
雅砻江官地水库环境因子和浮游植物群落结构的时空变化及相互关系
2021-09-25 02:22李天才甘维熊姚维志苏胜齐
西南农业学报 2021年8期
李 锋,李天才,甘维熊,姚维志,付 梅,熊 波,苏胜齐*
(1.西南大学水产学院,重庆 400700;2.西南大学渔业资源环境研究中心,重庆 400700;3.雅砻江流域水电开发有限公司,四川 成都 610000)
【研究意义】雅砻江官地水库建于2013年,处于西昌市与盐源县交界处,上游连接雅砻江锦屏大河湾,下游毗邻二滩水库[1],是雅砻江下游由人烟稀少地区转入人口密集城镇的枢纽,也是长江上游重要生态屏障和水源涵养地的组成部分。因此,掌握官地水库水质状况和浮游植物群落结构可为管理、保护和改善区域水生态环境提供基础资料和科学依据,有助于下游作为饮用水源的二滩水库水质变化分析和提前管理。【前人研究进展】水体理化指标和营养成分含量是评判水库水质等级的直接数据来源。浮游植物是水库生态系统中重要初级生产者,是水库生态系统食物网的基础,其密度、结构直接影响高级捕食者的种群结构,继而对整个生态系统产生重要影响[2-3]。藻类多数为单细胞生物,对环境因子的变化极为敏感,故而其种类组成、密度分布、多样性及特殊种类可反映水体环境因子及污染状况,是评价水库生态系统的重要指标[4-6]。研究[7-8]表明,总体上湖泊型水库污染程度高于河道型水库,下游水库又高于上游水库;养殖型、灌溉型水库污染程度高于发电型水库,临近城镇或工业园区的水库污染程度高于山区源头水库。关于环境因子和浮游植物的调查多数是因为水库已出现不同程度的污染,前人研究目的大多为掌握水库基本情况,提出针对性治理方案[7-10]。【本研究切入点】官地水库地处偏僻,属于山区源头发电-河道型水库[11],多年观察初步判断其水质良好。而本研究参照成熟的水库环境因子和浮游植物调查方法,历经1年4个季度的调查,旨在系统掌握官地水库水质状况和浮游植物群落结构。【拟解决的关键问题】分析存在的官地水库环境因子和浮游植物群落的时空变化特征,并梳理相互之间的关系,为保护水质、预防恶化提供科学管理依据。
1 材料与方法
1.1 采样时间与断面
官地库区回水长度近60 km,库区河道狭窄,并无大的支流汇入,因此仅在干流主要位置设置采样断面即可。经现场勘察、分析,沿江自上游到下游设置A/B/C 3个采样断面(表1和图1)。每个断面采样时采集左右两岸水体和浮游植物样本,以两岸平均数作为调查结果。分别于2018年冬季(12月)和2019春季(3月)、夏季(7月)、秋季(10月)完成官地库区周年水质及浮游植物采样调查。
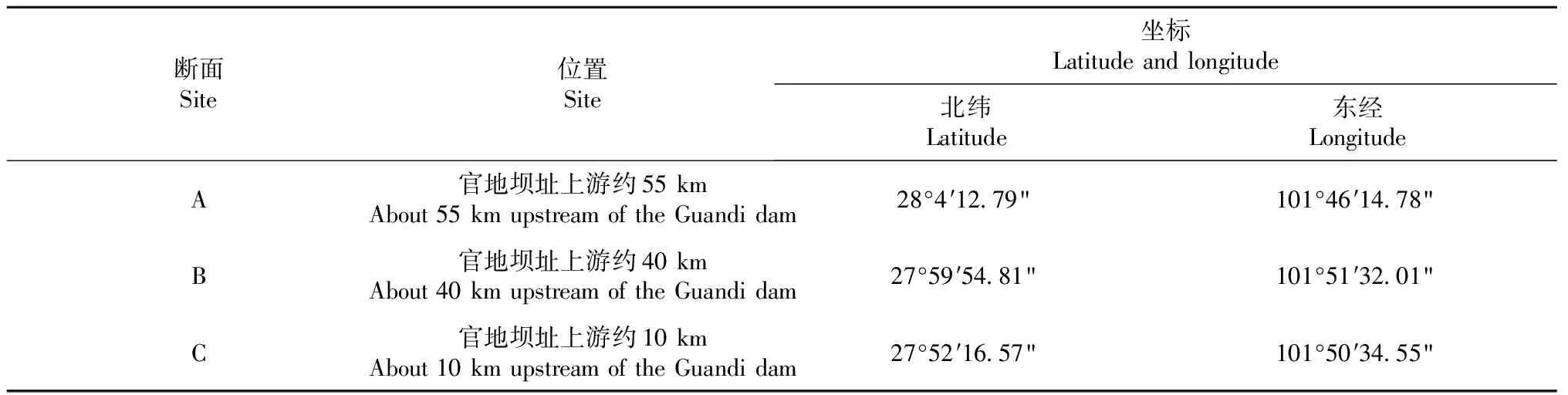
表1 官地库区采样断面设置
1.2 水样采集和环境因子测定
水体理化指标测定按照《水环境检测规范》(SL 219-98)和《水库渔业资源调查规范》(SL 167-96)进行。水温(WaterTemperature, WT)和溶解氧(Dissolved Oxygen,DO)使用YSI55型溶氧仪现场测定;pH值使用便携式pH计现场测定;电导率(Conductivity, CT)使用DOB33A型电导仪现场测定;透明度(Transparency, SD)使用塞氏盘直接现场测定。水体硬度(Hardness of Groundwater,HG)、化学需氧量(Chemical Oxygen Demand,COD)、总氮(Total Nitrogen, TN)、总磷(Total Phosphorus, TP)等环境因子参照《水和废水监测分析方法(第四版)》[12]在西南大学渔业资源与环境中心实验室进行测定。
1.3 浮游植物样本采集和分析
用2 L有机玻璃采水器分别定量表层、30和50 cm水层水样,均匀混合后取1 L水体作为浮游植物分析样本。浮游植物水样加15 mL 1.5%(V/V)鲁哥(Lugol)氏液进行固定后带回实验室进行分析。浮游植物水样用浮游生物沉淀器在实验室沉淀24 h后吸出上清液,剩余20~50 mL浓缩水样倒入浮游生物标本瓶中以待定量鉴定。定量鉴定时将浓缩水样摇匀,取0.1 mL置于计数框内,在10×40倍镜下观察、鉴定、计数浮游植物。每瓶样本计数取两次观测的平均值,同一样本的两次定量鉴定值不大于平均值的5%为有效观测,否则继续观察直至符合要求。浮游植物种类鉴定主要参照《中国淡水藻类》[13]、《中国内陆水域常见藻类图谱》[14]、《中国常见淡水浮游藻类图谱》[15]等资料。
1.4 数据处理
采用 Excel 2016对水体环境因子和浮游植物群落结构进行汇总分析、作图;采用SPSS 22.0进行Pearson相关性分析;采用Conaco 4.5进行聚类、冗余分析并作图。
ρ=10nv
式中,ρ为浮游植物密度(个/L),n为两次观测到藻类数量的平均值,v为1 L水样经沉淀浓缩后的体积(mL);
Pi=Ni/N
式中,Pi为第i个类群在群落中的数量占比,N为群落个体总数,Ni为第i个类群的数量;
H′=-∑Pi×log2Pi
式中,H′为香农威纳指数(Shannon-WienerIndex);
J′=H′/log2S
式中,J′为均匀度指数(Pielou’s Evenness Index),S为群落种群数量;
式中,D′为辛普森多样性指数(Simpson’s diversity index);
Y=Pi×fi
式中,Y为种群优势度(大于0.02即为优势种群),fi为第i个种群出现的频率。
2 结果与分析
2.1 水体环境因子
2.1.1 水体理化性质及主要营养成分 官地库区水温空间上整体表现为上游低于下游;季节上则表现为冬季转向夏季过程中水温逐渐升高,而后再降低的循环;全年水温10.9~22.6 ℃,平均值14.9 ℃。溶解氧空间上仅冬季各断面差异较大,表现为上游高于下游,其它季节各断面无明显差异;季节上则整体表现为冬季逐渐降低至秋季后再升高的循环;全年溶解氧为8.5~16.2 mg/L,平均值为10.8 mg/L。冬、春和秋季各江段pH值均无明显差异,在8.35~8.46小幅变化,仅秋季各江段明显高于其它季节;全年pH呈弱碱性,变幅8.35~8.97,平均值8.51。透明度在空间上总体表现为自上而下逐渐升高的趋势,仅秋季A断面高于其它断面;季节上均表现为自秋季逐渐降低至夏季后再升高的循环,其中夏季远低于其它季节;全年透明度为21~534 cm,平均值为238 cm。硬度在空间上各断面无大的差异,季节上则均表现为春季逐渐降低至秋季,而后再升高的循环;全年硬度为2.5~6.0 mmol/L,平均值为4.0 mmol/L。秋、冬和春季各断面水体电导率均无明显差异,在239.0~263.2 μs/cm小幅变化,仅夏季各断面水体电导率明显较低,全年平均电导率为243.4 μs/cm。春、夏和秋季各季节内各断面COD均无明显差异,仅冬季各断面COD有较大差异;从季节上COD则均表现为自春季逐渐升高至冬季,而后转为降低的循环;全年COD为1.3~10.6 mg/L,平均值为5.1 mg/L。
官地库区秋、冬和春季各季节内各断面总氮含量无明显差异,仅夏季为自上而下快速升高;从季节上均表现为自夏季逐渐升高至冬季,而后再下降的循环;全年总氮为0.10~0.94 mg/L,平均值为0.54 mg/L。总磷从空间上无明显变化趋势,季节上总体表现为自春季逐渐升高至秋季,而后再下降的循环;全面总磷含量为0.006~0.085 mg/L,平均值为0.042 mg/L。氮磷比仅在冬季表现为自上而下逐渐降低,其它季节均为自上而下逐渐升高的趋势;从季节上仅A断面表现为自冬季逐渐降低值秋季,而后再升高的循环趋势,其它断面均为自春季逐渐降低至秋季而后再升高的循环趋势;全年为2.3~91.6,平均值为29.4。
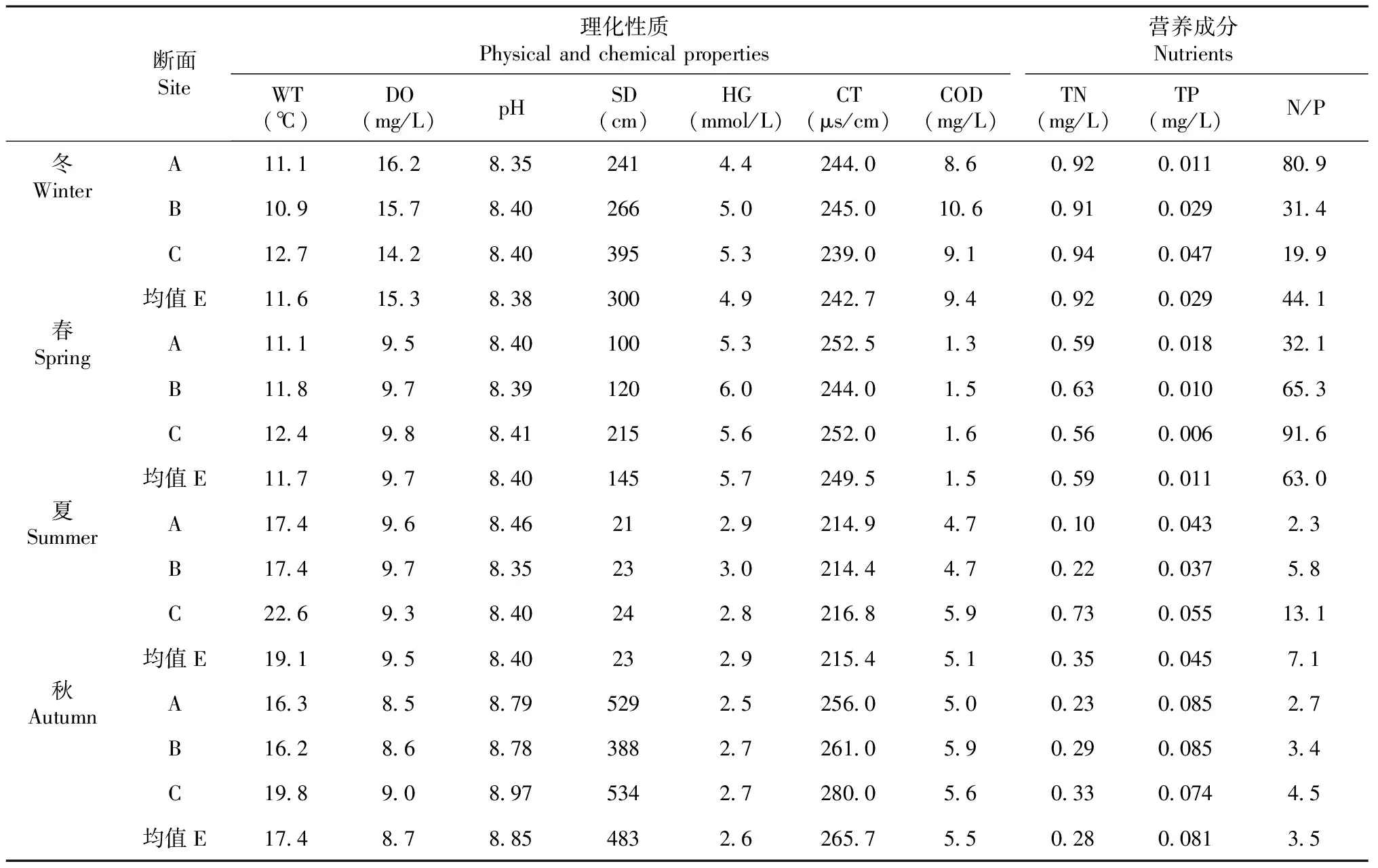
表2 官地库区水体理化性质及主要营养成分
2.1.2 水体环境因子间Pearson相关性 环境因子中除pH和总磷外均与水温呈负相关关系,其中溶氧、总磷、氮磷比与水温相关性显著,而硬度与之达极显著相关;化学需氧量、总氮与溶氧均呈极显著正相关关系;pH与透明度、硬度、电导率、总磷的相关性均达显著,其中与电导率、总磷为极显著正相关关系;透明度与pH、电导率、总磷均呈显著正相关关系,且前两项更达极显著程度;硬度与水温、pH、总磷均呈负相关关系,而与总氮、氮磷比均呈正相关关系,其中与水温、总氮相关性达极显著;电导率与pH、透明度均呈正相关相关,且后者相关性达极显著;总氮与溶氧、硬度均呈正相关关系,且后者相关性达极显著;总磷与水温、pH、透明度均呈负相关关系,与硬度、氮磷比均呈正相关关系,其中与pH、硬度、氮磷比相关性均达极显著;氮磷比与水温、硬度、总磷分别呈显著负相关、极显著正相关、极显著负相关关系。
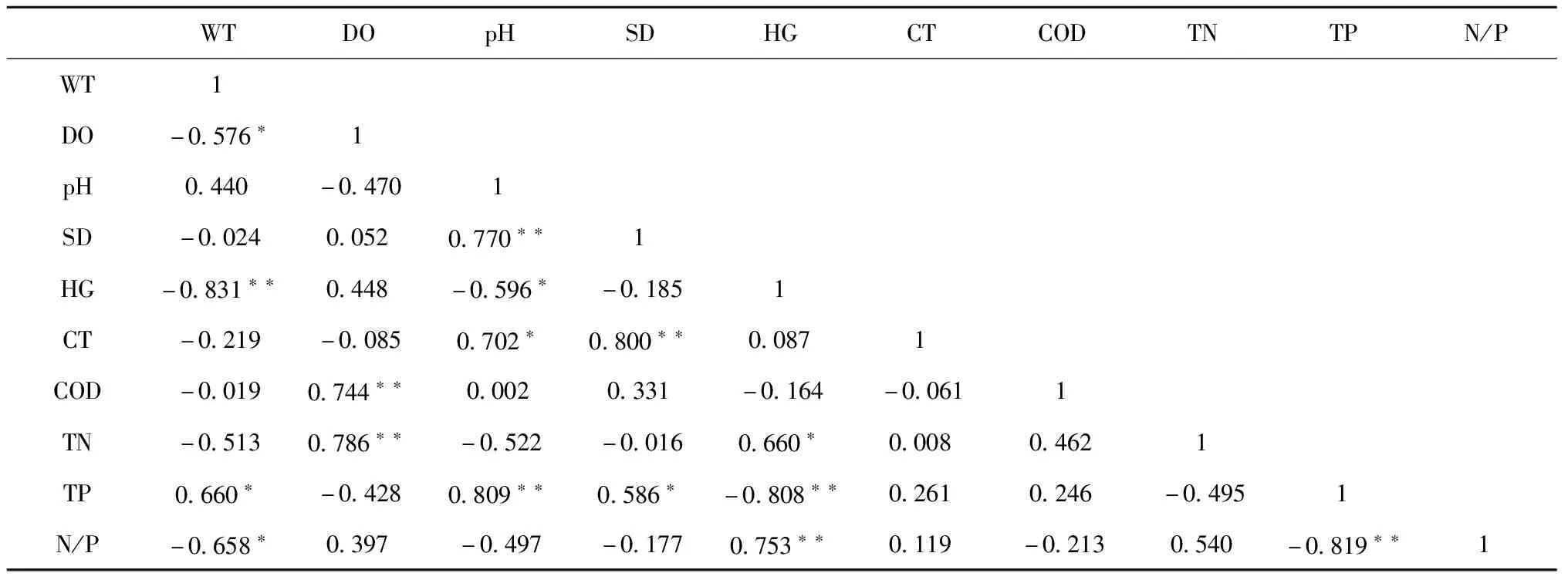
表3 水体环境因子间Pearson相关性
2.2 浮游植物群落结构
2.2.1 浮游植物群落多样性指数 官地水库浮游植物群落香浓威纳指数均高于1(图2),其中仅B断面的冬、春季低于2,且明显低于同季节的其它断面,而冬季A和C、夏季A和B断面高于3;全年香农维纳指数变幅为1.01~3.80,平均值为2.50。均匀度指数空间上均表现为B断面最低,C断面最高,仅夏季完全相反;季节上A断面表现为自春季逐步升高至冬季后转而降低的循环,B断面则表现为自春季升高至夏季后转而逐步降低的循环,C断面则为春夏季向秋冬季升高的趋势;全面均匀度指数变幅为0.27~0.78,平均值为0.59。辛普森指数空间上变化趋势与均匀度指数相同;季节上A和B断面均为夏季逐步降低至春季后转为升高的循环,而C断面春季显著高于其它季节;全年辛普森指数变幅为0.25~0.88,平均值为0.67。综合比较,C断面冬春季多样性指数均显著低于其它断面和季节,秋季各断面上各类多样性指数均无显著差异。
2.2.2 浮游植物群落组成及藻类密度 官地库区全年鉴定出6门97种(属)藻类(图3),其中硅藻门40种(属),绿藻门33种(属),裸藻门10种(属),蓝藻门7种(属),甲藻门4种(属),隐藻门3种(属);冬季共有6门42种(属),春季共有6门45种(属),夏季共有6门56种(属),秋季3门16种(属);总体来看,硅藻门和绿藻门种(属)数在全年和各季度均为最多,而秋季藻类门类数和种(属)数均显著低于其它季节。全年藻类平均密度为139.8×104cell/L(图4),空间上藻类密度自上游而下逐渐升高,A、B、C断面藻类密度分别为98.6×104、134.7×104、186.2×104cell/L;冬至秋季藻类密度分别为25.3×104、194.83×104、244.53×104、94.8×104cell/L,表现为自夏季降低至冬季后升高的循环。
2.2.3 浮游植物群落优势门类 从全年视角来看(图5-a),官地库区绿藻数量占比达42.5%,远高于其它类群,是绝对优势门类;蓝藻和硅藻数量占比分别达24.2%和19.3%,是亚优势门类。从季节视角来看(图5-b),绿藻数量除春季占比低以外,其它季节占比均为最高,是绝对优势门类;而蓝藻仅在春季占比最高,为绝对优势门类;硅藻仅在夏季占比略低于10%,其它季节占比在25%上下小范围波动,为亚优势门类。从断面视角来看(图5-c),冬季A、C断面优势门类均为绿藻和硅藻,B断面蓝藻为绝对优势门类;春季A、B断面绝对优势门类分别为硅藻和蓝藻,C断面优势门类为硅藻和蓝藻;夏季A断面隐藻和绿藻为优势门类,B、C断面绿藻均为绝对优势门类;秋季各断面均以绿藻为绝对优势门类;总体上绿藻为优势门类的断面和季度最多,硅藻次之。
2.2.4 浮游植物群落优势种群 从全年视角(表4)来看,优势种群有26种(属),其中硅藻和绿藻门各有8种(属),蓝藻门有4种(属),裸藻和隐藻门各有2种(属),甲藻门仅1种(属);而优势度大于0.1的仅有硅藻门美丽星杆藻,绿藻门湖生卵囊藻、卵形衣藻、小球衣藻、小球藻,蓝藻门不定微囊藻、水华微囊藻,隐藻门卵形隐藻,其中小球衣藻优势度最高,达0.713。
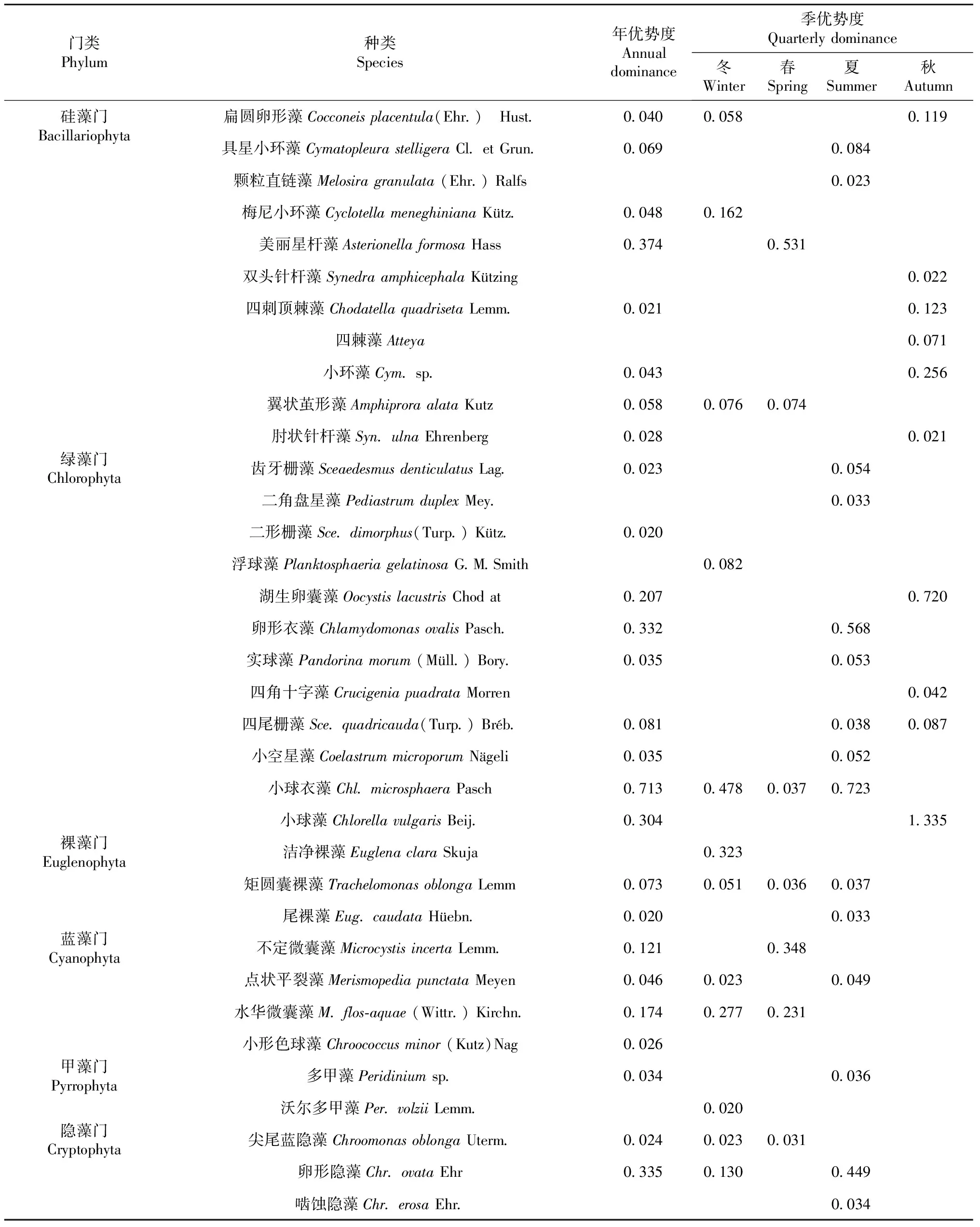
表4 不同时间尺度下各藻类种群优势度
从季节视角来看,冬季优势种群有12种(属),各门类分布较为均匀;春季优势种群有7种(属);夏季优势种群有15种(属),主要集中在绿藻门;秋季优势种群有10种(属),仅分布在硅藻和绿藻门。优势度大于0.1,冬季有硅藻门梅尼小环藻、绿藻门小球衣藻、裸藻门洁净裸藻、蓝藻门水华微囊藻、隐藻门卵形隐藻;春季有硅藻门美丽星杆藻、蓝藻门不定微囊藻和水华微囊藻;夏季有绿藻门卵形衣藻和小球衣藻、隐藻门卵形隐藻,秋季有硅藻门扁圆卵形藻、四刺顶棘藻、小环藻以及绿藻门湖生卵囊藻、小球藻。总体上看,绿藻门小球衣藻在冬春夏季均为优势种群,而绿藻门小球藻在秋季优势度为全年最高。
从断面视角(表5)来看,冬季A断面绿藻门小球衣藻为绝对优势种群,B断面蓝藻门水华微囊藻为绝对优势种群,C断面硅藻门翼状茧形藻、绿藻门浮球藻和小球衣藻、裸藻门洁净裸藻数量占比均超过10%;春季A、B、C断面绝对优势种群分别为硅藻门美丽星杆藻、蓝藻门不定微囊藻、蓝藻门水华微囊藻;夏季A断面优势种群为隐藻门卵形隐藻,B断面优势种群为蓝藻门点状平裂藻,C断面优势种群为蓝藻门卵形衣藻和小球衣藻;秋季绿藻门小球藻和湖生卵囊藻为各断面优势种群,其中小球藻为B断面绝对优势种群。不同季节各断面中,占比超过5%且出现次数超过3次的藻类分别有硅藻门美丽星杆藻和绿藻门湖生卵囊藻、小球衣藻、小球藻,其中小球衣藻高达5次。

表5 各断面不同季节占比大于5%的藻类种群
2.3 浮游植物群落结构与环境因子相关性分析
2.3.1 浮游植物群落多样性指数与环境因子冗余分析 第一轴和第二轴特征值分别为0.897和0.089(图6,Lengths of gradient=0.184),两轴对浮游植物群落多样性的解释量为98.6%。香农威纳指数、均匀度指数和辛普森指数均与化学需氧量、透明度、电导率、溶解氧呈显著正相关性,与水温呈显著负相关性;种群数量与氮磷比、硬度、总氮、溶解氧均呈显著正相关性,与总磷、水温、pH均呈极显著负相关性。硬度、水温、化学需氧量和总磷是驱动多样性指数变化的主要环境因子。
2.3.2 浮游植物门类与环境因子冗余分析 第一轴和第二轴特征值分别为0.403和0.262(图7,Lengths of gradient=1.209),两轴对浮游植物群落门类的解释量为66.5%。硅藻门和绿藻门与透明度、pH、总磷、电导率均呈正相关性,与溶氧、总氮、硬度、氮磷比均呈负相关性,而裸藻门与硅藻门和绿藻门几乎相反;蓝藻门与水温、化学需氧量均呈显著负相关性,与氮磷比呈显著正相关性,而甲藻门几乎相反;隐藻门与化学需氧量、溶氧均呈显著正相关性,与电导率呈显著负相关性。
2.3.3 浮游植物优势种群与环境因子冗余分析 第一轴和第二轴特征值分别为0.356和0.160(图8,Lengths of gradient=3.762),两轴对浮游植物群落多样性的解释量为51.6%。美丽星杆藻与除电导率、透明度外的环境因子相关性均达到显著,而湖生卵囊藻与之相似;卵形衣藻仅与pH、化学需氧量、透明度、电导率等均呈显著负相关性,与其它环境因子相关性很弱;小球藻仅与总磷、pH、化学需氧量、透明度、电导率等均呈显著负相关性;小球衣藻与总磷、pH、化学需氧量外的环境因子相关性均达显著;不定微囊藻、点状平裂藻和卵形隐藻均与电导率、透明度、化学需氧量、pH呈显著负相关性,水华微囊藻与之几乎相反。按图8可将占比大于5%的藻类种群大致分为III类,第I类仅与pH、化学需氧量、透明度、电导率相关性较强,与其它环境因子关联较弱;第II类与第I类几乎完全相反,第III类又几乎与第I类相同。由sol文件可知,pH、透明度、电导率和总氮是驱动占比大于5%藻类种群变化的主要环境因子。
3 讨 论
3.1 水体环境因子
根据《地表水环境质量标准》(GB 3838—2002),官地水库水体溶解氧、pH、化学需氧量等指标全年均处于I类水质标准范围内。冬春季水体总氮含量变幅为0.56~0.94 mg/L,处于III类水质标准范围内;夏秋季绝大多数断面介于0.2~0.5 mg/L,且平均值分别为0.35和0.28 mg/L,可视作处于II类水质标准范围内。由于官地水库属于高山峡谷河道型库区,水体交换量大[16],为日调节水库[11],库区有一定流速,可采用一般地表水总磷标准来衡量其水质类别。冬春季水体总磷含量很低,绝大多数断面低于0.02 mg/L,且两季平均值为0.020 mg/L,可视作处于I类水质标准范围内;夏秋季水质总磷变幅为0.37~0.85 mg/L,处于II类水质标准范围内。总体看来,官地水库水体溶解氧高、呈弱碱性、营养物质含量低,属于寡营养水体,水质处于I~III类之间。
官地水库水温夏秋季升高,而夏秋季也是雅砻江流域雨季来临的时节,雨水增多入库流量扩大带来的磷元素促使总磷升高、氮磷比下降,但水量猛增也稀释了原本水体中的氮、钙、镁等营养元素,使总氮和硬度迅速降低。夏季由于雨水过多造成洪流,致使该季度水体透明度骤降;秋季上游更清澈的水体持续汇入库区,促使水体透明度迅速加深。一般水体硬度与电导率呈显著正相关性[17],但官地库区水体硬度和电导率变化趋势完全不同,相关性很弱,这可能是入库水体硬度和电导率差异较大,致使汇合时两者“中和”造成硬度与电导率变化不一致。总体看来,官地水库属于日调节型,水体交换量大,水质受入库水体影响较深,呈现较为明显的季节变化趋势。
3.2 浮游植物群落多样性
官地水库浮游植物群落香农威纳指数全年平均值为2.50,水质总体处于β-中污带,夏季属于清洁水质;均匀度指数绝大多数大于0.50,全年平均值为0.59,水质总体均匀度良好;辛普森指数绝大多数大于0.50,全年平均值为0.67,表明水体总体良好。研究表明[18-20],水温升高有助于浮游藻类快速生长和繁殖,影响浮游植物群落丰度的主要环境因子,与本研究中藻类密度与水温呈极显著正相关性结论一致。官地水库常年水温较低,冬春夏季水温升高促使部分种类迅速生长,虽致使整体群落密度和种群数量增加,但部分藻类密度比例增长速度高于种群数量增加的效果,从而造成群落香农威纳指数、均匀度指数和辛普森指数均与水温呈负相关性;而且秋季总氮和氮磷比持续降低造成种间竞争激烈[3],使得群落密度和种群数量快速下降,而种群数量降低的效果又相对高于密度,从而形成前文和种群数量与水温呈显著负相关性的结果。总体上,水温为浮游植物群落种群数量和藻类密度的限制因子;调查前期总磷也为限制因子,后期则总氮也为限制因子[21]。
3.3 浮游植物群落优势类群
官地水库冬春季以硅藻门、绿藻门和蓝藻门藻类为主,夏秋季以硅藻门和绿藻门藻类为主,硅藻为全年主要类群之一预示着水质良好[22];总体上为绿-硅藻水相,与国内大多数水库、湖泊类似[23-25]。通常大型水库和湖泊冬春季水温低,更有利于硅藻生长,而更喜高温的蓝藻则在水温较高的夏秋季成为优势类群[26-28]。由于官地水库夏秋季为雨季,雨水增多,入库增大,水体流速加快,此时有较硬硅壳、体积和重量均较大的硅藻可像河中泥沙一样借助一定的纵向流速与流速梯度等产生表面压力所形成的浮力来悬浮于水中[29],从而占据生长优势成为优势类群;而冬春季水体流速低,体积和重量较小且具备气囊的蓝藻占据生长优势成为优势类群[13, 30]。所以官地水库浮游藻类藻相主要随季节发生变化,而水体理化因子和营养含量则为所适应的藻类种群助力。
猜你喜欢
当代水产(2022年6期)2022-06-29
当代水产(2021年8期)2021-11-04
军事文摘(2020年20期)2020-11-16
大自然探索(2020年4期)2020-06-19
流行色(2019年10期)2019-12-06
装饰装修天地(2019年9期)2019-10-21
农业与技术(2019年11期)2019-07-12
小品文选刊(2016年21期)2016-11-27
知识窗(2013年7期)2013-05-14
