
瘤胃微生物在木质纤维素价值化利用的研究进展
2021-09-22 08:56:24郑娟善丁考仁青李新圃梁泽毅张剑搏杜梅丁学智
草业学报 2021年9期
郑娟善,丁考仁青,李新圃,梁泽毅,张剑搏,杜梅,丁学智,*
(1.中国农业科学院兰州畜牧与兽药研究所,甘肃省牦牛繁育重点实验室,甘肃兰州730050;2.中国农业科学院兰州畜牧与兽药研究所,农业农村部兽用药物创制重点实验室,甘肃 兰州730050;3.甘南藏族自治州畜牧工作站,甘肃 合作747000)
木质纤维素是自然界最丰富的可再生生物质资源,全球每年通过光合作用产生的植物纤维约2000亿t[1],其中尚有89%未被利用,相当大的一部分按废弃物处理。农作物废弃原料(如秸秆)大多数被浪费或者露天焚烧,这不仅使大量的生物质资源无法价值化利用,还造成严重的环境污染。为缓解人为温室气体(greenhouse gases,GHG)排放量增加造成的全球气候问题,如何妥善处理农作物废弃原料和农业工业废弃物成为实施新时代乡村振兴战略亟待解决的问题[2-3]。近年来随着世界环境污染的日益严重和能源危机的不断加剧,木质纤维素价值化利用和资源化开发诸如发酵生产乙醇和沼气(甲烷)等潜在再生能源物质逐渐成为研究的重点[4-5]。同时,瘤胃微生物在生物燃料生产中高效降解木质纤维素的能力得到广泛的关注,以取代不可再生的化石燃料。然而,由于木质纤维素在组成和结构上异常的复杂,其降解需要多种微生物的协同互作,且微生物的分类和代谢的多样性以及各种纤维素酶间的协同降解机制尚不清晰,在一定程度上严重制约了木质纤维素资源的价值化开发和利用[6-8]。
反刍动物的肉和奶为人类提供高质量蛋白质、必需矿物元素和脂溶性维生素,同时也是人类饮食结构中饱和脂肪酸和反式脂肪酸的主要来源[9],据统计,反刍动物饲养量约占全球家养动物的76%。中国工程院院士、著名草业科学家任继周等[10]认为,未来我国粮食的压力并非来自人的口粮,而是饲料用粮,今后应以草食家畜代替耗粮型家畜来缓解饲料用粮的压力。我国拥有大量的农作物副产品(如秸秆),可以有效缓解畜牧业发展中粮食和牧草资源供求的矛盾。反刍动物瘤胃是自然界高度演化且降解木质纤维能力最强的生态系统之一,大量的微生物相互作用从而有效利用粗饲料中纤维素、半纤维素和非蛋白氮等成分,供机体营养需要及肉奶的产出[11],研究表明,利用瘤胃液中分离的菌株接种进行厌氧消化可提高玉米(Zea mays)秸秆[12]、小麦(Triticum aestivum)秸秆[13]和稻草的降解效率[14-15]。因此,反刍动物瘤胃微生物在宿主营养(为单胃动物作为新型益生菌和后生元)和工业应用(如在生物燃料生产、制浆造纸和纺织工业等方面的应用)中具有潜在的利用价值。本研究通过对相关文献的回顾和积累,将从以下4个方面进行详细综述:1)瘤胃内木质纤维素降解的微生物基础;2)降解木质纤维素相关酶;3)瘤胃酶和微生物作为饲料添加剂的研究;4)瘤胃微生物在木质纤维素价值化的研究,以期为瘤胃微生物在木质纤维素生物质转化为燃料、化学品和其他生物制品价值化利用等方面的研究和应用提供一定的理论参考。
1 瘤胃内木质纤维素降解的微生物基础
木质纤维素主要由纤维素(35%~50%)、半纤维素(25%~30%)和木质素(25%~30%)组成,并相互作用形成了具有高抗性和难降解的生物质结构(图1)[16]。随着生物技术及微生物组学的发展,瘤胃微生物被视为宿主体细胞“第一基因组”之后的“第二基因组”[17],不仅对宿主营养的吸收利用和畜产品质量至关重要,而且还为工业应用提供了潜在的机会[18-19]。因此,研究瘤胃微生物对木质纤维素的降解机理,提高木质纤维素瘤胃利用率并促使其在工业价值化利用的研究基础,是对瘤胃微生物菌群结构、功能和整体代谢途径的系统性认识。
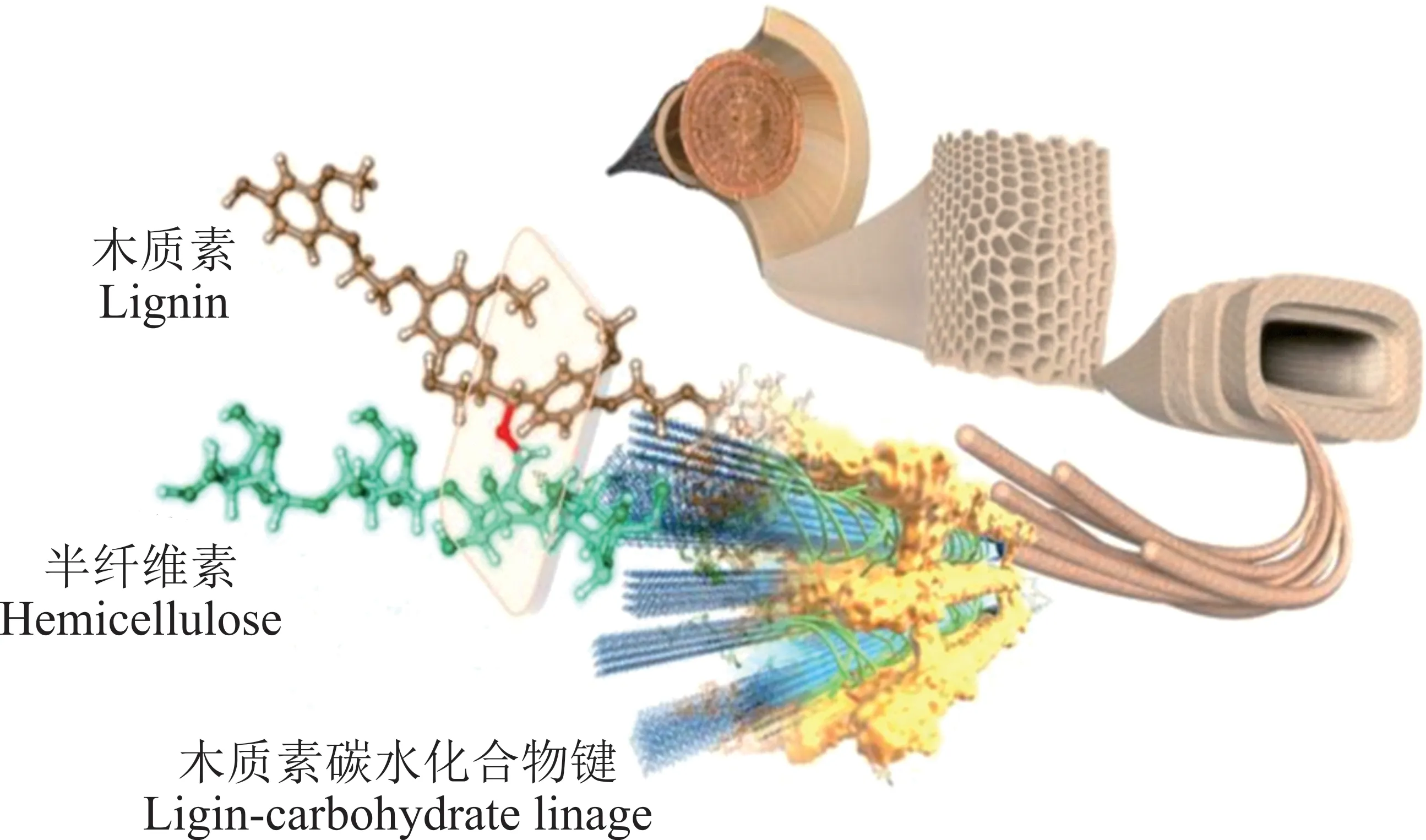
图1 木质纤维素结构Fig.1 Schematic of lignocellulose structure
木质纤维素表面组分(蜡质、纤维素、半纤维素和木质素)具有不同的氧/碳原子比(oxygen/carbon,O/C)和相对的C1~C3峰面积[20]。因此,O/C和C1~C3相对峰度的变化可以很好地反映纤维素表面组分变化。木质纤维素经瘤胃液发酵后其表面组分,特别是蜡质和木质素含量显著降低[21-22]。这意味着某些功能菌株(真菌和细菌)可以有效地去除蜡质和木质素的表面物质,增加可降解纤维素和半纤维素的可触性,从而实现瘤胃微生物对木质纤维素的有效降解。由于纤维素表面主要被蜡质和木质素颗粒覆盖,从而阻止了水解酶对木质纤维素的降解。Hu等[21]将小麦秸秆用瘤胃微生物厌氧发酵5 d后,原子力显微镜(atomic force microscope,AFM)清晰地显示了秸秆表面的纤维孔和裂缝,同时直观地观察了瘤胃微生物去除覆盖在秸秆表面蜡质和木质素颗粒的过程[21]。营养物质通过纤维孔扩散至微生物,并将溶解的碳转移到非纤维素分解微生物[23-24],这暗示着隧穿效应可能是瘤胃微生物降解秸秆的一种可能机制,同时瘤胃微生物并非完全降解纤维中所有蜡质和木质素组分。这为降解木质素的真菌在瘤胃的初始阶段占主导地位,但在随后逐步下降提供了合理的解释。同时,瘤胃厌氧真菌菌丝能够穿透植物细胞壁并溶解木质素成分,通过细胞分泌胞外游离的纤维素酶、半纤维素酶和酯酶等一系列高活性的酶,还可产生复合纤维小体降解木质纤维素[25]。瘤胃细菌对木质纤维素的降解模式与真菌略有不同,瘤胃细菌通过产生各种功能蛋白,组装成由催化区域、支架蛋白、碳水化合物结合模块(carbohydrate binding module,CBM)和黏连蛋白等构成的复合纤维小体,附着于细胞表面进而对木质纤维素进行分解[26]。
同时,反刍动物瘤胃中的细菌、真菌和原虫在木质纤维素降解过程中形成一个复杂的共生系统,共同参与植物细胞壁的降解。细菌数量庞大,并且有多种代谢途径[21,27],因此,瘤胃细菌在纤维素降解过程中占主导地位。瘤胃真菌在大颗粒、大片段植物纤维降解过程中起着重要作用。由于瘤胃真菌能产生一系列具有高度活性的酶,厌氧菌在植物细胞壁中占有优先地位,因此在降解植物细胞壁方面,真菌比其他微生物具有更大优势[28]。但由于瘤胃中真菌数量较少,对木质纤维素的降解机理和纤维素分解总量的贡献尚不明晰,在其生理生化特性、酶的结构和代谢途径等方面还需深入研究。高巍[29]发现,不同微生物区系的发酵速率和产气速率具有显著的变化,其中全瘤胃液的产气率和降解率最高,其次是原虫、细菌和真菌混合培养,而单独培养的最低。这说明真菌、细菌和原虫具有独立降解纤维素的能力,但效率不高;因此,瘤胃微生物对木质纤维素的降解不是某一种或者两种微生物所能及的,而是一个非常复杂的相互协同作用过程。另有研究报道,只有甲烷菌在寄生菌体内形成共生体,才能充分利用原生动物代谢的氢和甲酸产生甲烷[30]。由于缺乏对瘤胃微生物相互作用的深入研究,这些理论都需要进一步的验证。
2 瘤胃微生物降解木质纤维素相关酶的研究
瘤胃中纤维素类物质的降解是多种不同类型纤维素酶共同作用的过程。瘤胃微生物可分泌种类繁多且数量庞大的木质纤维素降解酶。真菌可附着于木质纤维素端部,厌氧菌游动的孢子和营养体菌丝可分泌一系列与木质纤维素降解相关的酶系,各种水解酶沿着假根进入植物细胞内部,对木质纤维素进行深度降解[31];细菌首先吸附于木质纤维素表面,然后从表面逐渐向内部生长,并将吸附在表面的木质纤维素结构进行破坏,从而降解木质纤维素[32]。无论微生物以何种方式进行木质纤维素降解,都以瘤胃微生物分泌的酶起主导作用,由于木质纤维素结构的复杂性,需多种木质纤维素降解酶的相互作用才能使其降解。
2.1 纤维素酶
糖苷水解酶(glucoside hydrolase,GHs)是水解碳水化合物分子与碳水化合物和非碳水化合物分子之间糖苷键连接的酶。纤维素酶是GHs的一个亚型,可降解纤维素中存在的糖苷-β-1-4键[33],纤维素酶由参与水解的三大类物质组成(图2):内切-β-1,4-葡聚糖酶(endo-β-1,4-glucanase),可降解纤维素中结晶度较低的区域,产生自由链末端;外切-β-1,4-葡聚糖酶(exo-β-1,4-glucanase),可从自由链末端切除纤维二糖或纤维丁糖;β-1,4-葡萄糖苷酶(β-1,4-cellobiase),可将纤维二糖等纤维寡糖水解为葡萄糖[34-35]。研究显示,主要的内切葡聚糖酶来自细菌的GH5、GH8和GH9家族以及真菌的GH6、GH7和GH45家族[36]。厌氧真菌菌丝能够穿透植物细胞壁并溶解木质素分泌一系列高活性的纤维素酶,高效降解植物细胞壁。Youssef等[31]表明瘤胃壶菌属(Piromyces)菌株CIA能够产生GH5、GH8和GH9中的内切-β-1,4-葡聚糖酶作用于纤维素分子链的非结晶区,水解β-1,4-糖苷键,且GH1和GH3家族中的β-1,4-葡萄糖苷酶,可有效降解纤维二糖、三糖和纤维素。Steenbakkers等[37]研究同样发现瘤胃壶菌菌株E2可产生β-1,4-葡萄糖苷酶和内切-β-1,4-葡聚糖酶,对纤维素的降解发挥着重要的作用。另外,也有研究表明瘤胃细菌如纤维丁酸弧菌(Butyrivibrio fibrisolven)和白色瘤胃球菌(Ruminococcus albus)也可分泌纤维素降解酶,在植物细胞壁的降解中发挥着一定的作用[38]。
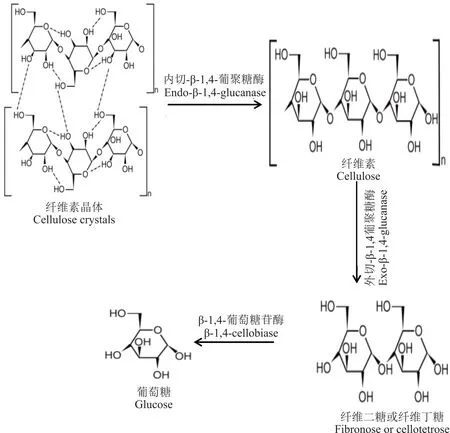
图2 纤维素酶催化机理Fig.2 Cellulase catalytic mechanism
2.2 半纤维素酶
半纤维素是非纤维素植物结构多糖,其包含通过各种键连接的己糖和戊糖[30]。由于半纤维素在单糖组成上的多样性和结构上的高度分支化,半纤维素降解酶系从种类和数量上都要比纤维素酶更加丰富和复杂。半纤维素酶是半纤维素(木聚糖、甘露聚糖、半乳聚糖和阿拉伯聚糖)降解酶的统称,如木聚糖酶、木糖苷酶、甘露聚糖酶、甘露糖苷酶和其他辅助酶[35](图3)。一般β-木糖苷酶(β-xylosidase)和β-1,4-内切木聚糖酶(β-1,4-endoxylanase)水解木聚糖主链,具有木葡聚糖酶活性的β-1,4-葡萄糖苷酶(β-1,4-glucosaccharase)和β-1,4-内切葡聚糖酶(β-1,4-endoglucanase)降解木聚糖,β-甘露糖苷酶(β-mannosidase)和β-1,4-内切甘露聚糖酶(β-1,4-endomannanase)水解甘露聚糖(hydrolyze mannan)。而一些辅酶如香豆酸酯酶(p-coumaric acid esterases)、葡萄糖醛酸酶(glucuronidase)、阿魏酸酯酶(ferulic acid esterase)、阿拉伯呋喃糖酶(arabinofuranosidase)等,主要负责降解木聚糖的支链部分[39-40]。Wang等[41]研究发现瘤胃厌氧真菌Neocallimastix patriciarumW5可分泌β-1,4-葡萄糖苷酶、β-1,4-内切木聚糖酶和β-木糖苷酶,有效降解半纤维素。已有研究表明,瘤胃厌氧真菌分泌的木质纤维素酶比市场应用的里氏木霉(Trichoderma reesei)和曲霉(Aspergillus nidulans)的活性更高,尤其是瘤胃壶菌属的酶活性[42]。Avgustin等[43]研究发现栖瘤胃普氏菌属(Prevotella ruminicola)和丁酸弧菌属(Butyrivibrio)具有来自不同家族GH的酶系,它们不仅具有水解淀粉和半纤维素的能力,还能水解蛋白质,并能同时发酵糖和氨基酸。Dodd等[44]研究也表明栖瘤胃普氏菌菌株能产生胞外木聚糖酶,可降解完整牧草中的半纤维素。同时,产琥珀酸纤维杆菌(Fibrobacter succinogenes)和黄色瘤胃球菌(Ruminococcus flavefaciens)会产生半纤维素酶,在半纤维素降解过程中有非常重要的作用[45]。
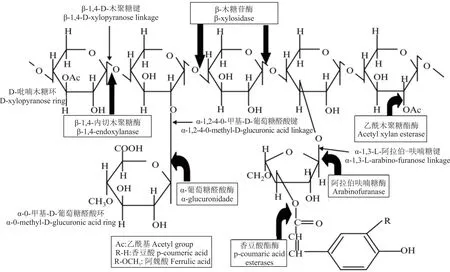
图3 具有不同取代基的木聚糖结构和木聚糖酶作用位点Fig.3 Xylan structure with different substituents and sites of attack by xylanase
瘤胃微生物也可分泌阿魏酸酯酶、乙酰酯酶、果胶酶。酯酶能够打开细胞壁中酚酸和半纤维素形成的酯键,使半纤维素从木质素和纤维素的复合体中释放出来;果胶酶能够降解植物细胞壁果胶成分,释放出酸性果胶和甲醇,这些物质可为其他降解酶的酶解提供反应底物,在植物细胞壁降解的初始阶段具有非常重要的意义[46]。
2.3 纤维小体
研究发现,瘤胃微生物还可合成高分子量和多功能的纤维小体[38],纤维小体是由厌氧微生物(细菌和真菌)产生的自组装蛋白质复合物,由结构蛋白和催化模块组成,其中结构蛋白包括支架蛋白、锚定蛋白等,催化模块包括降解植物细胞壁所必需的纤维素酶、半纤维素酶等[47-48]。众所周知,纤维小体对纤维降解具有很高的催化效率,这可能是由于催化亚基之间的邻近性导致有序的底物隧穿效应[26]。纤维小体能够有效降解木质纤维素,在植物细胞壁的黏附和降解中发挥着至关重要的作用,但是有关纤维小体的组装和作用机理并不清楚。
此外,很多水解酶都具有多模块化的特点,即同时含有一个催化域(catalytic domain,CD)和一个无催化活性的碳水化合物结合模块,它们通过一段柔性肽连接区(linker)连接[49]。CBM是碳水化合物活性酶内的连续氨基酸序列,其离散折叠具有碳水化合物结合活性,CBM通过配置和增加糖基表面单个糖苷水解酶或多酶纤维素复合体的有效浓度来增强纤维素的水解,且CBM可以分解植物细胞壁的纤维素结构,使其转变为小颗粒释放出来,在结晶纤维素降解的最初阶段起非常重要的作用[50-51]。真菌纤维素分解酶几乎完全拥有家族1的碳水化合物结合模块(CBM1)[36,52]。Liu等[53]研究发现瘤胃厌氧真菌Neocallimastix patriciarum含有家族1的碳水化合物结合模块,具有相对简单的分子结构,CBM1使催化域定位到木聚糖附近,增加木聚糖表面有效酶浓度,促进对木聚糖的吸附与水解;但随着真菌的菌丝生长到细胞壁时,分泌的酶向目标底物的传递可能不需要复杂的CBMs[54]。研究表明厌氧细菌白色瘤胃球菌含有家族37的碳水化合物结合模块(CBM37),该模块能识别纤维素、木聚糖、几丁质等多种底物,在纤维素酶黏附过程中作为桥梁将细菌和多糖底物衔接,并帮助酶固定在细菌膜表面,实现高效率的纤维降解[55-56]。同时,研究发现瘤胃细菌Fibrobacter succinogenes的C-末端含有富含脯氨酸的连接区序列连接的家族6碳水化合物结合模块(CBM6),在木聚糖酶与底物的相互作用中起关键作用[26,57]。CBM虽然能识别并能与纤维素和许多其他多糖特异性紧密结合,但由于CBM的种类繁多且结构复杂,使得在纤维素酶中的确切作用仍然未知。
3 瘤胃微生物在木质纤维素价值化的研究
对反刍动物而言,瘤胃中木质纤维素分解菌(木质素、纤维素和半纤维素降解菌)和非纤维素分解菌紧密关联,相互作用[39]。非纤维素分解细菌通过初级降解释放可溶性多糖,并通过发酵将其转化为乙酸、丙酸、丁酸、二氧化碳和氢气等,作为营养物质被家畜利用[73]。木质纤维素因资源丰富,成本低廉,成为生产瘤胃酶、饲料添加剂、生物乙醇、有机酸和沼气等生物制品的重要原料。因此,以木质纤维素为原料,利用瘤胃中的微生物菌群或从瘤胃中获得的酶生产饲料添加剂、生物燃料和化学用品,在应对日益增长的饲粮需求、能源需求和处理环境污染问题方面具有至关重要的作用。
3.1 瘤胃酶和微生物作为饲料添加剂的研究
谷物是动物饲料的最大组成部分。通常动物饲料中小麦(Triticum aestivum)、大麦(Hordeum vulgare)和黑麦(Secale cereale)等含有较高的纤维量,其主要由非淀粉多糖(non-starch polysaccharides,NSP)组成,如小麦和黑麦中的阿拉伯木聚糖及大麦和燕麦(Avena fatua)中的β-葡聚糖,这些抗营养化合物通过增加动物小肠消化液的黏度来干扰消化,降低营养物质的吸收效率并诱发肠道功能紊乱[58]。研究表明利用多种酶如纤维素酶、葡聚糖酶和果胶酶可以减少饲料中的抗营养化合物。添加其他酶如木聚糖酶和β-葡聚糖酶,可水解NSP,降低谷物的黏度,并释放蛋白质和淀粉等营养物质[59]。因此,在动物饲料中使用外源酶可以提高饲料利用率,使动物的消化系统更健康。Bansal等[60]研究表明,在单胃动物的饲料中添加外源性酶(如纤维素酶、木聚糖酶、果胶酶和葡聚糖酶),通过促进各种植物细胞壁聚合物(如纤维素、木聚糖、果胶和葡聚糖)降解为单糖来提高饲料成分的消化率,从而提高饲料的利用效率[61]。Romero等[62]发现在荷斯坦奶牛中添加木聚糖酶的复合酶制剂可显著提高奶牛干物质采食量、泌乳量和牛奶中乳脂含量。国春艳[63]研究了木聚糖酶和纤维素酶对表观消化性能的影响,发现在全混合日粮(total mixed ration,TMR)中添加木聚糖酶和纤维素酶不仅提高了3~7月龄后备牛干物质的采食量和酸性洗涤纤维(acid detergent fiber,ADF)、中性洗涤纤维(neutral detergent fiber,NDF)表观消化率,且提高了后备牛的总能表观消化率和日增重。综上,瘤胃酶作为饲料添加剂,不仅改善了动物的生长和健康状况,提高了饲料的利用率,而且有利于减少“黏性粪便”,改善牲畜的生存环境。
反刍动物因瘤胃微生物种群的存在,能够降解植物中木质纤维素。但是单胃动物无法消化各种植物细胞壁聚合物,其他多种饲料成分也不能被家畜完全消化吸收。在过去的几十年,科学家们一直试图通过增加瘤胃中纤维降解来改善瘤胃的功能,利用饲料添加剂如化学制剂[64]或酶[65]以提高木质纤维素的降解,但结果并不理想。近年来,使用活细菌作为益生菌实现纤维素的降解受到学者们极大的关注[66]。研究表明,从印度大羚羊中分离的瘤胃壶菌菌株FNG5添加在水牛的饲料中不仅提高了羧甲基纤维素酶、木聚糖酶、微晶纤维素酶、乙酰酯酶、阿魏酸酯酶和蛋白酶的酶活性,而且显著提高了干物质(dry matter,DM)、有机物(organic matter,OM)、NDF和ADF的消化率和挥发性脂肪酸(volatile fatty acid,VFA)的浓度,促进了水牛对营养物质的吸收[67]。Lee等[68]从山羊中分离的瘤胃壶菌菌株KNGF-2直接饲喂于绵羊,通过增加细菌和真菌的数量,从而增加营养物质的消化率和氮的利用,进而改变瘤胃中VFA的生成途径;但研究发现,真菌酶未能显著改善营养物质的消化、氮的利用和瘤胃发酵参数,这可能是由于日粮中蛋白质可能被瘤胃微生物迅速降解,以至于酶可能会降低瘤胃微生物的生长或活性。Dey等[69]在犊牛饲料中添加瘤胃壶菌后发现,真菌的添加可显著提高犊牛的平均日增重、饲料的转化率和粗纤维(crude fiber,CF)、NDF和ADF的消化率。将山羊中分离的瘤胃壶菌菌株CN6添加在青贮饲料时,可提高发酵30 d青贮饲料的DM、NDF和ADF体外降解率[46],这与Lee等[70]的研究结果一致。有研究在高粗料日粮条件下,直接灌服瘤胃真菌能够提高育肥牛日增重、泌乳牛产奶量,同时能够提高瘤胃内VFA浓度、游动孢子数量和饲料利用率[71-72]。以上研究表明,瘤胃真菌作为饲料添加剂可改善青贮饲料的发酵品质和营养物质,提高粗饲料的利用率,进而改善动物的生产性能,促进动物健康生长。
3.2 在生物化学(有机酸)生产中的应用
琥珀酸作为一种重要的有机酸,存在于动植物、人和微生物体内,在生物代谢中起着重要作用。琥珀酸来源于碳水化合物的发酵,广泛应用于生产食品、药物、绿色溶剂和生物分解性塑胶等[74]。工业上重要的化合物是由木质纤维素预处理后得到的可发酵糖产生,由细菌和真菌组成的不同类型微生物将木质纤维素降解为葡萄糖单体[36]。研究表明,瘤胃细菌如产琥珀酸丝状杆菌、黄色瘤胃球菌和溶淀粉性琥珀酸单胞菌(Amylolytic succinate)等进行发酵产生琥珀酸,这种细菌种类根据生产区域的不同,可以自由使用各种植物材料,如草或玉米,从而使琥珀酸的生产更可持续和更具成本效益[75]。利用瘤胃细菌埃氏巨球菌(Megasphaera elsdenii)可生产丁酸、己酸和戊酸等短链脂肪族有机酸,特别是利用生物方法生产己酸可能为尼龙开辟可持续生产的途径。同时由于埃氏巨球菌对短链脂肪酸的抗性,可作为一种有效的微生物细胞工厂(microbial cell factories,MCF)[76]。但利用瘤胃微生物生产有机酸的生物技术尚不成熟,需进一步了解其作用机制,进而开发应用于实际生产中。
3.3 在乙醇生产中的应用
在所有的液体生物燃料中,生物乙醇由于具有替代不可再生石油的潜力而受到全球的广泛关注。因此,利用不同的原料生产生物乙醇的各种技术实施已成为研究重点。生物燃料主要是从天然油脂中提取的生物柴油和从农业剩余物发酵中提取的乙醇[4]。木质纤维原料的残渣在世界范围广泛存在,是最有前途的可再生能源生产基质之一。可再生资源生产生物燃料的技术已得到应用。微生物可分泌协同酶,将复杂的生物质降解为单糖,并将其发酵生成乙醇[77],在生物燃料生产中发挥着重要作用。有关瘤胃微生物在木质纤维素发酵中生产乙醇的研究在国内外均有报道,且有研究表明瘤胃细菌在厌氧条件下通过发酵木质纤维素材料可产生乙醇和氢气[78]。Pang等[79]从内蒙古牛瘤胃中分离的大肠杆菌菌株ZH-4除了产生葡聚糖酶、β-葡萄糖苷酶和纤维素酶有效降解玉米秸秆,还能产生乙醇和氢气,这不仅为学者研究瘤胃微生物进化机制提供了一个很好的机会,也为利用合并生物工艺直接从大肠杆菌中生产生物燃料和化学品提供新的方法与思路。Banerjee等[4]也研究表明,大肠杆菌因在基因上易于操作且能够消耗大量的有机碳资源,从而用来生产包括生物乙醇在内的多种化学物质。同时,在厌氧条件下,白色瘤胃球菌和黄色瘤胃球菌主要的最终代谢产物为乙醇和氢气[26]。随着生物技术的进步,学者们已经开发出一种新的细胞循环分批发酵(cell recycle batch fermentation,CRBF)系统,可以利用植物生物质中的各种碳水化合物以及农业和工业产生的生物质并将释放出的糖发酵成乙醇[80]。因此,利用成熟的基因组工程构建既能水解又能发酵的联合生物加工菌株,是现代生物技术的重要突破。但因瘤胃微生物的复杂性和技术的不成熟,还需进一步研究关于宿主生物的生理和代谢、生物化学和酶的结构、原料组成等。
3.4 在沼气生产中的应用
沼气发酵是在厌氧条件下厌氧微生物将有机质(如秸秆和粪便)转化成沼气的一种发酵方式,是利用生物质能源的有效途径。但因农作物秸秆类生物质富含纤维素和半纤维素等难以降解的成分,成为沼气发酵的限速因素。研究发现利用农作物秸秆进行沼气发酵时添加瘤胃真菌可改善发酵品质,提高生物质降解率和沼气产量[5,81]。Aydin等[82]在微藻中接种不同量的瘤胃真菌在促进微藻发酵和降解的同时提高了甲烷的产量。李倩等[83]针对醋糟中木质纤维素厌氧消化处置率低的问题,通过接种瘤胃微生物强化木质纤维素水解,表明瘤胃微生物的参与可显著提高底物产沼气和甲烷的产量,说明瘤胃微生物的介入可有效强化体系底物的降解能力,进而提升其产甲烷性能。Barragán-trinidad等[84]也证实了这一观点,他们利用瘤胃液作为水解微生物的来源对天然微藻的预处理进行研究发现,瘤胃微生物和微藻的混合可提高水解率和甲烷的产量,这可能是由于瘤胃真菌更专注于微藻细胞壁的降解。Li等[85]利用连续搅拌槽厌氧反应器以瘤胃液为接种菌在造纸剩余污泥的原料中生产沼气,沼气中CH4和CO2的含量分别为65.1%和34.2%。与以往研究结果相比,接种瘤胃微生物可显著提高甲烷产量,表明在现有厌氧系统中加入厌氧剂,可有效地生产沼气。魏亚琴[86]的研究表明,来自天祝放牧牦牛瘤胃真菌Neocallimastix frontalis和甲烷菌M.ruminantium共培养可高效将秸秆转化为CH4和乙酸,说明厌氧真菌应用在沼气发酵时能促进秸秆类生物质的水解,提高发酵过程中甲烷菌的数量,加速秸秆的降解,增加沼气的产量。
因此,瘤胃微生物在沼气发酵工程领域中具有广阔的应用前景,目前对瘤胃微生物的研究已经从体外培养、反应机制和性能、原始的反刍动物营养学向实用化角度转变[87]。但大部分研究仍基于传统思路,对瘤胃微生物在厌氧消化系统中还处于局部的纤维水解性能方面,忽略了瘤胃微生物菌群的复杂性,且利用瘤胃微生物将秸秆转换为沼气还处于可行性探索阶段,对降解机理和协同代谢过程尚不明确。因此,如何利用好瘤胃微生物这一天然的发酵微生物菌群,并将其应用到实际生产中是一个十分值得研究的问题。
4 展望
木质纤维素在可再生替代能源方面起着至关重要的作用,并日益成为世界上最大的能源资源之一。尽管国内外有关瘤胃微生物和瘤胃酶在木质纤维素转化为沼气、生物乙醇、有机酸及其他化学制品等相关的研究已取得了丰硕的成果,但由于缺乏对瘤胃微生物结构复杂性和各种木质纤维素酶互作机制的详细了解,且大多数瘤胃微生物无法在体外培养,从而阻碍了瘤胃微生物潜力的发挥。故对于瘤胃微生物在降解木质纤维素及价值化利用方面还需进行以下几方面研究:
深入探讨和研究瘤胃微生物降解木质纤维素的作用机理及瘤胃微生物与降解木质素相关酶间的相互作用机制,分离纯化获得低成本高效降解木质纤维素的纯化酶,为纯化酶在制浆造纸、纺织工业、食品和动物饲料等工业上的应用提供理论支撑。
瘤胃作为有效降解木质纤维素的微生态系统之一,不仅微生物多样、酶资源丰富,且发酵体系独特。因此,发现高效降解木质纤维素的新型菌株,研究新培养条件,同时构建分解木质纤维素复合菌系,探明复合菌系形成的机制,进一步了解复合菌系的机理和特点,并挖掘其应用潜力,拓展复合菌系的应用途径,以期在乙醇、沼气和有机酸等生物制品得到高效低成本生产。
通过综合基因组技术(基因组学、转录组学、蛋白组学、代谢组学等)的联合应用,对厌氧发酵中木质纤维素降解菌进行深入研究,挖掘菌株信息,明晰代谢通路,找到编码特定酶类的目标基因,揭示微生物群落的遗传潜力,以期利用基因工程获得快速、廉价、高效降解木质纤维素生物质的微生物和酶类资源,应用于实际生产。
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23 06:23:18
林产化学与工业(2022年4期)2022-09-13 09:47:12
中国饲料(2022年5期)2022-04-26 13:42:34
湖南饲料(2019年5期)2019-10-15 08:59:10
创新作文(小学版)(2018年7期)2018-08-20 03:11:02
国际木业(2016年8期)2017-01-15 13:55:23
国际木业(2016年3期)2016-12-01 05:04:53
国际木业(2016年1期)2016-12-01 05:04:11
兽医导刊(2016年12期)2016-05-17 03:51:35
兽医导刊(2016年12期)2016-05-17 03:51:31
