protein expression
2.3 PEDV E 蛋白能够内质网应激关键蛋白GRP78的上调
将pCMV-Myc-E 质粒分别转染HEK-293T、PK15 细胞系24 h 后收样,运用Western Blot 检测内质网应激关键蛋白GRP78 的变化,结果显示,转染pCMV-Myc-E 重组质粒实验组和毒胡萝卜素Thapsigargin(TG)处理组的GRP78 蛋白表达量与空白对照组相比有非常明显的增加,而转染pCMV-Myc 质粒实验组的GRP78 蛋白表达量与空白对照组相比则无明显差异(图3)。以上结果表明,PEDV E 蛋白转染HEK-293T、PK15 细胞能够引起GRP78 蛋白表达量的上调,进而引起UPR 反应和内质网应激。而pCMV-Myc 质粒转染HEK-293T、PK15 细胞并不能够引起GRP78 蛋白表达量的上调。
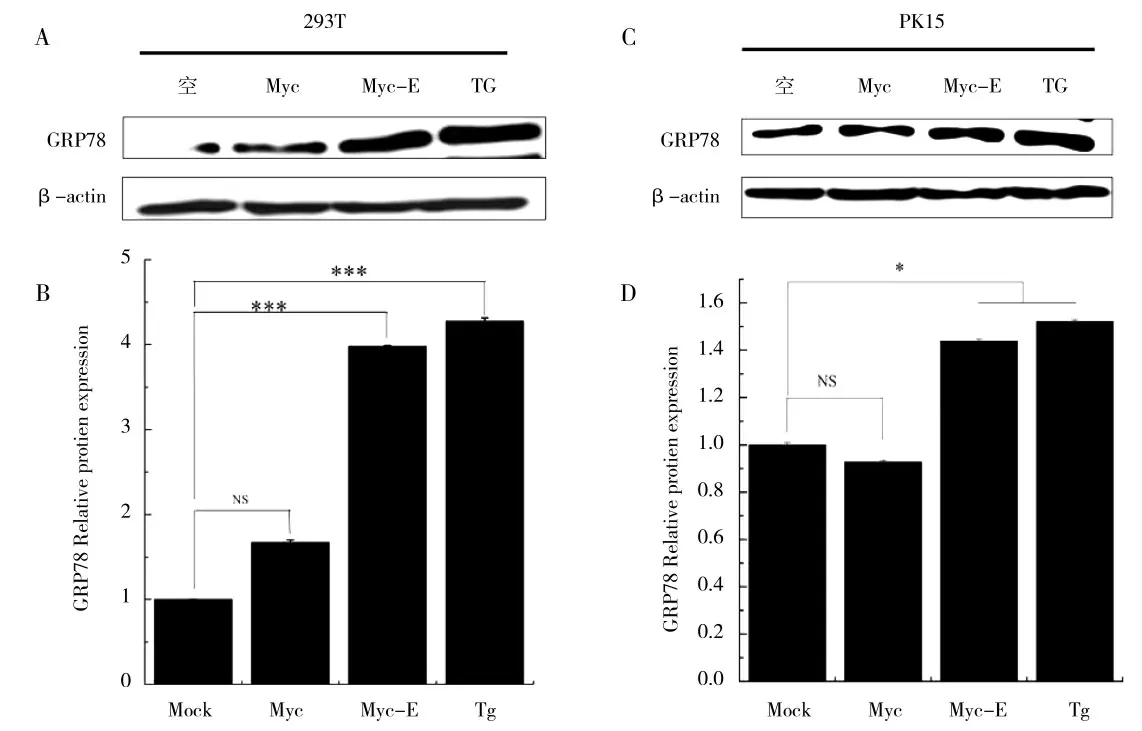
图3 PEDV E 蛋白能够引起UPR 关键蛋白GRP78 的上调Fig.3 PEDV E protein can cause the up-regulation of UPR key protein GRP78
2.4 PEDV E 蛋白剂量梯度引起UPR 关键蛋白GRP78 表达情况
利用pCMV-Myc-E 质粒以及pCMV-Myc 质粒以剂量梯度(800、1 200、1 600 ng)分别转染HEK-293T、PK15 细胞24 h 后收样。Western Blot 检测结果显示,UPR 关键蛋白GRP78 蛋白表达量随着转染pCMVMyc-E 质粒浓度的升高而表达量逐渐上调,而PEDV E 蛋白的表达量也逐渐增加;而GRP78 蛋白的表达量并不随着pCMV-Myc 质粒转染浓度的提升而增加(图4),以上结果表明,随着PEDV E 蛋白转染剂量的增多,GRP78 蛋白的表达量也随之增加,UPR 反应逐渐加剧,而随着pCMV-Myc 质粒转染剂量的增加,GRP78 蛋白的表达量无明显变化,这进一步说明pCMV-Myc 不能够引起内质网应激。
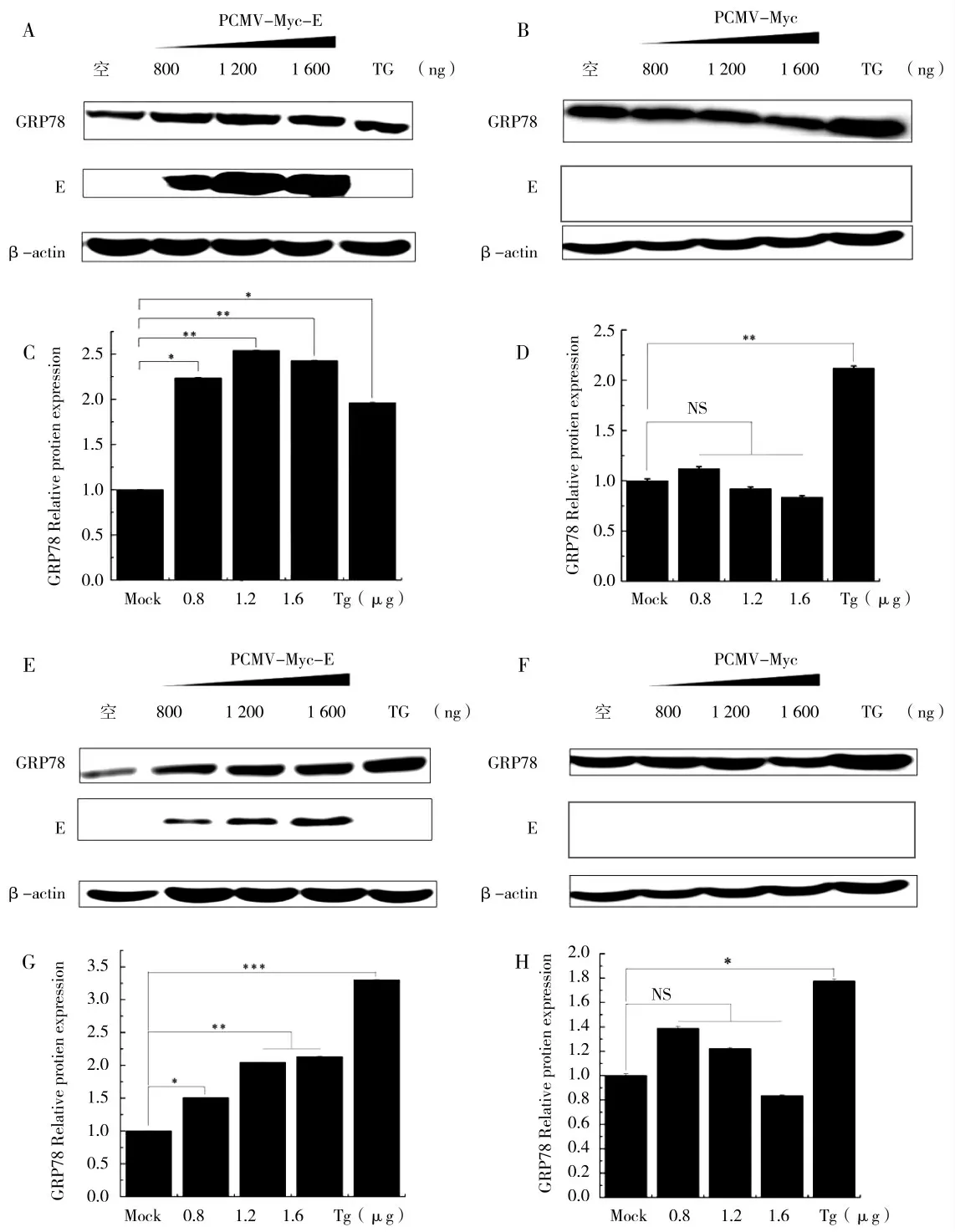
图4 PEDV E 蛋白剂量梯度引起UPR 关键蛋白GRP78 表达上调Fig.4 PEDV E protein dose gradient causes up-regulation of UPR key protein GRP78 expression
2.5 PEDV E 蛋白时间梯度转染引起UPR 关键蛋白GRP78 表达情况
利用pCMV-Myc-E 质粒以及pCMV-Myc 质粒以时间梯度(12、24、48 h),800 ng 分别转染HEK-293T、PK15 细胞收样。Western Blot 检测结果显示,内质网应激关键蛋白GRP78 蛋白表达量随着转染pCMVMyc-E 质粒转染时间的延长而表达量逐渐上调,而PEDV E 蛋白的表达量也逐渐增加;而GRP78 蛋白的表达量并不随着pCMV-Myc 质粒转染时间的延长而增加(图5),以上结果表明,随着PEDV E 蛋白转染时间的增加,GRP78 蛋白的表达量也随之增加,内质网应激反应逐渐加剧,而随着pCMV-Myc 质粒转染时间的增加,GRP78 蛋白的表达量无明显变化,这说明pCMV-Myc 不能够引起内质网应激。
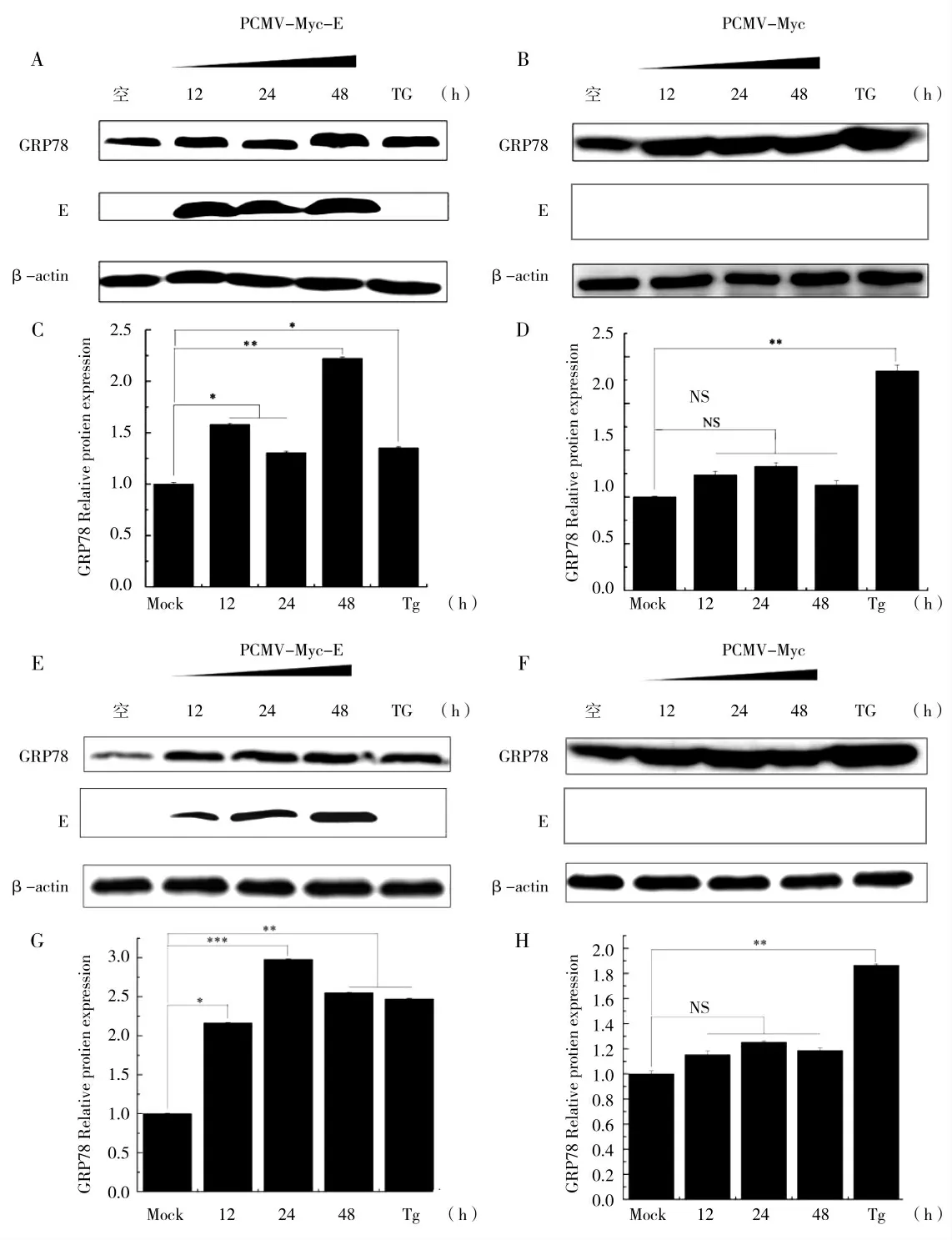
图5 PEDV E 蛋白时间梯度引起内质网应激关键蛋白GRP78 表达上调Fig.5 The time gradient of PEDV E protein causes up-regulation of the expression of GRP78,a key protein of endoplasmic reticulum stress
3 讨论
内质网是真核细胞内合成与运输蛋白质重要的细胞器,当外界环境发生变化,如:紫外灭活,与葡萄糖剥夺、缺氧、异常Ca2+的调节,以及病原体的入侵等等,内质网腔内会累积大量的未折叠或错误折叠蛋白质。这些未折叠蛋白质能够引起结合在未折叠蛋白反应(UPR)效应器上的葡萄糖调节蛋白78(GRP78)解离并且与未折叠蛋白质结合,激活UPR反应,进而激活内质网应激。目前已有研究发现多种病毒感染能够激活UPR 反应,如:TGEV 病毒感染细胞会激活全部的三种UPR 信号通路,并且PERKeIF2α 信号通路可以负调控TGEV 病毒的复制[17];陈全钢的研究发现猪繁殖与呼吸综合征病毒感染MARC-145 细胞能够激活PERK 和IRE-1 信号通路,但不激活ATF6 信号通路[18];肠道病毒EV71 感染细胞能够激活IRE1-XBP1 和PERK 信号通路,而且抑制磷酸化的IRE1 时,病毒的复制能力增强,抑制IRE1 核酸内切酶活性会导致EV71 病毒复制能力的减弱[19]。
PEDV 自2010 年在我国大规模爆发以来,由于其对仔猪高致死率以及高突变性,给我国养猪业带来了巨大的经济损失。而E 蛋白又是构成PEDV 的结构蛋白之一,能够参与病毒囊膜的形成并且在病毒复制与出芽过程中发挥重要作用[20]。因此,研究PEDV E 蛋白对内质网的刺激以及内质网应激下游分子调控PEDV 的复制就显得尤为重要。研究发现,PEDV E 蛋白能够引起内质网应激标志性蛋白GRP78 的增加,说明PEDV E 蛋白能够引起内质网应激反应,由于PEDV E 蛋白定位在细胞质上,当PEDV E 蛋白表达量随着转染剂量和转染时间的增加而增加,导致内质网中未折叠或者错误折叠蛋白数量上升,GRP78 蛋白与UPR 反应效应器分离,激活了UPR 反应,但是具体是激活UPR 反应的哪几条信号通路,对内质网应激下游分子的调控还需后续研究证明,以其为研制出PEDV 与宿主互作提供理论基础。
猜你喜欢
科技视界(2022年10期)2022-05-20
中国药房(2022年9期)2022-05-17
科学与生活(2021年16期)2021-11-25
青岛大学学报(医学版)(2021年5期)2021-11-17
商品与质量(2021年31期)2021-08-20
昆明医科大学报(2019年2期)2019-09-10
科教导刊·电子版(2018年25期)2018-10-31
新课程·中学(2018年10期)2018-02-19
中国医学创新(2017年15期)2017-06-12
中国实用医药(2009年5期)2009-05-08
