
远缘杂交转育抗烟草花叶病N基因同源基因片段的初步研究
2021-09-23 13:33:02焦芳婵童治军方敦煌肖炳光袁欣婕陈学军李永平
种子 2021年8期
焦芳婵, 童治军, 方敦煌, 肖炳光, 袁欣婕, 陈学军, 李永平
(1.云南省烟草农业科学研究院, 昆明 650021;2.江西省农业科学院蔬菜花卉研究所, 南昌 330200)
普通烟草花叶病(Tobacco mosaic virus,TMV)是云南等烟叶主产区分布广、为害重的主要病害之一[1],TMV广泛存在于活体寄主、植物残体、大田土壤,即使在低温和高温条件仍能保持一定的病毒侵染活力[2],而且其寄主范围非常广,能侵染茄科(烟草、番茄、茄子)、葫芦科(西瓜)、十字花科(油麦菜)等[3]。
选育抗TMV的品种,是应对TMV为害最为经济和有效的手段。野生烟草-心叶烟中含有显性单基因TMV免疫基因N,前人已将N基因转育到了栽培烟草,N基因也是人类首次克隆的植物抗病基因[4]。我国利用该抗源已选育10多个高抗TMV的烟草品种[5-9],但由于连锁累赘的存在,迄今仍没有适宜的高抗TMV的烤烟品种进行大规模推广种植[10]。除了利用烟草基因组序列数据设计引物,从大批量的杂交后代筛选短的抗TMV片段外[11],从野生烟草资源中,转育新的TMV抗源也是应对措施之一。Yuan等[12]报道,在圆锥烟草(N.paniculataPI 555550)等烟属野生种中,其高抗TMV的特性由N基因家族其他成员提供。但迄今尚未见N基因同源基因片段(简称N同源片段,下同)转入栽培烟草的报道。本试验利用烤烟品种红花大金元与圆锥烟草(N.paniculata)开展种间远缘杂交研究,以期将新的TMV抗性基因导入栽培烟草,创制新的抗病种质资源,为选育抗TMV的烟草品种提供材料与数据支撑。
1 材料与方法
1.1 试验材料
所用材料为烤烟主栽品种红花大金元(简称HD,染色体2 n=48)、圆锥烟草(N.paniculataPI 555550简称Np 555550,染色体2 n=24)、含有N基因的栽培烟草RBST及野生烟草(N.glutinosa)由云南省烟草农业科学研究院提供。
1.2 杂 交
分别以HD、Np 555550做父母本,进行正反杂交,每个组合进行20个左右杂交,以纸管法套袋,一周后统计杂交结实率(即坐果数与杂交花的比值)。
1.3 杂交种SSR及GISH分析
以QIAGEN DNeasy plant kit试剂盒提取杂交种及双亲DNA。SSR-PCR扩增体系为20 μL,其中DNA样品1.5 μL(15~30 ng·μL-1)、10×PCR Reaction Buffer(Mg2+plus)2 μL、25 mmol·L-1dNTPs 1.2 L、10 μmol·L-1正、反向引物各2 μL、rTaq0.75 U,最后加水至20 μL。PCR反应程序为:95 ℃预变性5 min;95 ℃变性30 s、57 ℃退火30 s、72 ℃延伸30 s,共计30个循环。参照Bindler等[13]、童治军等[14]所用SSR引物表,筛选10对SSR引物进行SSR分析。
基因组前处理,液氮研磨提取体细胞杂交种基因组DNA,用Hind Ⅲ酶切纯化,用ROCHE的DIG HIGH PRIME DNA标记试剂盒对基因组进行地高辛标记。GISH分析参照Mark W.Chase等[15]的方法进行标记探针、压片、烘片、制片预处理、洗脱等处理。
1.4 Np基因特异标记开发及杂交种的抗性鉴定
参照Lewis R.S等[10]及Yuan等[12]的方法,分别合成N基因及Np基因特异引物,其中Np基因特异引物经过PCR筛选,选定solo-F 1-TGATGCACAAAGATCCCGTT+solo-R 2-GACCTCTGTGCATTCATCTTATCA为Np基因特异引物。以双亲及杂交种、RBST及N.glutinosa为模版,进行N基因片段及Np基因片段PCR扩增。
1.5 TMV接种抗性鉴定
参考周文兵等[16]的方法,即半叶枯斑法接种TMV,在25 ℃温室自然光照下,以10 mg·mL-1的0.2 mL花叶病病毒汁液摩擦接种半叶,另外一半涂去离子水(ck),共接种6片叶子,观察杂交种是否有枯斑来判定抗病性。
2 结果与分析
2.1 Np 555550和HD的远缘杂交
以HD及Np 555550为双亲,通过正、反交两种方法进行杂交(见表1)。由表1可以看出,染色体数目多的栽培烟草红大为母本,其杂交结实率为0,而以圆锥烟草为母本,其杂交结实率达36.8%。

表1 红花大金元及圆锥烟草N.paniculata(PI 555550)杂交结实结果Table 1 Fruiting results of cross between N. tabacum (Honghuadajinyuan) and N. paniculata (PI 555550)
2.2 杂交种的SSR及GISH鉴定
10对SSR引物中,TM 100有稳定的扩增效果,其序列为:TM 100 F 5’-TGGAGGAACCAACAAGGAAG-3’及TM 100 R 5’-GTCCGACAGTATCTTCGCAA-3’。远缘杂交种F1含双亲带(图1)。
F1杂交种的GISH鉴定表明,母本用地高辛标记,用Rhodamine anti-DIG-sheep显红色荧光,父本用生物素标记,用Alexafluor 488 Streptavidin显绿色荧光,母本父本重合的染色体是黄色荧光。红色荧光共计8条,绿色荧光共计4条,黄色荧光(红绿重合)共计28条,总染色体条数共计40条(图2)。
2.3 Np基因特异标记开发及杂交种的抗性鉴定
N基因片段及Np基因片段PCR扩增结果(图3~图4)表明,圆锥烟草及杂交种不能扩增出540 bpN基因特异片段,而可以扩增出500 bp大小的Np基因片段。
对红花大金元、N.paniculata及其F1苗期人工接种TMV,接种7 d后,红花大金元出现花叶症状,N.paniculata和杂交种全部为枯斑、对TMV表现免疫(表2、图5)。
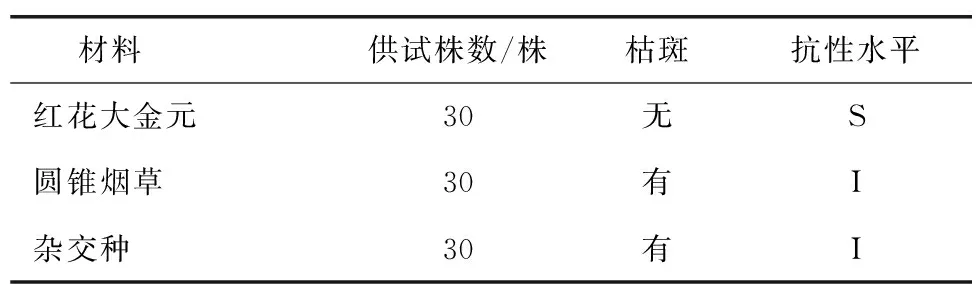
表2 杂交种TMV抗性鉴定结果Table 2 Identification of TMV resistance of hybrid
3 讨 论
截止2017年,国内烟草相关研究单位对2 000余份种质资源(包含重复鉴定资源)开展过TMV抗性鉴定[14-22],其中大部分表现枯斑的烟草资源包括烤烟、白肋烟、香料烟、晒烟、雪茄烟,不包括黄花烟、野生烟等[11],且这些资源的抗性都来自于N基因。目前国内外利用抗TMV资源选育的抗TMV烟草品种因为连锁累赘,在生产上的推广年限不长、推广面积不大。寻找新的TMV抗源是势在必行,而在野生烟草中具有普通烟草所不具有的一些重要基因,尤其是抗病抗虫基因。
Yuan等[12]研究表明,N.paniculata烟草中Np基因(GenBank:KP 278475)编码区有3 393个核苷酸,与N基因的编码区序列有205个SNPs差异,一个8核苷酸的缺失和一个3核苷酸的插入,核苷酸一致性为93.9%。Np基因中8核苷酸的缺失导致其后11个氨基酸序列与N蛋白中的序列不同。在203个位于8核苷酸缺失前的SNPs中,有137个引起氨基酸替换。203个SNPs中23个位于LRR区域内的高度变异位点,其中21个引起氨基酸的替换。其余116个位于8核苷酸缺失前的非同义突变SNPs分布于不同的区域:14个位于TIR区域,34个位于NBS区域,27个位于LRR区域,41个位于其他区域。N基因与N同源片段的高度变异位点的Ka/Ks比值为1.87,说明这两个基因在N.glutinosa和N.paniculata物种分化后受到了多样化选择。本研究以圆锥烟草为母本,将圆锥烟草的N基因同源基因Np片段转育到栽培烟草红花大金元中,为烤烟抗病育种提供了新的种质材料。
H.C.Sharma[17]在其他作物的远缘杂交结果表明,远缘杂交由于亲本间的基因组成存在较大差异,杂种一般以染色体数较多或染色体倍性高的作母本,更易克服杂交障碍获得远缘杂交种。本研究以染色体数少的圆锥烟草为母本,栽培烟草为父本也同样获得了杂交种,但其具体的分子机理有待下一步开展深入研究。另外,N基因同源基因Np片段导入栽培烟草后,其是否也存在较大的连锁累赘,是否也对烤烟产质量有较大的影响,仍有待下一步开展回交转育后进行比较分析。
猜你喜欢
科学(2023年2期)2023-05-30 08:57:46
环境科学与管理(2023年3期)2023-04-29 03:02:27
麦类作物学报(2022年5期)2022-05-20 09:43:24
今日农业(2021年14期)2021-11-25 23:57:29
四川蚕业(2020年2期)2020-07-10 03:14:48
科学导报(2018年47期)2018-05-14 12:06:10
现代园艺(2017年21期)2018-01-03 06:41:41
上海农业学报(2017年3期)2017-04-10 12:39:24
中国种业(2016年11期)2016-12-01 05:43:51
现代农业(2015年5期)2015-02-28 18:40:52
