
草地早熟禾NAC基因鉴定及非生物胁迫下表达模式分析
2021-09-25 03:11朱瑞婷牛奎举张然马晖玲
草原与草坪 2021年4期
朱瑞婷,牛奎举,张然,马晖玲
(甘肃农业大学草业学院,草业生态系统教育部重点实验室,甘肃省草业工程实验室,中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
生存环境对植物生长具有重要影响[1],其中,干旱、高盐、低温、高温、重金属等是抑制植物生长发育的主要非生物胁迫因素。干旱胁迫会限制植物生长,改变植物渗透平衡[2]。重金属会破坏植物细胞中的蛋白质结构,产生有害物质[3]。盐胁迫会破坏质膜的完整性从而抑制植物生长[4]。高温胁迫会影响植物的生理代谢,对植物生长造成一定的损害[5]。在长期演变过程中,植物自身形成了一套复杂的防御系统来抵御不良环境的伤害,在这些防御方法中,转录因子家族在植物响应外界环境胁迫的转录调控中起到了关键作用[6]。
转录因子是一类通过与胁迫相关基因的特异顺式作用元件相结合从而激活或抑制靶基因转录的DNA结合蛋白,参与植物在非生物胁迫下的响应[7]。NAC家族是植物特有的一类转录因子,具有不同的生物学功能。NAC家族在N端具有保守的NAC结构域,大约有150个氨基酸,C端为高度变异的转录激活区[8]。其功能涉及植物生长发育、激素调控和胁迫响应等重要方面[9]。目前在很多植物中已经鉴定出大量NAC家族成员,如石榴(Punicagranatum)[10]、果梅(Prunusmume)[11]、苦荞(Tartarybuckwheat)[12]、西瓜(Citrulluslanatus)[13]等。不同物种间家族成员的数量与功能存在重要差异,但大量的NAC转录因子在非生物胁迫反应中起到调控作用[14-17]。如玉米(Zeamays)ZmNAC84在烟草中的过表达可增强耐旱性[18],过表达OsNAC5可以增强水稻(Oryzasativa)的耐盐性[19],低温胁迫下番茄(Solanumlycopersicum)SINAM1的过表达减轻了番茄在遭受低温胁迫后的氧化损伤[20]。
草地早熟禾(Poapratensis)是禾本科早熟禾属植物,广泛分布于北温带冷湿润地区,是一种多年生冷季型禾草,叶形美观,具根状茎,是优良的草坪草。草地早熟禾喜冷凉湿润的气候,耐寒性较强,但抗旱性较差[21]。随着全球性气候、生态和环境问题日益严重,尤其是全球变暖的加剧,干旱、高盐、高温、重金属污染已经成为制约农业发展的主要因素[22],提高草坪草的抗逆性是目前草坪研究中的热点问题。草坪草抗逆基因的鉴定对了解其响应逆境胁迫的分子机制和抗逆新品种的选育具有重要意义。有关非生物胁迫下草坪草NAC基因表达模式分析的报道较少,草地早熟禾NAC基因尚未被完全鉴定。本课题组前期进行了草地早熟禾抗旱、抗寒、耐镉转录组分析,发现NAC基因家族基因参与草地早熟禾的逆境胁迫响应。本研究以前期组装的草地早熟禾转录组数据库为基础,对具有全长cDNA的草地早熟禾NAC基因进行鉴定并分析其不同胁迫下的表达模式,为草地早熟禾NAC基因家族的分子调控机理研究提供理论依据,有助于优良基因资源的挖掘和新品种培育。
1 材料和方法
1.1 供试材料与生长条件
试验材料为草地早熟禾品种巴润,由北京克劳沃公司提供。采用砂培法种植,将清洗干净的沙子等量的装入直径为9 cm、高度为17 cm的育苗钵中,种子直接播种后于生长室中培养。培养条件为:白天温度为25℃,时间为16 h,光照强度为6 000 lx,夜晚温度为20℃,时间为8 h,湿度为50%。播种20 d后,用Hoagland营养液培育,持续培养4周,在此期间每隔5 d换1次营养液和蒸馏水。
1.2 植物处理
选择长势一致的幼苗,对其进行6组处理,每组处理4个重复,第1组为对照:正常营养液栽培;第2组为盐处理:200 mmol/L NaCl溶液栽培;第3组为干旱处理:15% PEG-6000溶液栽培;第4组为低温处理:0℃培养;第5组为高温处理:38℃培养;第6组为重金属处理:3 mmol/L CdCl2溶液栽培。处理时间6、24、48 h、7 d,每次取样4次重复,取其叶片,迅速放入液氮中冷冻后保存于-80℃。
1.3 全长草地早熟禾NAC基因的鉴定
二穗短柄草(Brachypodiumdistachyon)NAC蛋白序列可根据文献提供的登录号在NCBI上搜索获得(https://www.ncbi.nlm.nih.gov/)[23]。在BioEdit软件中,将本实验室已有和报道的5个草地早熟禾转录组数据库作为本地数据库[24-28],以二穗短柄草NAC蛋白序列作为请求序列,Program选择:‘tblastn’,Expectation Value选择E-20,搜索本地草地早熟禾数据库。将所得到的序列通过NCBI网站中的blastX进行搜索比对,获得候选序列。
1.4 序列分析及构建系统进化树
通过ExPASy在线网站上的Prat Param工具对草地早熟禾NAC家族基因的理化性质进行初步预测,包括基因的大小、分子量、等电点、不稳定指数、亲水性的平均值和脂肪指数。通过MEME网站(http://meme-suite.org/tools/meme)对草地早熟禾NAC基因的保守基序进行分析。根据在线网站(https://www.ncbi.nlm.nih.gov/orffinder)预测草地早熟禾NAC基因的编码蛋白序列,采用邻近法在MEGA 7.0软件上对PpNACs进行聚类分析。
1.5 荧光定量PCR(qRT-PCR)
RNA提取使用天根公司植物RNA提取试剂盒,RNA反转录使用Prime Script RT reagent Kit with gDNA Eraser(Perfect Real Time)试剂盒,第1步去除基因组DNA反应,第2步为反转录合成cDNA,稀释20倍作为模板。使用NCBI在线网站Prime-BLAST( https://www.ncbi.nlm.nih.gov/tools/primer-blast/)进行引物设计(表1)。利用天根荧光定量试剂盒进行qRT-PCR分析,20 μL反应体系如下:模板5 μL,引物2 μL,ddH2O 3 μL,2×SuperReal PreMix Plus 10 μL。荧光定量PCR的条件为:95℃预变性15 min,40个循环的PCR扩增,包括95℃变性10 s和58℃退火30 s。内参基因为ACT,每个样品设3次技术重复,基因的相对表达量使用2-ΔΔCT法计算[29]。

表1 定量PCR引物
1.6 数据分析
采用SPSS 23.0软件进行T检验,并用GraphPad Prism 8软件绘图。
2 结果与分析
2.1 草地早熟禾NAC基因家族的生物信息学分析
2.1.1 草地早熟禾NAC转录因子的鉴定 根据二穗短柄草的NAC蛋白序列,在BioEdit软件中搜索本地草地早熟禾数据库,通过搜索比对得到15个草地早熟禾NAC全长cDNA序列,具有完整的ORF序列,根据草地早熟禾拉丁名将其命名为PpNACs(表2)。结果表明,15个草地早熟禾NAC基因中分子量在61 184.22(PpNAC18)~163 713.04(PpNAC20),等电点在4.89~5.04,都属于酸性蛋白;不稳定系数除了PpNAC16外都大于40,属于不稳定蛋白;亲水性系数在0.705~1.022,属于疏水性蛋白(大于零表示疏水性蛋白,小于零表示亲水性蛋白,在±0.5之间为两性蛋白)。
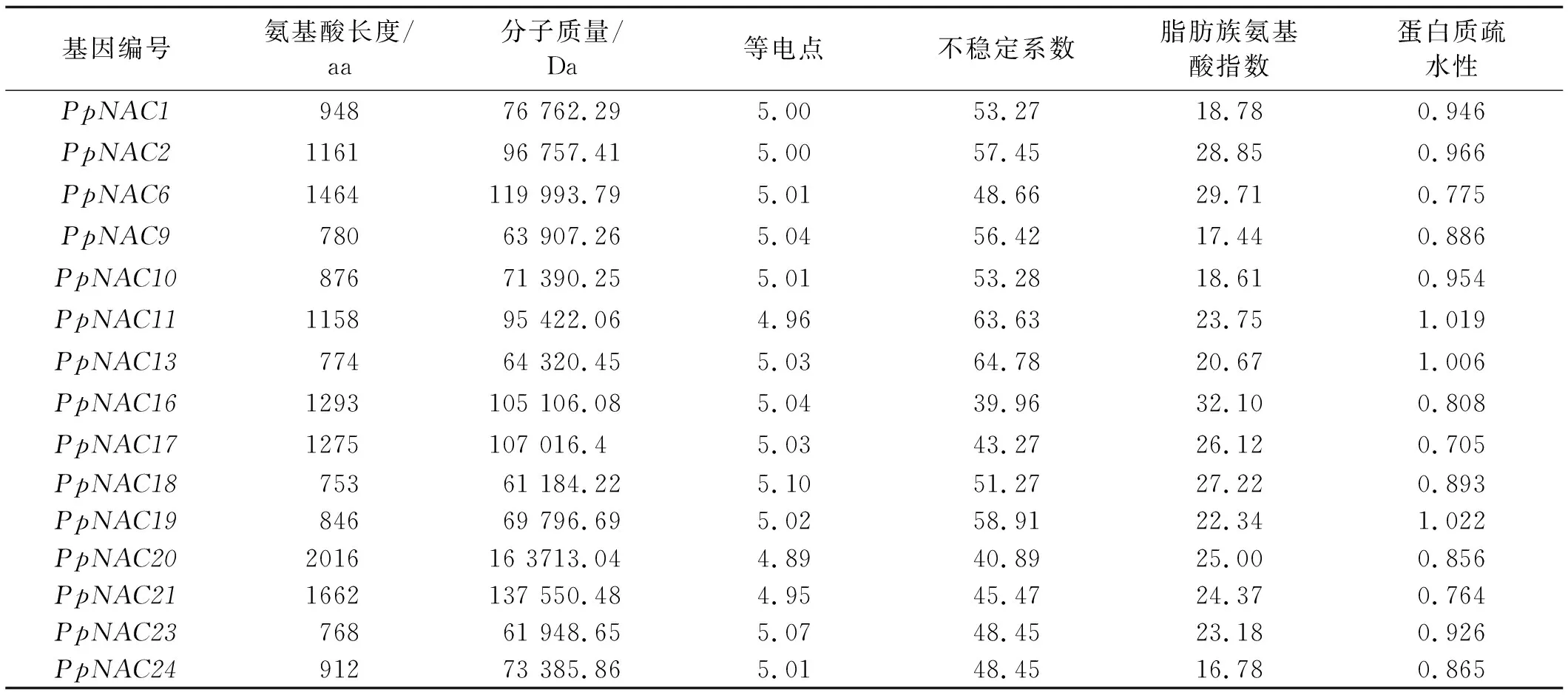
表2 15个全长草地早熟禾NAC转录因子理化性质分析
2.1.2 PpNACs基因的motif分析和聚类分析 采用MEME网站在线预测草地早熟禾NAC基因的保守结构域,可知15个NAC家族基因一共由12种motif组成(图1)。其中,motif 2和motif 7在所有草地早熟禾NAC基因中都存在。草地早熟禾NAC基因由多种motif组成,除PpNAC16、PpNAC18、PpNAC21和PpNAC23外都具有motif 1-7。Motif 1在80%的PpNACs中存在,Motif 2和motif 6在87%PpNACs中存在。
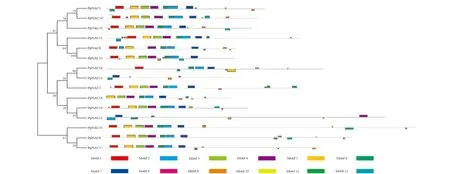
图1 草地早熟禾NAC基因的基序结构和聚类分析Fig.1 Motif composition and cluster analysis of NAC genes in Poa pratensis
对15个NAC家族基因进行聚类分析,可分为3大类,第1类包括PpNAC1、PpNAC9、PpNAC10、PpNAC11、PpNAC13、PpNAC24,都含有motif 1-7;第2类包括PpNAC2、PpNAC16、PpNAC18、PpNAC19、PpNAC21、PpNAC23,都含有motif2和motif7;第3类包括PpNAC6、PpNAC17和PpNAC20,都含有motif 1-7、motif 9和motif 11。
2.2 草地早熟禾NAC基因家族表达模式分析
2.2.1 重金属胁迫下NAC基因的表达PpNAC9和PpNAC16随着时间的增大,呈现逐渐上升的趋势,在7 d时达到最大值,分别是对照的6.53倍和8.27倍;PpNAC1、PpNAC2在6 h时表达量显著增大,随后呈现下降又上升的趋势;PpNAC23在48 h时表达量显著增大,在其他时间段无显著性变化;PpNAC10和PpNAC18在7 d时表达量达到最大,分别是对照的223.33倍和99.72倍;PpNAC1、PpNAC2、PpNAC10、PpNAC11、PpNAC13、PpNAC16、PpNAC17、PpNAC19、PpNAC24在4个时间段内均有显著性变化(图2)。
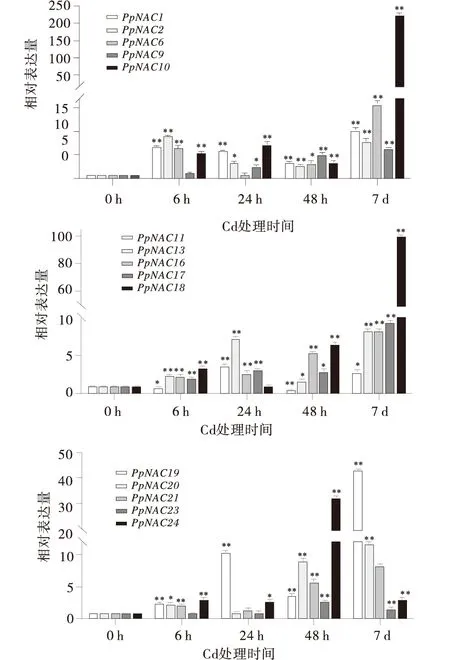
图2 重金属胁迫下NAC基因的表达分析Fig.2 Relative expression level of NAC genes under heavy metal treatment注:竖线表示标准误,*表示差异显著(P<0.05),**表示差异极显著(P<0.01)。下同
2.2.2 盐胁迫下NAC基因的表达PpNAC9和PpNAC13在四个时间段内都有显著性变化,PpNAC9、PpNAC10、PpNAC11、PpNAC16、PpNAC18、PpNAC19、PpNAC20、PpNAC23在7 d时表达量达到最大值,PpNAC10在7 d时表达量是对照的50.88倍,PpNAC24在6 h时表达量显著增大,在48 h时表达量达到最大,是对照的36.33倍。PpNAC1、PpNAC13、PpNAC17在6 h是表达量显著性增大,随后呈现先下降后上升的趋势,响应短期胁迫和长期胁迫(图3)。

图3 盐胁迫下NAC基因的表达分析Fig.3 Relative expression level of NAC genes under salt treatment
2.2.3 干旱胁迫下NAC基因的表达PpNAC9、PpNAC11、PpNAC17在干旱胁迫48 h内变化不大,在7 d时表达量有明显的增大,响应长期干旱胁迫。PpNAC1和PpNAC13在0~48 h内呈现先上升后下降趋势,在48 h~7 d呈现上升趋势且在7 d时表达量达到最大,分别是对照的94.08倍和80.30倍。在干旱胁迫下PpNAC2和PpNAC24在6 h时表达量显著增大,达到最大值,而其他基因在7 d时表达量达到最大值(图4)。

图4 干旱胁迫下NAC基因的表达分析Fig.4 Relative expression level of NAC genes under drought treatment
2.2.4 低温胁迫下NAC基因的表达 在低温胁迫下,PpNAC2和PpNAC13在6 h表达量显著增大,24 h持续增大达到最大值,48 h呈现下降趋势,表达水平受到抑制,7 d又出现上升趋势。PpNAC1和PpNAC10的表达量呈现先上升后下降的趋势,在24 h达到峰值,分别是对照的66.38倍、98.46倍。PpNAC17的表达量随时间的增大表现出上升趋势。PpNAC21的表达量与对照相比在短期处理时表现出显著增大,在长期处理时表现出显著减小。低温胁迫下PpNAC1、PpNAC6、PpNAC10、PpNAC13、PpNAC16、PpNAC17的表达量均高于对照(图5)。

图5 低温胁迫下NAC基因的表达分析Fig.5 Relative expression level of NAC genes under low temperature treatment
2.2.5 高温胁迫下NAC基因的表达 研究表明,该家族大部分成员在高温7 d时的表达水平较高。在高温胁迫下,PpNAC16的表达量均具有显著性变化,表达水平上调。随着高温处理时间的增大,PpNAC17、PpNAC19和PpNAC20的表达持续上调,在7 d时处于较高的表达水平,分别是对照的5.85倍、5.35倍和11.63倍(图6)。

图6 高温胁迫下NAC基因的表达分析Fig.6 Relative expression level ofNAC genes under high temperature treatment
3 讨论
在其他高等植物如拟南芥(Arabidopsisthaliana)、水稻中,NAC转录因子的功能已被阐明,具有相似结构域的蛋白质具有相同或相似的生物学功能[30],已有文献报道,构建进化树预测其功能的方法可应用于其他物种[21],因此,通过对草地早熟禾和二穗短柄草NAC基因家族的系统进化树分析,有助于预测同一亚族的PpNACs基因的功能。已知的NAC基因家族成员参与植物的生长发育和不同胁迫反应,揭示了NAC基因的序列多样化导致功能的多样化[31],为草地早熟禾NAC基因家族的鉴定提供了坚实的基础。草地早熟禾可用于草坪草的种植和提高城市绿化覆盖率,但耐寒性强,抗旱性较差。NAC基因家族具有高度保守的结构域,对植物的非生物胁迫应答过程具有重要的作用。本试验从草地早熟禾转录组数据库中鉴定出15个全长cDNA 序列的NAC基因。PpNACs家族成员的等位点在4.89~5.04,与李伟[32]克隆所得PpNACs基因的等位点为5.30相一致,都属于酸性蛋白;与二穗短柄草和水稻NAC基因家族成员数目相差较大,可能是在进化过程中出现了丢失,也可能是由于转录组数据库有限而导致NAC基因家族成员的不完整性。根据NAC基因的保守结构域分析发现,PpNACs家族基因在N端含有基本的NAC结构域,NAC结构包括N端的DNA结合区和C端的转录激活区,N端大约有150个氨基酸残基,分为A~E 5个亚结构域, A、C、D结构域为主要的亲水区,E结构域为主要的疏水区,在PpNACs转录因子家族成员中都含有E结构域,都属于疏水性蛋白。
在重金属胁迫下,PpNACs基因随着胁迫时间的延长呈现不同的趋势,而在此胁迫过程中PpNAC10、PpNAC18、PpNAC19和PpNAC24为高表达。研究表明,水稻中NAC基因参与铜胁迫过程[33],在西瓜中NAC基因家族在缓解镉胁迫中可能具有重要作用[34],关于NAC基因家族在重金属胁迫中的研究很少,初步推测PpNAC10、PpNAC18、PpNAC19和PpNAC24在镉胁迫过程中起到重要作用。PpNACs基因在盐胁迫下大部分呈现上升下降再上升的趋势,而且在第7 d时都表现为上调,PpNAC10和PpNAC24在盐胁迫过程中表现出高表达量,出现的峰值时间不同,分别在7 d和48 h。研究表明,过表达ONAC045可提高水稻的耐盐性[35],在拟南芥中过表达CiNAC3表现出更强的耐盐性[36]。因此,PpNAC10和PpNAC24基因可作为研究草地早熟禾耐盐性的候选基因。在干旱胁迫下,所有PpNACs基因在不同的时间响应程度不同,表达量均有不同程度的增加,在第7 d时表达量具有显著差异性(除PpNAC24外),均高于对照。PpNAC1、PpNAC6、PpNAC10、PpNAC13基因在7 d表现出高表达量,而在拟南芥中超量表达NAC基因可提高其抗旱性[37]。在干旱胁迫下水稻OsNAC095基因的表达量增加,通过减少体内水分的丧失提高水稻的抗旱性[38]。同样在本试验干旱胁迫下所有基因的表达量都有所增加,因此推测这些基因参与草地早熟禾对干旱胁迫的调控,PpNAC1、PpNAC6、PpNAC10、PpNAC13可能在抗旱性方面起到重要作用。
在低温胁迫下,PpNACs基因具有不同的表达趋势,有的基因在胁迫过程中表现为全部上调,如PpNAC1、PpNAC6、PpNAC10、PpNAC16和PpNAC17;有的基因在胁迫过程中的表达趋势为先上升后下降再上升,如PpNAC2和PpNAC13;有的基因则表现为先上升后下降再上升又下降的趋势,如PpNAC6、PpNAC9、PpNAC16、PpNAC21和PpNAC24。在高温胁迫下,除PpNAC20外的其他基因表达量的变化在10倍以内。PpNACs基因表达量的变化趋势具有多样性,如PpNAC1、PpNAC16和PpNAC20在胁迫过程中都表现为上调;PpNAC16、PpNAC23和PpNAC24在胁迫过程中为上升下降上升下降趋势。由此发现在高低温胁迫下PpNAC1和PpNAC16都表现为上调,PpNAC24都呈现“M”趋势,它们在温度胁迫下具有相同的变化趋势,可能是受到相同的分子调控,参与相同的调控路径。在茶树(Camelliasinensis)中CsNAC1和CsNAC2响应温度胁迫并有复杂的调控机制[39]。本试验表明PpNACs基因响应高温和低温胁迫,不同基因响应高低温胁迫程度不同,体现了PpNACs基因在温度胁迫下具有复杂的调控性。研究发现,低温胁迫下番茄中SlNAM1基因的过表达会提高渗透物的含量,降低超氧阴离子的含量,提高植株的抗寒性[20]。PpNAC1和PpNAC10在24 h表现为高表达量,PpNAC18在48 h表现为高表达量,它们可以作为草地早熟禾对低温胁迫下调控的候选基因。研究发现在番茄中抑制SlNAC1的表达影响热激蛋白的积累,转基因植株的抗性降低,SlNAC1在番茄耐高温中起到正调控作用[40],本研究中PpNAC20在高温胁迫下为高表达,它可能在耐高温过程中起到重要作用。
在本试验中同时分析了PpNACs基因在重金属、高盐、干旱、高温和低温胁迫下的表达情况,15个PpNACs基因具有不同的表达模式,这些基因参与多种信号传导途径,为确定基因功能提供了重要信息[41-42]。试验发现PpNAC10在重金属、高盐、干旱和低温胁迫下为高表达,PpNAC18在重金属、干旱和低温胁迫下为高表达,PpNAC10和PpNAC18在多种非生物胁迫下表现突出,因此可将PpNAC10和PpNAC18作为候选基因进行功能验证。
4 结论
根据二穗短柄草的蛋白序列和草地早熟禾转录组数据,通过生物信息学分析得到15个PpNACs基因全长cDNA序列,具有完整的开放阅读框。15个PpNACs转录因子都是酸性蛋白,除了PpNAC16,其他都属于不稳定蛋白,疏水性蛋白。PpNACs成员通过聚类分析可分为3类,Motif分析发现大部分PpNACs基因家族成员具有motif 1、2、3、4、5、6、7,具有相似的结构。通过qRT-PCR分析草地早熟禾在不同非生物胁迫下的表达水平发现,15个PpNACs基因均能响应不同的非生物胁迫,具有不同的表达模式,与重金属胁迫相关的基因有PpNAC10、PpNAC18、PpNAC19和PpNAC24;与耐盐性相关的基因有PpNAC10和PpNAC24;与抗旱性相关的基因有PpNAC1、PpNAC6、PpNAC10和PpNAC13;与耐寒性相关的基因有PpNAC1、PpNAC10和PpNAC18;与耐高温相关的基因有PpNAC20,综上发现PpNAC10和PpNAC18在多种抗逆过程中起到重要作用,可作为后续功能验证试验的候选基因。
猜你喜欢
幼儿100(2020年31期)2020-11-18
小哥白尼(军事科学)(2019年9期)2019-12-21
疯狂英语·初中版(2019年4期)2019-09-10
广州大学学报(自然科学版)(2019年1期)2019-05-07
电影(2019年3期)2019-04-04
小太阳画报(2018年6期)2018-05-14
少儿科学周刊·少年版(2017年3期)2017-06-29
天津科技大学学报(2016年1期)2016-02-28
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10
现代检验医学杂志(2015年2期)2015-02-06
