
两种等级喀斯特石漠化地区生物结皮对土壤养分恢复的影响
2021-09-09 04:47郑智恒熊康宁容丽池永宽
生态环境学报 2021年6期
郑智恒 ,熊康宁 *,容丽 ,池永宽
1. 贵州师范大学喀斯特研究院,贵州 贵阳 550001;2. 国家喀斯特石漠化防治工程技术研究中心 贵州 贵阳 550001;3. 贵州师范大学地理与环境科学学院,贵州 贵阳 550001
生物结皮又称生物土壤结皮(Biological soil crust,BSC),由菌类微生物、藻类、地衣、苔藓等隐花植物通过其菌丝体分泌多聚糖胶结土壤沙粒后形成的地表复合体,是生态系统中的重要组成部分(韩炳宏等,2017)。凭借着独特的生理结构与强大适应机制广泛分布于石漠化、荒漠化、极地等恶劣生境中,BSC的演替发育在土壤的形成过程中起到重要的作用。它能够有效改善土壤结构与理化性质,提高土层的稳定性和生产力,减少水土流失,一定程度上有效防控地质灾害的发生(张元明,2005)。促进土壤环境中物质能量循环与积累,调控水文过程,并进一步促进植被的发育演替以及提高地区生物多样性建设。基于不同的母岩基质背景下,BSC的群落结构有一定的差异,群落异质性与生态因子也存在区别,诸多因素对 BSC的物种构成、植株高度、个体密度、植被覆盖度等生理指标与生态类群等也会产生一定的影响(吉雪花等,2014)。喀斯特地区的BSC研究报告表明,结皮植被广泛存在于该地貌区各类型土壤表层,且不同石漠化小生境下的 BSC群落构成有所不同(李军峰等,2015;申家琛等,2017)。
石漠化是喀斯特地区基于母岩成土时间长、土壤贫瘠、基岩裸露度高、植被覆盖率低等特殊生境背景下,叠加过度的人类经济行为而形成的一种石质荒漠化景观,是一种土地退化现象(Zhang et al.,2020)。熊康宁等(2012)根据岩石裸露度、植被覆盖度、生物多样性等指标将石漠化等级分为无石漠化、潜在石漠化、轻度石漠化、中度石漠化、强度石漠化和极强度石漠化6个等级。其中,土壤的理化性质是反映土壤质量关键因素,对喀斯特石漠化地区土壤环境物质能量守恒及生境间耦合机制与生物多样性建设等研究具有重要意义。核桃树(Juglans)、花椒树(Zanthoxylum)是喀斯特地区治理石漠化的特殊经济型林木,在撒拉溪研究区和花江研究区推广并应用,为当地的生态建设与经济建设带来了一定的收益。但由于核桃树、花椒树生长周期长,枝、叶稀疏,对降雨下渗与径流侵蚀拦截作用有限,同时这类型经济林对土壤中养分掠夺较强,林下灌、草等植被很难存活,极易造成林下大面积地表裸露,水土流失、地下漏失等现象越发严重,进一步诱发石漠化形成(Miralles et al.,2012;Song et al.,2017)。BSC是喀斯特石漠化地区重要的地表覆盖植被,与不同类型的生境契合度高、抗逆性强,能直接通过特殊的生理结构借助根系从体表直接吸收水分和养分,促进土壤中养分含量的积累,有效改善土壤理化性质,在维持喀斯特土壤系统的动态平衡过程中扮演重要角色(Zhao et al.,2015;Zhou et al.,2020)。因此,在核桃林、花椒林下培育BSCs能有效减少外营力对土表的侵蚀,获得较大的生态收益。主流的石漠化治理多采用“林-灌-草”植被组合模式、混农林立体发展模式搭配水土漏失阻控技术等方式,由于喀斯特地貌区岩石大面积裸露于地表,地形分割显著,植被立地条件差,不适合草本、木本类植物的大量种植,常规的防治手段所需周期长且生态收益不稳定(Ying et al.,2014;Antoninka et al.,2020)。与传统的石漠化治理方法相比,人工培育BSC前期成本低,中期培植容易,后期养护方便,生长周期较短且存活率高,耐胁迫能力好、生态价值高、与不同生境的兼容度高(Gao et al.,2018)。依靠其良好的生态修复功能成为环境治理的先锋植物,可为现有的喀斯特石漠化防治措施提供新型的补充(郑智恒等,2020)。
目前,尽管BSC对土壤理化性研究已有部分报告,但大多集中于荒漠化地区,针对喀斯特石漠化地区生物结皮的相关研究仍十分缺乏(符裕红等,2020),该区的研究主要集中在喀斯特地区BSC群落的分类构成、演替习性分析,植物持水性及成土能力等部分功能特性方面(李冰,2009;程才等,2019;杨睿等,2021)。为此,本研究选取以潜在-轻度石漠化为主体的高原山地区和以中度-强度石漠化为主体的高原峡谷区作为不同等级的石漠化研究区域,系统分析不同石漠化生境背景下生物结皮对土壤中 10种关键理化指标的影响(土壤化学指标:pH值、全氮、速效氮、全磷、速效磷、速效钾、土壤有机碳;土壤物理指标:土壤容重、土壤田间持水量、土壤孔隙度),探讨BSC对不同石漠化生境下土壤理化性的影响及其对土壤养分恢复的效用,以期为利用生物结皮进行石漠化生态防治提供科学依据。
1 研究地区与研究方法
1.1 研究区概况
本研究选取中国南方喀斯特石漠化地区中最具代表性的贵州石漠化区域作为研究区,选取研究区Ⅰ:毕节撒拉溪喀斯特高原山地潜在—轻度石漠化典型区域(以下简称撒拉溪研究区),研究区Ⅱ:关岭—贞丰花江喀斯特高原峡谷中度—强度石漠化典型区域(以下简称花江研究区)。
研究区Ⅰ(表 1)位于贵州省毕节市撒拉溪镇(27°12′30″—27°16′50″N,105°03′10″—105°05′08″E),地貌类型以高原山地为主,该区地形破碎,地势起伏显著,平均海拔在1410—1780 m。流域年均降水量达900 mm,降水主要集中在6—10月,占全年降水量的50%以上,年平均气温13.8℃,相对湿度90%左右,无霜期在245 d以上,岩石以碳酸盐类的石灰岩为主,土壤类型以黄壤土以及紫砂土为主,潜在、轻度石漠化面积占该研究区总面积的68.65%以上。野生植被有厚果崖豆藤(Millettia pachycarpa)、火棘(Pyracantha angustifolia)、刺梨(Rosa roxbunghii)皇竹草(Pennisetum hydridum)、狼尾草(Pennisetum alopecuroides)、黑麦草(Lolium perenne)等组成的草、刺、藤、灌丛,以及零星分布的马尾松(Pinus massoniana)、响叶杨(Populus adenopoda)、杜鹃(Rhododendron simsii)、光皮桦(Betula luminifera)等林木(Li et al.,2020)。本研究区内不同地貌部位分布有藻类、地衣、混生和苔藓生物结皮,结皮总盖度为86%,其中藻类、地衣、混生和苔藓结皮的盖度分别为 11%、14%、23%和38%。生物结皮中藻类主要有爪哇伪枝藻(Scy-tonema javanicum)、具鞘微鞘藻(Microcoleus vaginatus)、隐鞘鞘丝藻(Lyngbya cryp-tovaginatus)等,地衣以皮果衣(Dermatocarpon miniatum)、盾鳞衣(Placidium squamulosum)、缠结茸枝衣(Seirophora contortuplicata)等为主,苔藓主要包括土生对齿藓(Didymodon vinealis)、丛生真藓(Bryum caespiticium)、小口小石藓(Weisia microstoma)、美灰藓(Eurohypnum leptothollum)、卷叶湿地藓(Hyophila involuta)、羽枝青藓(Brachythecium plumosum)等(Zhang et al.,2020)。
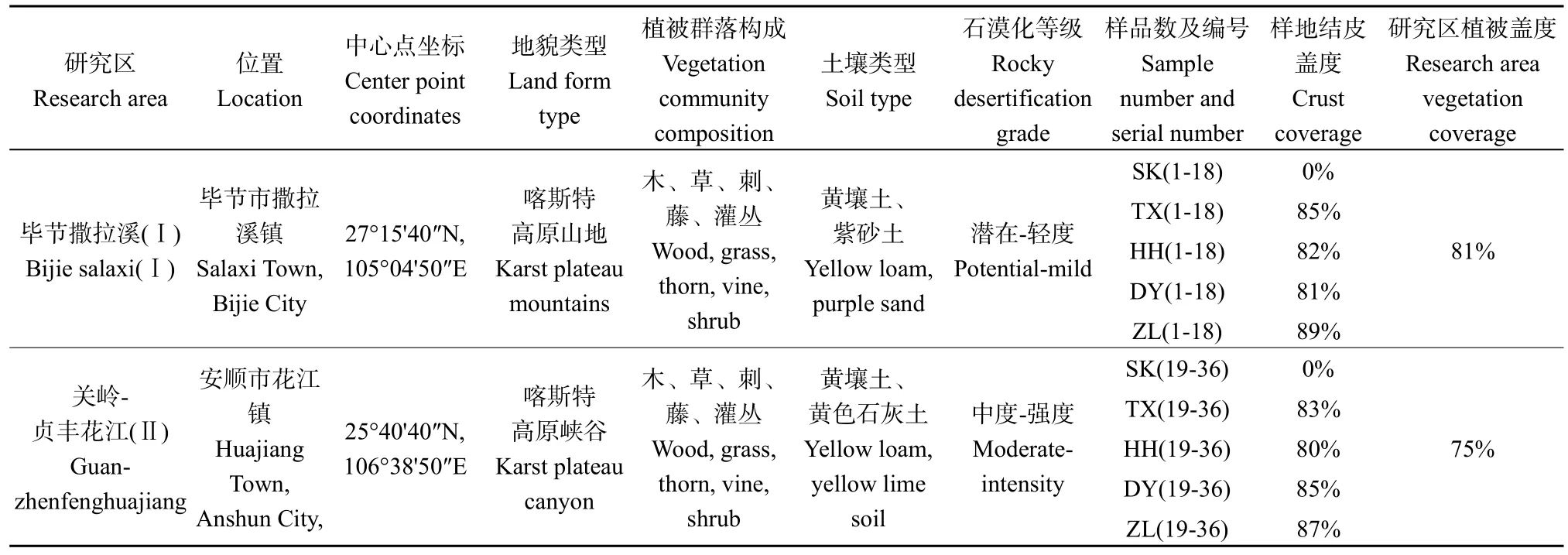
表1 试验区基础信息及样地设置Table 1 Basic information and plot settings of the experimental area
研究区Ⅱ(表 1)位于贵州省安顺市关岭县和黔西南州贞丰县交界处的峡谷两岸(25°39′20″—25°41′20″N,106°37′30″—106°39′49″E),地貌类型以高原峡谷为主,地形分割显著,高低落差较大,平均海拔在450—1450 m。该研究区流域年均降水量达1100 mm,降水主要集中在5—10月,占全年降水量的 80%以上,年平均气温 17.8 ℃,相对湿度80%左右,无霜期在300 d以上,岩石以碳酸盐类的石灰岩为主,土壤类型以黄壤和黄色石灰土为主,中度、强度石漠化面积占该研究区总面积的87.72%(Dai et al.,2018)。野生植被多以火棘(Pyracantha angustifolia)、刺梨(Rosa roxbunghii)、悬钩子(Rubus corchorifolius)、皇竹草(Pennisetum hydridum)、狼尾草(Pennisetum alopecuroides)等为主的草、刺、藤、灌丛,以及零星分布的柏木(Cupressus funebris)、构树(Broussonetia papyrifera)、马尾松(Pinus massoniana)等林木。本研究区内不同地貌部位分布有藻类、地衣、混生和苔藓生物结皮,结皮总盖度为82%,其中藻类、地衣、混生和苔藓结皮的盖度分别为12%、16%、20%和34%。该区生物结皮中藻类主要有爪哇伪枝藻(Scy-tonema javanicum)、具鞘微鞘藻(Microcoleus vaginatus)等,地衣以缠结茸枝衣(Seirophora contortuplicata)、盾鳞衣(Placidium squamulosum)等为主,苔藓主要包括土生对齿藓(Didymodonvinealis)、丛生真藓(Bryum caespiticium)、小口小石藓(Weisia microstoma)、美灰藓(Eurohypnum leptothollum)等(刘润等,2018)。
1.2 试验设计与样品采集
本实验选择在研究区中 10年以上的撂荒地内进行,以典型优势种藻类结皮(选取爪哇伪枝藻为研究对象,藻类盖度大于 80%)、地衣结皮(选取皮果衣为研究对象,地衣盖度大于 80%)、混生结皮(选取爪哇伪枝藻和皮果衣和美灰藓,苔藓、地衣和藻类混生,占比分别为5꞉3꞉2)和苔藓结皮(选取美灰藓为研究对象,苔藓盖度大于80%)所生长的地表土壤作为研究对象(以生物结皮为主要植被类型,无其他植被生长的土表层)。在撒拉溪示范区(Ⅰ)与花江示范区(Ⅱ)分别选取地势较平坦的区域,各设置3个15 m×15 m灌木样方,在每个大样方中设置3个3 m×3 m的草本样方,分别调查灌木及草本等植物群落构成及盖度。最后在样方内灌丛间BSC覆盖地段分别选取1 m×1 m的空白组(裸地)、藻类结皮、地衣结皮、混生结皮和苔藓结皮的实验小样方各3个(即3个重复)。按照S型布点法选取4种BSC组及空白组下方的结皮层和0—5、5—10 cm 3种土样层。于2019年9月采集样品,每个研究区各采取 45个物理和 45个化学土壤样品,共计180个混合样品。
1.3 样品处理与测定
所有土壤样品用PE塑封袋密封带回实验室内进行风干,去除残留的碎石、枯落物后过0.149 mm筛子,用于分析土壤的理化性质(高丽倩等,2018)。
(1)田间持水量采用重量法。
(2)结皮层土壤容重采用涂膜法,下层土壤容重与土壤孔隙度采用环刀法。
(3)土壤有机碳含量采用重铬酸钾氧化-外加热法测定。
(4)全氮测定采用高氯酸-硫酸消煮,凯氏定氮法测定。
(5)速效氮测定采用碱解扩散法。
(6)全磷采用高氯酸-硫酸消煮-钼锑抗比色法测定。
(7)速效磷采用氟化铵-盐酸浸提,钼锑抗比色法测定。
(8)速效钾测定采用1mol pH 7.0乙酸铵浸提-火焰光度法测定。
(9)pH值用5꞉1水土比浸提测定。
(10)土壤养分恢复指数(RI,Soil nutrient recovery index),以空白组为对照,计算BSC生长下的各种土壤理化性质与空白组间的差异,最后将各属性的差异求和平均,以定量描述不同类型生物结皮对土壤养分的恢复的作用(高丽倩,2018)。土壤恢复指数的计算公式:

其中,Xi为某一类型BSC下第i个土壤属性值,X'i为空白组第i个土壤属性值。
1.4 数据处理
运用Excel 2013和SPSS 22.0软件进行数据处理分析,利用单因素方差分析法比较不同类型BSC之间数值及其相对应下层土壤的理化性质差异,利用最小显著差异法(LSD法)进行显著性检验(α=0.05)。对不同石漠化环境BSC下层土壤的理化性质间进行Pearson相关性分析,同时对比BSC与空白组间差异,计算土壤恢复指数。运用 Origin 2018、Adobe Photoshop CS6软件进行制图。
2 结果与分析
2.1 生物结皮在撒拉溪潜在—轻度石漠化生境下对土壤理化性的影响
2.1.1 不同类型 BSC对下层土壤物理性质变化的影响
由图1可知,撒拉溪研究区内不同类型的BSC在结皮层中土壤田间持水量和土壤孔隙度表现为苔藓结皮>混生结皮>地衣结皮>藻类结皮>空白,而土壤容重则表现出相反的规律,空白>藻类结皮>地衣结皮>混生结皮>苔藓结皮。不同类型 BSC的土壤田间持水量、土壤孔隙度、土壤容重均差异显著(P<0.05)。

图1 撒拉溪研究区不同类型结皮及下层土壤物理和化学性质Fig. 1 Physical and chemical properties of different types of crusts and their underlying soils in the Salaxi research area
在结皮层下0—5 cm与5—10 cm土层中,土壤田间持水量和土壤孔隙度变化规律与结皮层相似,均表现为苔藓结皮>混生结皮>地衣结皮>藻类结皮>空白(图 1)。相反,土壤容重在结皮下层土壤中为空白>藻类结皮>地衣结皮>混生结皮>苔藓结皮。除藻类结皮和地衣结皮间土壤空隙度差异不显著(P>0.05),混生结皮在5—10 cm土层中与藻类结皮和地衣结皮间不显著(P>0.05),两个土层中苔藓结皮与混生结皮,地衣结皮与藻类结皮间的土壤容重差异不显著(P>0.05),其余类型BSC间在0—5 cm与5—10 cm土层中差异均显著(P<0.05)。
土壤垂直剖面上,通过比较各土层间土壤田间持水量、土壤孔隙度、土壤容重的变化规律,除BSC土壤容重呈现出随着土壤深度而增加,其他指标中同类型 BSC表现出由结皮层向深层依次减小的趋势。在0—5 cm与5—10 cm土层中,地衣结皮土壤间孔隙度及藻类结皮土壤孔隙度、土壤田间持水量的差异不显著(P>0.05),其余类型BSC下层土壤间均差异显著(P<0.05)。
2.1.2 不同类型 BSC对下层土壤化学性质变化的影响
结皮层中全氮、全磷、有机碳、速效氮、速效磷、速效钾含量均表现出苔藓结皮>混生结皮>地衣结皮>藻类结皮>空白(图1)。其中全氮、全磷、有机碳、速效氮在不同类型 BSC间差异均显著(P<0.05);混生结皮、地衣结皮和藻类结皮间速效磷含量差异不显著(P>0.05),地衣结皮和藻类结皮间速效钾含量差异不显著(P>0.05),其他类型BSC指标在速效磷、速效钾中差异显著(P<0.05),不同类型BSC间的pH值差异不显著(P>0.05)。
在结皮层下0—5 cm以及5—10 cm土层中,全氮、全磷、有机碳、速效氮、速效磷、速效钾均表现出与结皮层相似的规律,均表现为苔藓结皮>混生结皮>地衣结皮>藻类结皮>空白(图1)。除地衣结皮和藻类结皮间速效钾含量差异不显著(P>0.05),混生结皮、地衣结皮和藻类结皮间速效磷含量差异不显著(P>0.05),以及苔藓结皮和混生结皮,地衣结皮和藻类结皮间速效氮含量差异不显著(P>0.05);其他参数在0—5 cm与5—10 cm土层中,不同类型BSC间差异均显著(P<0.05)。
土壤垂直剖面上,分析各个土层间全氮、全磷、有机碳、速效氮、速效磷、速效钾的变化规律,相同类型 BSC表现出由结皮层向深层依次减小的趋势,结皮层大于下层土壤。地衣结皮和藻类结皮在下层土壤间速效钾、有机碳含量差异不显著(P>0.05),同时混生结皮下层土壤速效钾差异不显著(P>0.05),其余类型BSC相关参数在各土壤层间均呈显著性差异。土壤pH值随着土层深度增加而增大,结皮层小于下层土壤,除苔藓结皮下层土壤间pH值差异显著外(P<0.05),其余类型结皮下层土壤间的pH值差异均不显著(P>0.05)。
2.2 生物结皮在花江中度—强度石漠化生境下对土壤理化性的影响
2.2.1 不同类型 BSC对下层土壤物理性质变化的影响
花江研究区内不同类型的 BSC在结皮层中土壤田间持水量和土壤孔隙度表现为苔藓结皮>混生结皮>地衣结皮>藻类结皮>空白,但土壤容重则为相反的规律,表现为空白>藻类结皮>地衣结皮>混生结皮>苔藓结皮(图2)。不同类型BSC中,除地衣结皮和藻类结皮的土壤孔隙度、土壤容重的差异不显著(P>0.05),苔藓结皮和混生结皮,地衣结皮和藻类结皮间的土壤田间持水量差异不显著(P>0.05),其他指标均差异显著(P<0.05)。
在结皮下0—5 cm与5—10 cm土层中,土壤田间持水量和土壤孔隙度变化规律与结皮层相似,均表现为苔藓结皮>混生结皮>地衣结皮>藻类结皮>空白。只有土壤容重在结皮下层土壤中表现为空白>藻类结皮>地衣结皮>混生结皮>苔藓结皮(图2)。在下层土壤中,苔藓结皮和混生结皮,地衣结皮和藻类结皮间的土壤田间持水量差异不显著(P>0.05),地衣结皮和藻类结皮间的土壤空隙度无显著差异(P>0.05),混生结皮、地衣结皮、藻类结皮三者间的土壤容重差异不显著(P>0.05),其余类型BSCs间在0—5 cm与5—10 cm土层中差异均显著(P<0.05)。
土壤垂直剖面上,对比各土层间土壤田间持水量、土壤孔隙度、土壤容重的数值差异,除土壤容重含量随着土壤深度而增加,其他指标同类型BSC表现出由结皮层向深层依次减小的趋势。不同类型BSC土壤容重在各层间差异显著(P<0.05),4种结皮的土壤孔隙度在结皮层和0—5 cm土层中无显著性差异(P>0.05),但与5—10 cm土层间呈显著性差异(P<0.05)。地衣结皮与藻类结皮的土壤田间持水量在结皮层和下层两个层次土壤间均无显著差异(P>0.05),其余类型BSC各个土壤层间的指标均差异显著(P<0.05)。
2.2.2 不同类型 BSC对下层土壤化学性质变化的影响
结皮层中全氮、全磷、有机碳、速效氮、速效磷、速效钾含量均表现出苔藓结皮>混生结皮>地衣结皮>藻类结皮>空白(图2)。其中全氮、有机碳、速效氮在不同类型BSC间差异均显著(P<0.05),除地衣结皮和藻类结皮间全磷、速效钾、速效磷含量差异不显著(P>0.05),其他类型BSC在全磷、速效钾、速效磷中差异显著(P<0.05)。不同类型BSC间的pH值差异不显著(P>0.05)。
在结皮层下0—5 cm以及5—10 cm土层中,全氮、全磷、有机碳、速效氮、速效磷、速效钾均表现出与结皮层相似的规律,均表现为苔藓结皮>混生结皮>地衣结皮>藻类结皮>空白(图2)。除地衣结皮与藻类结皮的全磷、速效氮、速效磷和速效钾含量差异不显著(P>0.05),其他参数在0—5 cm与5—10 cm土层中,不同类型BSC间差异均显著(P<0.05)。
土壤垂直剖面上,对比结皮层及下层土壤间全氮、全磷、有机碳、速效氮、速效磷、速效钾的数值差异,相同类型BSC表现出由结皮层向深层依次减小的趋势,结皮层大于下层土壤。地衣结皮和藻类结皮在结皮层与 0—5 cm土层中有机碳含量差异不显著(P>0.05),但在0—5 cm与5—10 cm土层中速效磷含量差异不显著(P>0.05);同时地衣结皮和藻类结皮速效钾含量各层间变化不显著(P>0.05)。其余类型 BSC相关参数在各土壤层间均呈显著性差异(P<0.05)。土壤中 pH值随土层深度增加而增大,结皮层小于下层土壤,除苔藓结皮下层土壤间 pH值差异显著外(P<0.05),不同类型结皮下层土壤间的 pH值差异均不显著(P>0.05)。
2.3 生物结皮土壤理化性质相关性分析
2.3.1 撒拉溪研究区生物结皮土壤理化性质相关分析
由表 2可知,撒拉溪研究区内生物结皮覆盖下土壤pH值与全氮、速效氮、全磷、速效磷、速效钾、土壤有机碳、土壤容重、土壤田间持水量、土壤孔隙度指标占比不相关。土壤容重与全氮、速效氮、全磷、速效磷、速效钾、土壤有机碳、土壤田间持水量、土壤孔隙度呈显著负相关。除pH值和土壤容重外,其他指标间均呈显著正相关。

表2 撒拉溪研究区生物结皮土壤理化性质相关系数Table 2 Correlation coefficients of physical and chemical properties of biological crust soil in the Salarxi research area
2.3.2 花江研究区生物结皮土壤理化性质相关分析
由表3可知,花江研究区内生物结皮覆盖下土壤pH值与全氮、速效氮、全磷、速效磷、速效钾、土壤有机碳、土壤容重、土壤田间持水量、土壤孔隙度指标占比不相关。土壤容重与全氮、速效氮、全磷、速效磷、速效钾、土壤有机碳、土壤田间持水量、土壤孔隙度呈显著负相关。除pH值和土壤容重外,其他指标间均呈显著正相关。

表3 花江研究区生物结皮土壤理化性质相关系数Table 3 Correlation coefficients of physical and chemical properties of biological crust soils in the Huajiang research area
2.4 不同类型生物结皮对土壤养分恢复的影响
土壤养分恢复指数(RI)可以指示土壤恢复程度,本研究以空白组为对照,利用土壤恢复指数公式,计算不同类型BSC生长下的土壤恢复指数。由图3可知,在撒拉溪研究区:相比空白组,苔藓结皮、混生结皮、地衣结皮和藻类结皮的土壤养分恢复指数分别为25.1%,18.6%,10.4%和6.3%。由图4可得,在花江研究区:对比空白组,苔藓结皮、混生结皮、地衣结皮和藻类结皮的土壤养分恢复指数分别为25.8%,18.9%,7.6%和5.7%。研究表明,在不同石漠化等级地区,生物结皮对土壤养分均具有一定的恢复功能,不同演替阶段的BSC对土壤养分恢复程度有所差异,具体表现为苔藓结皮和混生结皮>地衣结皮和藻类结皮。
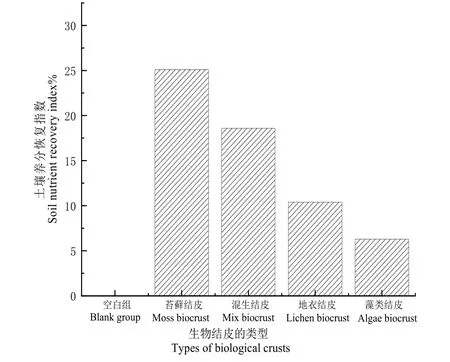
图3 撒拉溪研究区生物结皮对土壤养分恢复的影响Fig. 3 Impact of biological crusts on soil nutrient restoration in the Salarxi research area

图4 花江研究区生物结皮对土壤养分恢复的影响Fig.e 4 Effect of biological crusts on soil nutrient restoration in the Huajiang research area
3 讨论
3.1 生物结皮对喀斯特石漠化地区土壤理化性质及土壤养分恢复的影响
BSC是喀斯特石漠化地区重要的地表覆盖植被,凭借着特殊的生理结构与良好的适应机制维持着土壤生态系统的动态平衡(张元明等,2002)。对土壤环境的改善、水文环境的稳定、动植物的发育演替进程具有积极效用,在喀斯特生态系统的影响中扮演着重要角色。已有研究报告表明,各类型BSC的形态功能各异,组分构成多样,因此生态功能与生态收益具有一定的差异性(郑云普等,2009)。本研究中,在两个不同石漠化等级研究区内BSC覆盖下土壤的全氮、速效氮、全磷、速效磷、速效钾、土壤有机碳、土壤容重、土壤田间持水量、土壤孔隙度均表现为苔藓结皮>混生结皮>地衣结皮>藻类结皮(P<0.05)(图 1、2)。一方面,不同类型BSC中生物量存在差异,种群间差别一般体现为苔藓结皮>地衣结皮>藻类结皮。在演替发育过程中,BSC的分泌物及表层的凋落物和生物残体等被微生物通过分解作用形成腐殖土壤层,促进了土壤中碳、氮、磷、钾等养分的积累,这也是影响BSCs下土壤理化性的主要原因(朱远达等,2004)。此外,结皮植被起始于藻类、菌类的繁殖,随着浅层土壤环境的改善,地表盖度和粗糙度呈逐渐增大,捕获大气降尘的能力也逐渐增强(葛秋诗等,2021),结皮植被的厚度、土壤中细颗粒物质和养分含量不断增加,地衣结皮和苔藓结皮开始发育,此时结皮的蓄水保水能力、固土及含养功能显著增强(李茜倩等,2018)。在这个过程中,组成植物种从低等到高等,种群丰富度及群落多样性从小到大,BSC将随着时间的推进及微生境条件的改善完成它的演化,苔藓结皮作为演替末期的植被类型,在各类型BSC中拥有最高的生态功能。另一方面,BSC的演替有效改善地区的水热组合条件等环境因子,伴随着生物量的提高和生境条件的改善,结皮层及土壤层中微生物的生物多样性不断增加,进一步的加强了微生物的分解作用,保证了土壤中碳、氮、磷、钾等养分的有效积累(苏延桂等,2011)。
BSC是喀斯特石漠化区土壤重要的碳、氮来源,BSC中的叶绿素含量随着其演替发育而增加,通过光合作用、呼吸作用、代谢过程等生理调节功能,碳、氮固定能力逐渐增强。研究人员研究发现藻类结皮的年固碳量为 2.9—11.2 g·m−2·a−1、地衣结皮为 3.5—37.2 g·m−2·a−1、苔藓结皮为 26.8—64.9 g·m−2·a−1(张健等,2019)。同时,BSC 覆盖下的土壤氮转化速率及其对温度的敏感性均高于无结皮土壤,土壤中供氮能力得到加强,进而提升氮转化和循环效率,对于土壤系统的平衡稳定至关重要。实验人员在贵州喀斯特地区相关研究明确其固氮活性介于2.5—61.5 μmol间,其中藻类结皮平均固氮活性 (14.5 μmol)<地衣结皮 (19.7 μmol)<苔藓结皮 (28.2 μmol),BSC的年平均固氮量在13.3—38.13 mg·m−2·a−1之间,得出藻类结皮<地衣结皮<苔藓结皮(从春蕾等,2017)。研究者对南非莫洛波盆地中BSC的理化性质进行了研究,结果表明BSC可以通过固定和保持碳和氮的能力来促进土壤中碳、氮的稳定增长(Thomas et al.,2006)。实验人员在腾格里沙漠和科尔沁沙漠也都得出了 BSC能显著提高土壤中碳、氮、磷、钾等养分的结果(Hu et al.,2015;李新荣等,2018),这与本文的研究结果一致。揭示了BSC对生态系统中碳、氮等元素输入以及转化为土壤中养分供给植物生长的事实,证明喀斯特生境下BSC对碳、氮、磷、钾循环的关键作用以及对土壤养分积累做出的突出贡献。此外,本研究中两个研究区土壤容重表现为藻类结皮>地衣结皮>混生结皮>苔藓结皮,这与研究者在黄土沙漠区被区研究结果相同(杨巧云等,2019)。
除pH值外,在撒拉溪及花江研究区内结皮层与下层土壤间其他理化性质相关性均呈显著性(表2、3),说明结皮层与下层土壤理化性质的变化规律具有同步性。对比土壤垂直剖面数值,BSC的结皮层与0—5 cm土层养分含量明显高于5—10 cm土层,表现出由结皮层向深层依次减小的趋势,这与研究人员在澳大利亚南部地区及在中国新疆古尔班通古特沙漠地区及在中国黄土高原地区研究得出的结论一致(Bao et al.,2005;Steggles et al.,2019;Zhang et al.,2019),验证了BSC能够影响地表下的土壤理化性质并且对下层土壤的影响由浅及深呈递减趋势。此外,在这两个不同等级石漠化研究区中各类型的BSC因演替周期不同,结构组成及生态功能各异,导致不同BSC所覆盖的土壤养分存在一定差异,本研究表现为藻类结皮<地衣结皮<混生结皮<苔藓结皮,这与研究者在中国新疆古尔班通古特沙漠地区,研究人员在伊朗干旱区的实验结论一致(Wang et al.,2018;Kakeh et al.,2020)。此外,BSC覆盖下的结皮层中养分在降水入渗过程中会发生淋溶作用,进而对土壤中下层养分含量产生影响,本研究结论与实验人员在腾格里沙漠的实验结论相反(都军等,2018),但与学者在贵州喀斯特地区,秦福雯等在中国西北地区所得结论相一致(秦福雯等,2019;郑智恒等,2020)。这是由于在干旱-半干旱沙漠地区降雨量少,降水入渗深度有限,入渗养分一般停留在5 cm以上的土表层,而本实验研究区流域降雨量大,土表养分随着降水入渗过程中会发生淋溶作用,影响着中下层土壤养分含量。另一方面,沙漠地区多以砂质土壤为主,在撒拉溪研究区和花江研究区内多以黄壤土和紫砂土为主,相比于沙漠地区土壤中碳、氮、磷等养分含量更高,综合对比气候、水文、植被类型及盖度、地质地貌等诸多因素,撒拉溪及花江研究区拥有较好的土壤环境条件。因此,实验中土壤的碳、氮、磷、钾等养分含量相比干旱-半干旱沙漠地区数值更高。此外,两个研究区中土壤pH值在不同类型BSCs中差异不显著,但随土层深度增加而呈增大趋势,这与研究者在西班牙的塔韦纳斯沙漠及中国腾格里沙漠的研究得出的结论一致(Miralles et al.,2012;Song et al.,2017)。原因可能是,在 BSC演替发育过程中,BSC中隐花植物大量繁殖,随着光合作用和呼吸作用等生理功能的增强,造成植物分泌的有机酸及根系中微生物分解物不断积累以及降雨入渗后中和了酸碱度,多因素共同导致该地区土壤pH值变化不明显。
中国南方喀斯特地貌区,生境条件复杂,降雨量大,基岩裸露度高,母岩成土速率较慢,地表侵蚀严重且自然灾害频发,极易形成石漠化现象。BSCs通过其独特的功能特性有效改善了土壤环境,对喀斯特土壤系统中土壤的形成发育,土壤养分的有效累积与循环以及维持土壤环境稳定性具有特殊意义。本研究结果验证了BSC在不同喀斯特石漠化生境下对土壤环境中碳、氮、磷、钾等元素含量存贮与积累以及对土壤养分恢复所做出的突出贡献(图1—4)。
4 结论
不同类型 BSC在各等级石漠化地区对土壤理化性质影响各不相同,但均对地表下土壤养分积累具有良好的效用,有效改善了喀斯特生态脆弱区土壤环境状况。对比各类型BSC的土壤理化指数及土壤养分恢复指数,实验结果表现为苔藓结皮>混生结皮>地衣结皮>藻类结皮,演替末期的苔藓结皮拥有最高的生态效益。在土壤的垂直剖面上,各类型BSCs下层土壤中碳、氮、磷、钾等养分含量均表现为随土壤深度加深而降低,说明BSC对表层土壤影响最为显著。除土壤pH值外,不同类型BSC结皮层与下层土壤间理化性质均呈显著相关。此外,对比各类型研究区中BSC对土壤养分含量的影响,可以发现在不同生境背景下 BSC对土壤理化性的影响规律相似但影响程度存在着一定差别,这主要由于在不同区域内土壤类型、土壤温湿度、气候条件、动植物群落构成、地形地貌等诸多生态因素差异所致。BSC依靠独特的可抗逆性与良好的生态修复功能成为环境治理的新型植被,未来建议加强喀斯特生境下BSC的人工培植技术、BSC与喀斯特生境间耦合机制、BSC与喀斯特岩溶过程的互作机制、喀斯特地区BSC耐胁迫特性及生物多样性稳定机制等方面的相关研究。
猜你喜欢
绿色科技(2022年4期)2022-03-21
现代园艺(2021年19期)2021-12-08
军事文摘(2021年18期)2021-12-02
润·文摘(2021年11期)2021-11-20
当代水产(2021年8期)2021-11-04
内蒙古林业调查设计(2021年2期)2021-06-29
潍坊学院学报(2020年2期)2021-01-18
军事文摘(2020年20期)2020-11-16
防护林科技(2020年3期)2020-05-13
今日农业(2019年10期)2019-01-04
