
亚热带针阔混交林土壤-植物-大气连续体(SPAC)中水稳定同位素特征
2021-09-09 04:47:16王锐章新平戴军杰罗紫东贺新光关华德
生态环境学报 2021年6期
王锐,章新平, ,戴军杰,罗紫东,贺新光, ,关华德
1. 湖南师范大学资源与环境科学学院,湖南 长沙 410081;2. 湖南师范大学/地理空间大数据挖掘与应用湖南省重点实验室,湖南 长沙 410081;3. National Centre for Groundwater Research and Training, Flinders University, Adelaide, SA 5001, Australia
森林生态系统是陆地水分调节和物质交换的重要场所,对于维护区域生态平衡和环境保护具有重要的意义(娄源海等,2016;李龙等,2020)。大气降水作为水分的重要输入项对整个系统水分补给过程具有重要影响,而土壤中储存的水分又通过蒸发、下渗和植物蒸腾等方式进行重新分配,进而实现区域森林水循环过程的动态平衡(Luo et al.,2019;Dai et al.,2020;邓文平等,2017)。近年来,随着稳定同位素技术的逐渐成熟,利用稳定同位素示踪的方法可从微观上提取不同水体的水同位素信息,准确地揭示水分在区域森林生态系统中的运移规律(Dawson et al.,2002;Robertson et al.,2006)。相关研究表明,一个地区降水中稳定同位素变化特征与区域气候特征紧密相关,具体表现在降水、温度、湿度、风向风速、水汽输送过程以及云下二次蒸发方面的特征(吴华武等,2012;黄一民等,2013;章新平等,2015;周慧等,2018)。受降水补给的影响,土壤水很大程度上保留了降水中稳定同位素信息。与此同时,降水在入渗过程中受蒸发和前期土壤水分混合的影响,稳定同位素值发生明显变化,这种变化随深度增加逐渐减小(王锐等,2020a)。
植物作为联系土壤和大气的介质,是区域森林系统水循环过程中最重要的环节。了解植物水中氢氧稳定同位素变化有助于揭示其在局地水体氢氧稳定同位素循环中的贡献(Snyer et al.,2010;Stahl et al.,2013;罗伦等,2013)。研究发现,除少数盐生或旱生植物外,大部分植物在蒸腾之前体内水中稳定同位素不发生分馏,当各水源中稳定同位素存在差异时,通过比较植物茎杆水和各水源中稳定同位素组成可实现对植物水分利用来源的判别(Ehleringer et al.,1992;Brunel et al.,1995)。在植物蒸腾过程中,叶片中的水分通过气孔与外界水汽发生稳定同位素的交换,较轻的水分子会优先逸出叶片,使得叶片中水稳定同位素富集(Dongmann et al.,1974;Farquhar et al.,2003;Butt et al.,2010)。娄源海等(2016)研究发现,相较于湿润的雨季,高温低湿的旱季植物叶片水中δ18O更富集。李龙等(2020)研究证明,不同植物由于叶面积指数和叶龄的差异,叶片水中稳定同位素也存在较大的差异。植物叶片水中稳定同位素变化除了与环境水分状况和植物自身生理特征有关外,还受各气象因子影响(温学发等,2008;周盼盼等,2016)。王小婷等(2016)通过比较季节尺度和日尺度上玉米叶片水中稳定同位素变化与各气象因子的相关性发现,在不同时间尺度上影响叶片水中稳定同位素变化的因子也存在显著的差异。目前,尽管关于森林系统内部水循环过程中稳定同位素变化的研究较为丰富,但主要集中在干旱半干旱地区(Che et al.,2019;党宏忠等,2020),对湿润地区森林系统内部水循环过程中稳定同位素变化还研究较少,且相关研究多基于短时间历时的观测数据,无法发现各水体中稳定同位素长时间动态变化特征及其细节。
本研究以亚热带湿润地区长沙为研究区域,以针阔混交林内樟树(Cinnamomum camphora)和刺杉(Cunninghamia lanceolate)作为研究对象,基于2017年3月—2019年9月期间降水、土壤水、植物茎杆水和叶片水中稳定同位素数据以及两次典型晴日的植物茎杆水和叶片水中稳定同位素的逐时数据,并结合相关气象因子,分析了降水、土壤水和植物水中稳定同位素的季节变化特征以及植物水中稳定同位素的日内变化特征。研究目的在于:(1)揭示区域森林系统内不同水体稳定同位素变化的影响机制;(2)结合各气象因子,明确在不同时间尺度下影响植物叶片水中稳定同位素富集变化的主要因子。
1 材料与方法
1.1 研究区概况
研究场地位于湖南省长沙市望城区八曲河村(28°22′09″N—112°45′43″E),平均海拔在 50 m 左右。受亚热带季风的影响,研究区气候四季分明,雨热同期。多年平均降水量1415 mm,年内降水存在明显季节差异,在 3—7月初,受夏季风影响,降水丰富;7月中旬至9月,在副热带高压控制下,伏旱严重。多年平均气温17.4 ℃,大于0 ℃的多年平均年积温5457 ℃,全年无霜期约272 d,植物生长期在330 d左右。研究场地的地下水深达18 m,由于人工栽植树木根系主要分布在1 m以内,因此地下水难以被利用。土壤以红色的粉砂质黏壤土为主。样地林分主要是以 15年生常绿阔叶乔木樟树(Cinnamomum camphora)和常绿针叶乔木刺杉(Cunninghamia lanceolate)、马尾松(Pinus massoniana)组成的人工混交林,郁闭度为 0.65—0.80,林分密度为 1000 plant·hm−2。虽为常绿混交林,但生长季初期伴随新叶生长出现大量枯叶掉落,地表常年枯落物质堆积厚度可达5 cm。此外,混交林外围分布有稀疏低矮灌木栀子花(Gardenia jasminoides)和野茶花(Camellia japonica),覆盖度在40%左右。
1.2 样品采集
于2017年3月—2019年9月采集植物茎杆和叶片样品。为避免降水干扰,样品采集一般在降水后3—10 d进行,采样频率为每月1—3次,共69次。采集植物样品时,分别选择3棵长势良好的樟树和刺杉作为固定样树,用于长期连续取样,其中,刺杉取样于2018年9月开始。采集枝条样品时,剪取冠层下部已木质化的小枝(直径0.3—0.5 cm,长4—6 cm),并将剪下枝条迅速去皮后装入 10 mL玻璃瓶内,并用Parafilm封口膜密封、编号、冷冻保存,待测定其水稳定同位素比率。采集叶片样时,为避免新生叶和衰老叶造成的同位素差异,一般剪取不同方位的成熟叶片,并将其混合后迅速装入10 mL玻璃瓶,并用Parafilm封口膜密封、编号、冷冻保存,待测定其水稳定同位素比率。每次植物样品采集至少取3个重复,并记录完整的采样时间,以对应相应时刻环境因子的变化。此外,两种植物日内动态变化过程的取样(枝条、叶片)于典型无风晴日的2019年5月23—24日和2019年8月28—29日进行,采样间隔为2 h,采集方式与上面相同。
土壤样品采集与植物样品采集同步进行,在样树附近利用手持式土壤取样钻机(科力SD-1,北京吉奥科技有限公司)钻取土壤样,每次钻取2个平行土样。钻孔深度100 cm,0—20 cm取样间隔为5 cm,20 cm以下取样间隔为10 cm。采集的土壤样品密封于10 mL的玻璃瓶中用于测定土壤水中稳定同位素比率。
降水取样分别在降水日(降水量≥0.1 mm)的北京时间08:00和20:00进行。将采集的降水样直接装入30 mL的聚乙烯样品瓶内,密封、编号且低温保存,待测定其稳定同位素比率。参照气象部门的规范,将当日20:00(代表当日08:00—20:00的累积降水量)和次日08:00(代表当日20:00至次日08:00的累积降水量)收集的降水量之和记为当日的降水量,相应时段的降水中稳定同位素值也记为当日。
1.3 环境因子的观测
土壤体积含水量采用布设在样地中心位置的云智能管式土壤水分温度监测仪(RWET-100,智墒,中国)测定。仪器探头位于0—100 cm每隔10 cm处,监测精度为±2.5%,监测时间自 2017年 2月起,采集频率为每60 min一次。土壤含水量监测数据利用烘干称质量法进行水分校正(王锐等,2020b)。
气温 T/℃、相对湿度 RH/%、太阳辐射RS/(W·m−2)、风速 U/(m·s−1)等气象数据均由设置在实验场地外 50 m处的微型自动气象站(232,WeatherHawk,美国)获取。监测时间自 2016年12月起,监测频率为每30分钟一次。此外,根据监测的T和RH可计算出林地附近的饱和水汽压差VPD/kPa,其计算公式为:
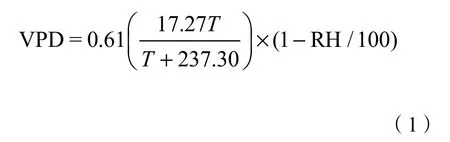
1.4 样品处理与测试
利用全自动真空冷凝抽提系统(LI-2100,北京理加联合有限公司)从土壤、植物茎杆和叶片中抽提出土壤水、植物茎杆水和叶片水。所有水样的氢、氧稳定同位素比率均利用气-液两用型稳定同位素分析仪(DLT-IWA-35EP,美国Los Gatos Research公司)测定。被测试水样中的稳定同位素丰度用相对于维也纳标准平均海洋水(V-SMOW)稳定同位素比率的千分差值(‰)表示:

式中:
Rsample和 RV-SMOW分别代表水样和标准平均海洋水中稳定同位素比率(D/1H或18O/16O)。
在测试植物叶片水中稳定同位素组成时选择正标样以提高测试结果准确度。δD和δ18O的测试精度分别为±0.6‰和±0.2‰。本文中,除降水和土壤水稳定同位素的平均值分别为相应时段内降水量和土壤含水量的加权平均值外,其他所有要素的平均值均指时段内的算术平均值。
由于植物样品在抽提过程中存留有机物,而有机物中的甲醇和乙醇会引起光谱干扰并造成被测定的植物茎杆水和叶片水中稳定同位素值出现偏差,因此需对水样中的醇类污染进行稳定同位素比率的校正(刘文茹等,2013)。由于所选植物水样中仅发现甲醇类污染,所以通过实验建立的甲醇类污染误差校正公式为:

式中:
ΔδD和Δδ18O分别代表由甲醇类污染导致的植物水样中稳定同位素值误差;
NB为窄带系数且随植物类型而变化,该参数主要反映水样中甲醇的污染程度,由 LGR公司的稳定同位素分析处理软件提供。
校正后的稳定同位素值为实测值减去误差值。
1.5 植物叶片水中稳定同位素富集程度
尽管植物根系吸收的水分在植物体内运输的过程中不发生稳定同位素分馏,但是植物蒸腾作用却会使得叶片水中稳定同位素富集(Farquhar et al.,2003)。叶片水中稳定同位素相较于源水(植物茎杆水)中稳定同位素的富集程度用ΔδL来表示,其计算公式为:

式中:
ΔδL表示叶片水中稳定同位素相较于茎杆水中稳定同位素的富集值;
δA代表植物叶片水中δD(或δ18O);
δB代表植物茎杆水中δD(或δ18O)。
1.6 时期的划分
通过查阅相关文献资料,长沙地区雨季主要集中在(3—6月),在7—9月受伏旱影响,降水少,干旱严重;而在10月至次年2月,虽然该时期降水少,但温度低,相对湿度较大,干旱发生频率小(王文等,2016;刘仲藜等,2020)。基于此,将观测期分为湿润期(10月至次年6月)和干旱期(7—9月)两个时段,深入探讨不同水分条件下研究区森林系统内部各水体中水稳定同位素的变化特征。
2 结果与分析
2.1 降水量与土壤含水量变化
图1给出了2017年3月—2019年9月期间日降水量和土壤含水量随时间的变化。根据统计,2017、2018和2019年的雨季(3—6月)降水日数分别为64、38和56 d,累积降水量分别为1095.0、366.0和758.9 mm。各年旱季(7—9月)降水日数分别为21、27和10 d,累积降水量分别为287.0、515.0和157.8 mm。此外,2017年10月至次年2月的总降水日数为52 d,累积降水量为290.9 mm;2018年10月至次年2月期间的总降水日数达83 d,累积降水量为 473.2 mm,较上年同期降水量增加62.7%。

图1 研究区日降水量(P)和土壤含水量(SWC)随时间的变化Fig. 1 Temporal variations of precipitation and soil water content in study area
观测期间土壤含水量在 20.9%—39.9%范围内变化,并随深度的增加逐渐递增。在时间变化上,表层0—10 cm土壤含水量的高值出现在2017年3—6月和2018年10月至次年6月,平均值分别为34.4%和 35.3%,这与两时期偏多的降水有关。在2017年10月至次年6月,由于降水较2017年3—6月和2018年10月至次年6月偏少,表层土壤含水量明显低于两时期,平均值为30.0%。比较发现,在2017、2018和2019年的7—9月,受较少降水补给和较强蒸散发的影响,表层土壤含水量较年内其它时段明显偏低,分别只有 26.7%、27.9%和21.9%,表明该时期研究区受干旱胁迫程度偏重。
2.2 降水和土壤水中δ18O的变化
降水作为重要的水源输入项,其稳定同位素组成直接或间接影响着土壤水和植物水中稳定同位素变化。由图2可知,观测期间降水中δ18O的变化范围为−13.32‰—2.85‰,最大值和最小值的出现的时间分别为4—5月和7—9月。根据表1的统计,降水中稳定同位素组成存在明显的季节差异。在2017年3—6月、2017年10月至次年6月和在2018年10月至次年6月的湿润期,降水中δ18O的平均值±标准差分别为 (−6.69‰±3.45‰)、(−5.29‰±2.74‰)和 (−4.81‰±2.46‰)。相较于湿润期,2017、2018和2019年的干旱期内降水中δ18O明显偏负,平均值±标准差分别为 (−10.38‰± 2.35‰)、(−8.93‰±2.06‰) 和 (−8.64‰±2.19‰)。降水中稳定同位素的季节差异主要与该地区水汽来源的季节变化有关。在湿润期,长沙地区水汽主要来自于西风环流所携带的大陆性气团,该气团具有湿度低,蒸发性强的特点,且冬季降水较少,一般以小雨为主,气团中稳定同位素在运输过程中被贫化程度低,因此长沙地区降水中稳定同位素更富集。在干旱期,来自热带海洋上的气团,在输送过程中受强烈对流作用,降水频繁,且降水量较大,剩余气团中同位素组成逐渐被贫化,因此长沙地区降水中稳定同位素较为贫化(吴华武等,2012)。
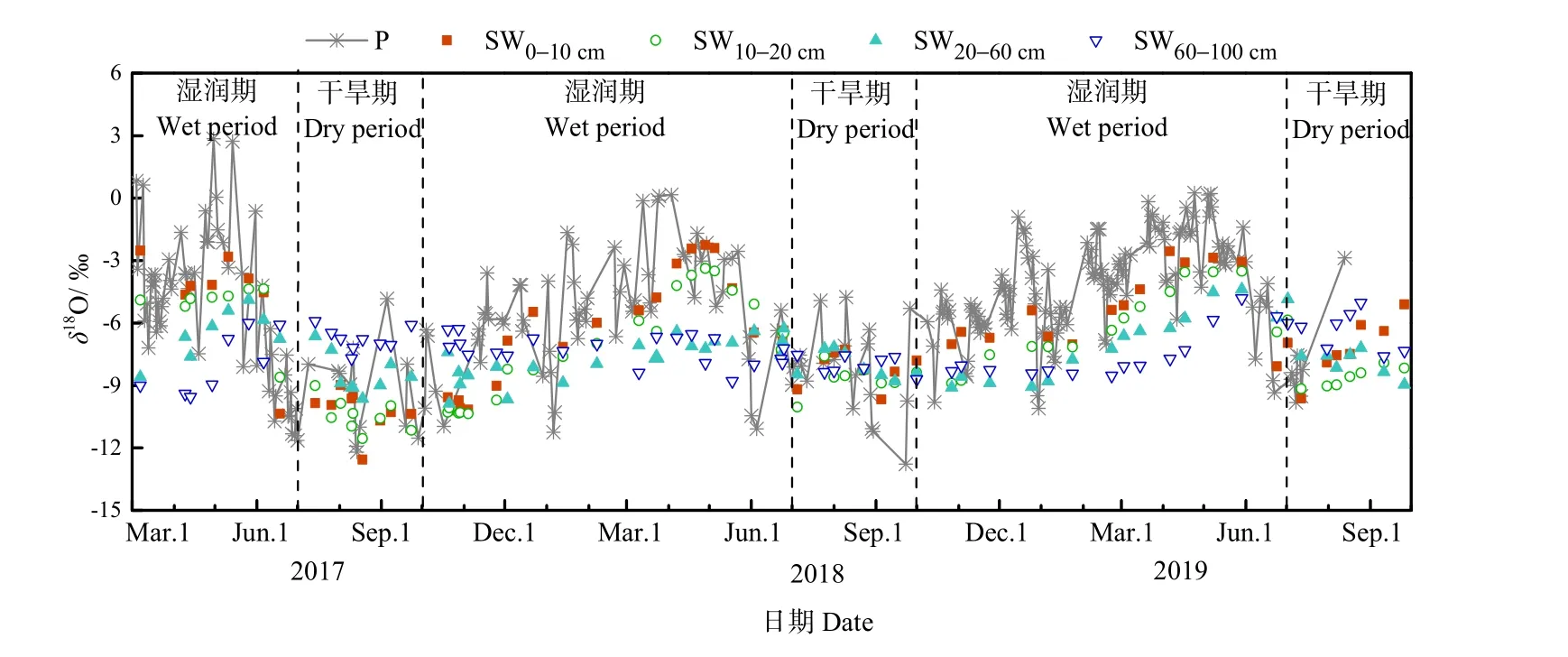
图2 观测期内降水(P)、土壤水(SW)中δ18O随时间的变化Fig. 2 Temporal variations of δ18O of precipitation (P) and soil water (SW) during the observation period

表1 干旱期、湿润期内降水(P)和土壤水(SW)中δ18OTable 1 δ18O in precipitation (P) and soil water (SW) in wet and dry periods ‰
为进一步分析不同深度土壤水变化特征和确定植物水分利用特征,根据不同深度土壤含水量和土壤水中稳定同位素组成的特点,将土壤含水量和稳定同位素组成相近的相邻土层合并为同一层次,因此将0—100 cm土壤剖面分为0—10、10—20、20—60和60—100 cm 4个层次。土壤水中δ18O的变化范围在表层 0—10 cm 最大,为−12.56‰—−2.25‰,并随深度的增加逐渐减小。可以看到,0—10、10—20、20—60 cm土壤水中δ18O达到最大值和最小值的时间与降水一致,分别为4—5月和7—9月。而60—100 cm土壤水中δ18O达到最大值和最小值的时间较降水存在滞后,分别为 6—7月和9—10月。这表明0—60 cm土壤水受降水直接补给影响较大,而60—100 cm深层土壤水对降水响应存在1—2个月的滞后。
与降水中稳定同位素变化相似,土壤水中δ18O存在明显季节变化,且随深度的增加季节变化逐渐减小(表1)。在2017年3—6月、2017年10月至次年6月和2018年10月至次年6月的湿润期,表层0—10 cm土壤水中δ18O明显偏正,平均值±标准差分别为 (−4.64‰±2.43‰)、(−6.48‰±2.65‰)和 (−5.44‰±1.87‰),且随深度的增加土壤水中δ18O逐渐偏负。在2017、2018和2019年的干旱期,表层土壤水中δ18O较各湿润期偏负,平均值±标准差分别为 (−10.19‰±1.02‰)、(−8.28‰±0.90‰)和(−7.14‰±1.35‰),随深度的增加土壤水中δ18O 整体呈偏正的趋势。不同时期土壤水中δ18O随深度的变化主要与降水在土壤剖面中的补给过程有关,降水优先补给浅层土壤水,在浅层土壤水达到饱和后推动前期土壤水向下逐层补给深层土壤水。
2.3 植物水中δ18O变化
2.3.1 植物水中δ18O的季节变化
图3给出了取样期间樟树和刺杉茎杆水和叶片水中δ18O随时间的变化,表2则统计了各湿润期和各干旱期内两种植物茎杆水、叶片水中δ18O和叶片水稳定同位素富集程度Δ18OL的平均值。

图3 樟树和刺杉茎杆水(XW)、叶片水(LW)中δ18O随时间的变化Fig. 3 Temporal variations of δ18O of xylem water (XW) and leaf water (LW) of C. camphora and C. lanceolate
取样期间樟树和刺杉茎杆水中δ18O变化范围分别为−9.40‰— −3.30‰和−8.35‰— −3.73‰。与降水相似,两种植物茎杆水中δ18O的最大值和最小值分别出现在4月和8月。时间变化上,在2017年3—6月、2017年10月至次年6月和2018年10月至次年6月的湿润期,樟树茎杆水中δ18O明显偏正,平均值±标准差分别为 (−4.59‰±0.43‰)、(−6.99‰±1.49‰)和 (−5.66‰±1.54‰);而在 2017、2018和2019的干旱期,樟树茎杆水中δ18O较各湿润期偏负,平均值±标准差分别为 (−7.84‰±0.85‰)、(−7.39‰±0.58‰)和 (−6.34‰±0.37‰)。比较发现,刺杉茎杆水中δ18O在2018年10月至次年6月的湿润期和在2019年7—9月的干旱期均与樟树茎杆水中δ18O接近,平均值±标准差分别为(−5.72‰±1.36‰)和 (−5.73‰±0.73‰),这可能与两时期内樟树与刺杉水分利用来源相似有关。结合表1中不同时期4层土壤水中δ18O比较发现,在各湿润期,两种植物茎杆水中δ18O与0—20 cm土壤水中δ18O接近;而在各干旱期,两种植物茎杆水中δ18O则主要与20—60 cm和60—100 cm的土壤水中δ18O相似。这表明两种植物在各湿润期主要利用0—20 cm的浅层土壤水,而在各干旱期则主要利用20—60 cm和60—100 cm的土壤水。
相较于茎杆水,叶片水中稳定同位素明显富集,且变幅更大。樟树和刺杉叶片水中δ18O的变化范围分别为−4.32‰—20.29‰和−1.60‰—15.84‰。由表2可知,各湿润期樟树和刺杉叶片水中δ18O均较各干旱期偏正,两种植物叶片水Δ18OL也在湿润期大于干旱期。根据Craig et al.(1961)对大气水线的定义,将植物叶片水中δD和δ18O之间的线性关系定义为叶片水线(LWL)。研究表明,LWL斜率的大小可指示植物叶片水中同位素动力分馏程度的强弱,斜率越小表示叶片水中稳定同位素组成受动力分馏作用越强(Yakir et al.,1990;Yepez et al.,2003)。结合表3,湿润期内樟树和刺杉 LWL 分别为δD=3.06δ18O−21.74(R2=0.68)和δD=2.44δ18O−16.84(R2=0.63)。干旱期内两种植物LWL 分别为δD=3.26δ18O−39.70(R2=0.67)和δD=2.61δ18O−27.72(R2=0.71),其斜率均大于湿润期内两种植物 LWL斜率。从时间变化上看,无论是樟树还是刺杉,叶片水中稳定同位素的富集程度和动力分馏程度均在湿润期大于干旱期。在湿润期,土壤水分充足,两种植物采取挥霍型水分利用策略,植物叶片水与大气水汽交换频繁。在干旱期,由于土壤水分亏缺严重,两种植物用水趋于保守,植物通常选择关闭部分气孔减少蒸腾耗水,植物叶片水与大气水汽交换受阻(罗紫东等,2016)。因此,在湿润期,樟树和刺杉叶片水中δ18O、Δ18OL较干旱期大,而LWL斜率则明显小于干旱期。

表2 干旱期、湿润期内樟树与刺杉茎杆水中δ18O、叶片水中δ18O和Δ18OLTable 2 δ18O in xylem water, δ18O and Δ18OL in leaf water of C. camphora and C. lanceolata in dry and wet periods ‰

表3 不同时间尺度上樟树和刺杉叶片水线方程(LWL)Table 3 Leaf water line (LWL) of C. camphora and C. lanceolata on different time scales
2.3.2 植物水中δ18O的日内变化
图4给出了湿润期和干旱期典型晴日内两种植物茎杆水、叶片水中δ18O的逐时变化。在所选湿润期的典型晴日,樟树和刺杉茎杆水中δ18O的平均值±标准差分别为 (−3.58‰±0.31‰)和 (−3.80‰±0.32‰);在所选干旱期的典型晴日,两植物茎杆水中 δ18O 平均值±标准差分别为 (−6.09‰±0.39‰)和(−6.63‰±0.34‰)。可以看出,樟树和刺杉茎杆水中δ18O均无明显的日内变化,这说明两种植物水分利用来源相对稳定。

图4 湿润期和干旱期典型晴日内樟树和刺杉茎杆水(XW)、叶片水(LW)中δ18O的逐时变化Fig. 4 Hourly variation of δ18O in xylem water (XW) and leaf water (LW) of C. camphora and C. lanceolata on typical sunny days during wet and dry periods
相较于茎杆水,樟树和刺杉叶片水中δ18O存在明显的日内变化。在湿润期和干旱期的典型晴日,樟树叶片水中 δ18O 分别在 9.90‰—16.25‰和−1.68‰—12.30‰内变化,刺杉叶片水中δ18O的变化范围则分别为 9.07‰—13.42‰和−0.58‰—10.01‰。在时间变化上,两种植物叶片水中 δ18O最大值均出现在16:00—18:00,而最小值则出现在06:00—08:00。与叶片水中δ18O对应,两种植物叶片水Δ18OL均在16:00—18:00最大,在06:00—08:00最小。
两时期典型晴日,樟树和刺杉叶片水中稳定同位素组成存在明显差异。在湿润期典型晴日樟树和刺杉叶片水中 δ18O偏正,平均值±标准差分别为(12.54‰±2.35‰)和 (10.77‰±1.56‰),对应的两种植物叶片水 Δ18OL分别为 (16.12‰±2.60‰)和(14.57‰±1.77‰)。在干旱期典型晴日,樟树和刺杉叶片水中 δ18O偏负,平均值±标准差分别为(6.58‰±4.26‰)和 (5.26‰±3.06‰),对应的叶片水Δ18OL也小于湿润期典型晴日,平均值±标准差分别为 (12.68‰±4.35‰)和 (12.20‰±3.25‰)。结合表3,樟树和刺杉 LWL在湿润期典型晴日分别为δD=1.05δ18O+17.24(R2=0.66)和 δD=2.17δ18O+1.03(R2=0.79)。在干旱期典型晴日,两种植物LWL分别为 δD=1.94δ18O−29.48(R2=0.94)和 δD=2.44δ18O−27.71(R2=0.92),其斜率大于湿润期典型晴日两种植物 LWL的斜率。上述结果表明,两种植物叶片水中稳定同位素富集程度和动力分馏程度在湿润期典型晴日更大,而在干旱期典型晴日更小,这可能与两晴日内植物茎杆水中 δ18O大小及其微气象环境有关。在湿润期典型晴日,由于两种植物叶片水水源(茎杆水)中δ18O较干旱期典型晴日明显偏正,在相同的蒸腾作用下,两种植物叶片水中δ18O 更富集。Butt et al.(2010)和 Lai et al.(2008)研究发现,在暖湿的环境中,植物叶片水内外水汽压差较小,不利于叶片水与大气进行水汽交换,植物叶片水中稳定同位素的分馏程度较弱。湿润期典型晴日T、RH和VPD平均值分别为23.8 ℃、49.5%和1.5 kPa;而干旱期典型晴日,T、RH和VPD均高于湿润期,平均值分别为 31.9 ℃、61.5%和 1.8 kPa,因此两种植物叶片水中稳定同位素的富集程度和动力分馏程度在湿润期典型晴日更大。
2.4 影响植物Δ18OL变化的气象因子
植物叶片水中稳定同位素富集变化过程主要受区域微生境的气象条件的影响(罗伦等,2013)。表 4给出了不同时间尺度上樟树和刺杉叶片水Δ18OL与各气象因子(温度T、相对湿度RH、太阳辐射RS、饱水汽压差VPD)相关系数。

表4 不同时间尺度下樟树和刺杉Δ18OL与各气象因子的相关关系Table 4 Correlation between leaf water Δ18OL and meteorological factors of C. camphora and C. lanceolata in different time scales
在季节变化上,无论是湿润期还是干旱期,樟树和刺杉叶片水 Δ18OL与 T均无显著相关关系(P>0.05),与RH和VPD分别呈极显著的负相关和正相关关系(P<0.01)。此外,在湿润期,两植物叶片水Δ18OL与RS表现出极显著正相关;在干旱期,樟树叶片水Δ18OL与RS呈显著的正相关,刺杉由于观测样本量较少,叶片水中Δ18OL与RS表现出不显著的正相关。这在一定程度上说明,在年内两种植物叶片水中稳定同位素富集程度主要受 RH、RS和VPD影响,而受T的影响较小。在湿润期和干旱期的典型晴日内,樟树和刺杉叶片水Δ18OL与T均表现出显著的正相关(P<0.05),与RH和VPD分别表现出呈极显著负相关和正相关。此外,在两时期典型晴日内,两种植物叶片水Δ18OL与RS均无显著相关关系。这表明在日内逐时变化中,T、RH和 VPD是影响两种植物叶片水中稳定同位素富集程度的主要因子,而RS对两种植物叶片水中稳定同位素富集程度的影响较小。
RH和VPD控制着植物蒸腾速率和气孔导度的变化,进而影响着植物叶片水Δ18OL(Farquhar et al.,2003;Butt et al.,2010;Barbour et al.,2000)。即当RH较大时,VPD则越小,叶片内外水汽压接近,气孔导度减小,植物蒸腾作用减弱,叶片水与冠层水汽的交换减弱,叶片水中稳定同位素富集程度低;反之,叶片水中稳定同位素富集程度低较大。因此,无论是季节变化还是日内变化,两种植物叶片水Δ18OL与RH和VPD相关性极为显著。
RS通过影响植物光合作用,进而影响叶片水中稳定同位素分馏。植物进行光合作用时,通过气孔进入叶片的一部分 CO2在与水合成葡萄糖的过程中,羰基氧与H2O分子中氧原子发生交换,使得叶片水氧同位素发生分馏(Guy et al.,1993);而另一部分CO2与叶片水在碳酸酐酶催化下,经过稳定同位素平衡反应后返回大气,叶片水中氧同位素同样发生分馏(罗伦等,2013)。因此在季节变化上,樟树和刺杉叶片水Δ18OL与RS的相关性显著;而在日内变化中,由于夜间RS几乎接近0,但两种植物叶片水 Δ18OL在夜间仍较大,故两种植物叶片水Δ18OL与RS相关性不显著。
除RH、VPD和RS影响外,樟树和刺杉叶片水中Δ18OL变化还与T有关。T通过调节冠层温度改变叶片水中稳定同位素的分馏过程,一般情况下,温度越高时,蒸发越强,剩余水体中稳定同位素越富集(Yang et al.,2015;Hua et al.,2019;Dai et al.,2020)。基于该原理,在日内变化中,两植物叶片水中Δ18OL随T的升高而逐渐增大。在季节变化中,各次取样主要在天气晴朗的早上 08:00—09:00,湿润期(或干旱期)内取样时刻的T值较为接近,因此可能导致Δ18OL与T的相关性不显著。
3 结论
受水汽来源的季节变化影响,研究区降水中δ18O表现出在湿润期(10月至次年6月)偏正,在干旱期(7—9月)偏负的季节变化。表层土壤受降水直接补给,土壤水中δ18O与降水中δ18O的季节变化趋势一致,且随深度的增加土壤水中δ18O季节变化逐渐减小。在湿润期,由于樟树和刺杉主要利用的0—20 cm土壤水,茎杆水中δ18O偏正;在干旱期,两种植物主要利用的20 cm以下土壤水且该层土壤水中δ18O偏负,因此茎杆水中δ18O较湿润期偏负。与茎杆水中δ18O较大的季节变化不同,在日内两种植物茎杆水中δ18O均不存在明显变化。
季节变化上,在环境水分充足时期,植物挥霍型水分利用策略加强了植物叶片水与大气的水汽交换,使得叶片水中稳定同位素富集程度和强动力分馏程度加大,即叶片水中δ18O和Δ18OL值更高。而在水分亏缺严重时期,植物通过关闭部分气孔减弱蒸腾耗水,使得叶片水中稳定同位素富集程度和动力分馏程度变小,即叶片水中δ18O和Δ18OL值明显更小。日内变化上,樟树和刺杉叶片水中δ18O和Δ18OL均在16:00—18:00达到最大,在06:00—08:00最小。相较于暖湿的天气(干旱期典型晴日),在干燥的天气(湿润期典型晴日)植物叶片水中稳定同位素富集程度和动力分馏程度更大,即植物叶片水中δ18O和Δ18OL更大。
T、RH、VPD和 RS分别通过调节植物冠层温度、叶片气孔导度、蒸腾速率以及光合速率等方式影响植物蒸腾过程,使得植物叶片水中稳定同位素发生明显分馏。不同时间尺度上影响植物叶片水中稳定同位素富集变化的气象条件存在差异。在季节变化上,两种植物叶片水中δ18O的富集程度主要受RH、RS和VPD的影响;在日内变化上,影响两种植物叶片水 δ18O富集程度的主要气象因子分别为T、RH和VPD。
猜你喜欢
新作文·小学低年级版(2022年4期)2022-08-30 07:23:00
辽河(2021年12期)2021-12-24 00:39:04
国画家(2019年5期)2019-12-31 01:27:16
北方音乐(2019年6期)2019-07-16 07:50:53
现代园艺(2018年3期)2018-02-10 05:18:14
读者(2017年11期)2017-05-16 16:36:18
三联生活周刊(2016年35期)2016-08-29 17:42:03
黄河之声(2016年24期)2016-02-03 09:01:52
黄河之声(2016年24期)2016-02-03 09:01:52
天然产物研究与开发(2014年6期)2014-04-27 14:15:59
