
高粱过敏性诱导反应基因家族HIR的鉴定及表达分析
2021-09-02 09:23方远鹏杜巧丽蒋君梅任明见李向阳田山君
中国粮油学报 2021年7期
陈 俊 方远鹏 杜巧丽 蒋君梅任明见 李向阳 田山君 谢 鑫,
(贵州大学农学院农业微生物特色重点实验室1,贵阳 550025)(国家小麦改良中心贵州分中心2,贵阳 550025)(贵州大学绿色农药与农业生物工程教育部重点实验室3,贵阳 550025)
高粱(Sorghumbicolor)是我国主要的杂粮作物之一,属于C4植物,具有光合作用效率高、抗旱性强等特点[1-3]。同时,高粱作为白酒原材料具有重要的经济价值。白酒是贵州省重要的支柱产业,白酒行业的发展带动了贵州省的高粱种植,其种植面积在贵州省逐年扩大,使得高粱连作现象较为普遍,由于病原的初始菌量积累和气候变化等因素,致使其病害危害程度逐年上升,病害危害面积逐年扩大。
病原菌侵染植物时会触发植物体内一系列的抗病反应[4],即诱导植物防御体系激发抗病基因的表达来应对病原菌的侵入。而过敏性诱导反应是众多植物抗病反应中最为常见的类型,常表现为病原菌侵染点附近的细胞组织坏死,阻止病菌的扩展和生长,诱发植物抗病反应[5]。过敏性诱导反应(Hypersensitive-induced reaction,HIR)基因家族是一类与植物HR相联系的基因家族,隶属PID(proliferation, ion and death,PID)家族,含有SPFH (stomatins, prohibitins, flotillins and HflK/C) 结构域[6,7]。Karrer等[8]最早在烟草中发现一种能够诱导β-葡萄糖苷酶的表达及植物叶片细胞坏死的蛋白NG1。随后,在玉米、大麦、水稻和辣椒等植物中HIR基因相继被报道[9-12]。HIR基因家族成员在植物抗病反应中承担着重要的角色。研究表明,CaHIR1基因是过敏性细胞坏死的正向调控因子,并且能够参与渗透胁迫的反应[13]。同时,HIR基因也能够参与单子叶和双子叶植物的抗病功能和HR反应[7]。目前,HIR基因家族在多种经济作物上已有研究,但在旱粮作物高粱上的影响鲜有报道。
本研究在全基因组的水平上对高粱HIR基因家族成员进行鉴定,并分析其成员的基本理化性质、染色体定位、基因结构、系统进化关系、以及所编码蛋白的二级结构进行分析。同时,利用qRT-PCR技术分析高粱HIR基因家族成员的组织特异性,脱落酸(Abscisic acid,ABA)、赤霉素(Gibberellin A3,GA3)和flg22胁迫时对其表达量的影响,以期为进一步挖掘高粱HIR基因家族成员在植物信号通路、调控植物HR反应等方面的作用提供参考。
1 材料与方法
1.1 材料
高粱品种:BTx623。脱落酸;赤霉素;flg22;SYBR荧光染料试剂盒;TRIzol;反转录试剂盒。
CFX96TMoptics module实时荧光定量分析仪,NanoDrop one微量核酸蛋白浓度测定仪,Centrifuge 5427 R低温离心机。
引物采用Primer Premier 5软件和qPrimerDB数据库相结合进行设计,选用SbEIF4a作为高粱的内参基因,引物序列如表1。

表1 qRT-PCR引物序列
1.2 方法
1.2.1 高粱HIR基因家族鉴定、染色体定位和理化性质分析
从NCBI(https://www.ncbi.nlm.nih.gov/)中下载拟南芥、水稻、玉米、大麦、小麦和辣椒HIR家族成员的氨基酸序列,运用Hmmer程序建立模型,检索高粱数据库,选取E值小于0.05的高粱基因序列;结合NCBI-CD结构域数据库进一步验证E值小于0.05的高粱基因序列的结构域;在phytozome(https://phytozome.jgi.doe.gov/pz/portal.html#)数据库中查询高粱第2、3、7、9和10号染色体的全长,及高粱HIR家族成员所在染色体数和位置区间;利用MG2C(http://mg2c.iask.in/mg2c_v2.0/)在线网站绘制高粱HIR基因家族染色体分布情况;利用在线网站ExPASy-Protparam(https://web.expasy.org/protparam/)分析高粱HIR家族成员的基本理化性质[3];使用Cell-PLoc 2.0(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)在线网站预测高粱HIR家族成员的亚细胞定位。
1.2.2 高粱HIR家族成员系统发育分析
为研究高粱HIR家族成员的进化模式,在NCBI(https://www.ncbi.nlm.nih.gov/)数据库中下载拟南芥、水稻、玉米、大麦、小麦和辣椒的HIR蛋白序列;利用MEGA7.0,以邻接法,bootstrap值设置为1 000[14],构建高粱与这些物种的系统进化树;利用Figtree软件对系统发育树进行美化。
1.2.3HIR家族基因密码子偏好分析
利用在线工具Chips(http://www.bioinformatics.nl/cgi-bin/emboss/chips)和Cusp(http://www.bioinformatics.nl/cgi-bin/emboss/cusp)在线网站分析拟南芥、水稻、玉米、小麦、大麦、辣椒和高粱家族基因密码子的偏好性。
1.2.4 高粱HIR家族基因结构和HIR家族成员保守基序分析
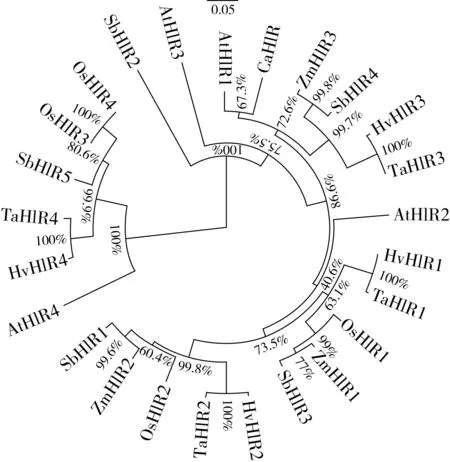
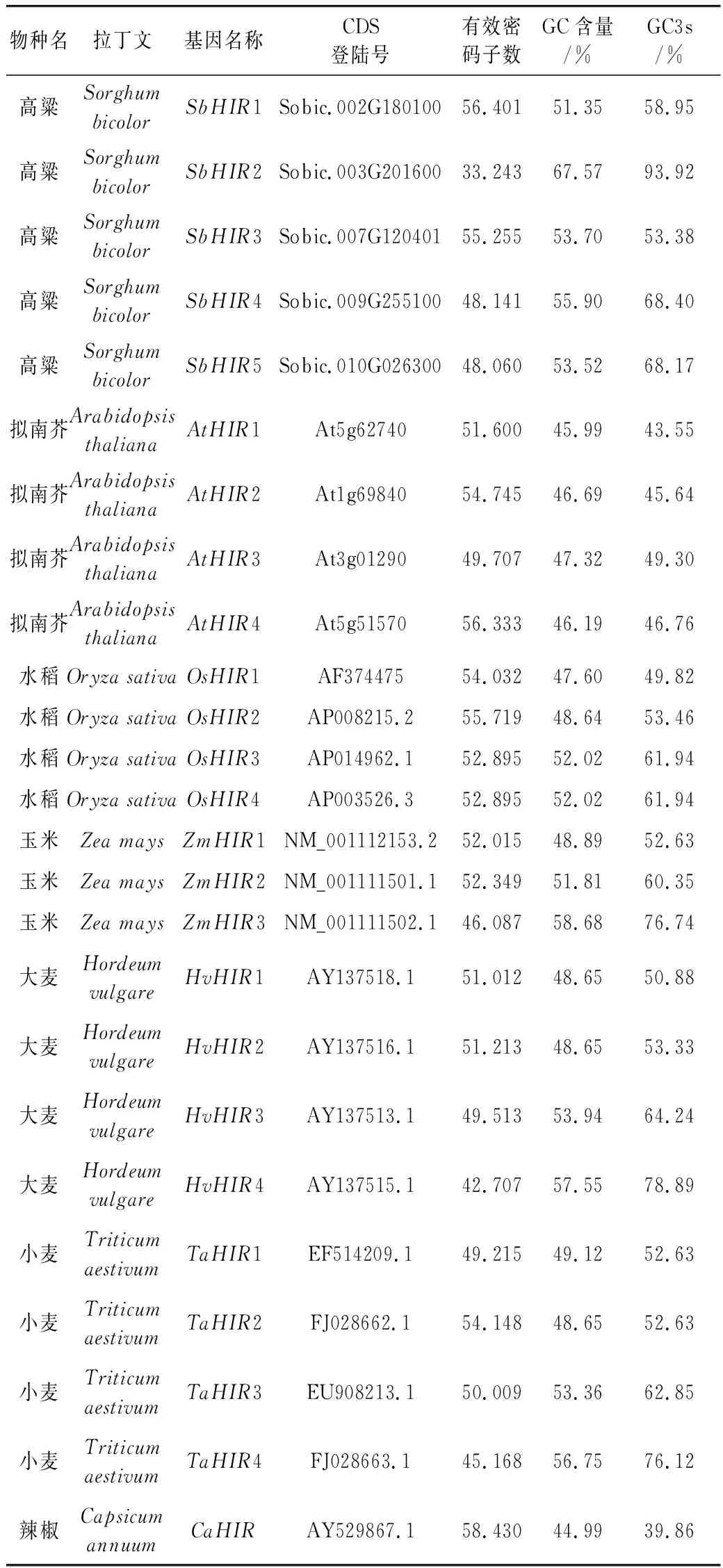
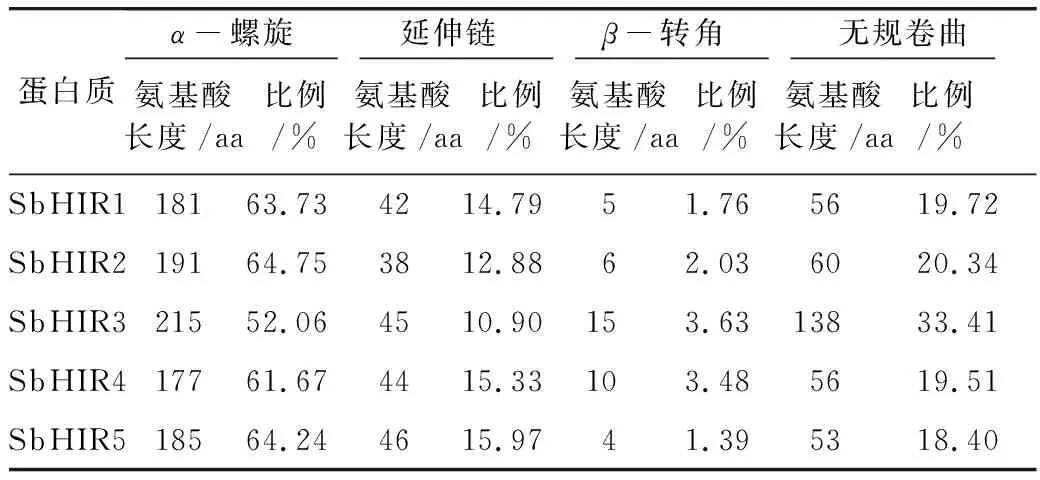
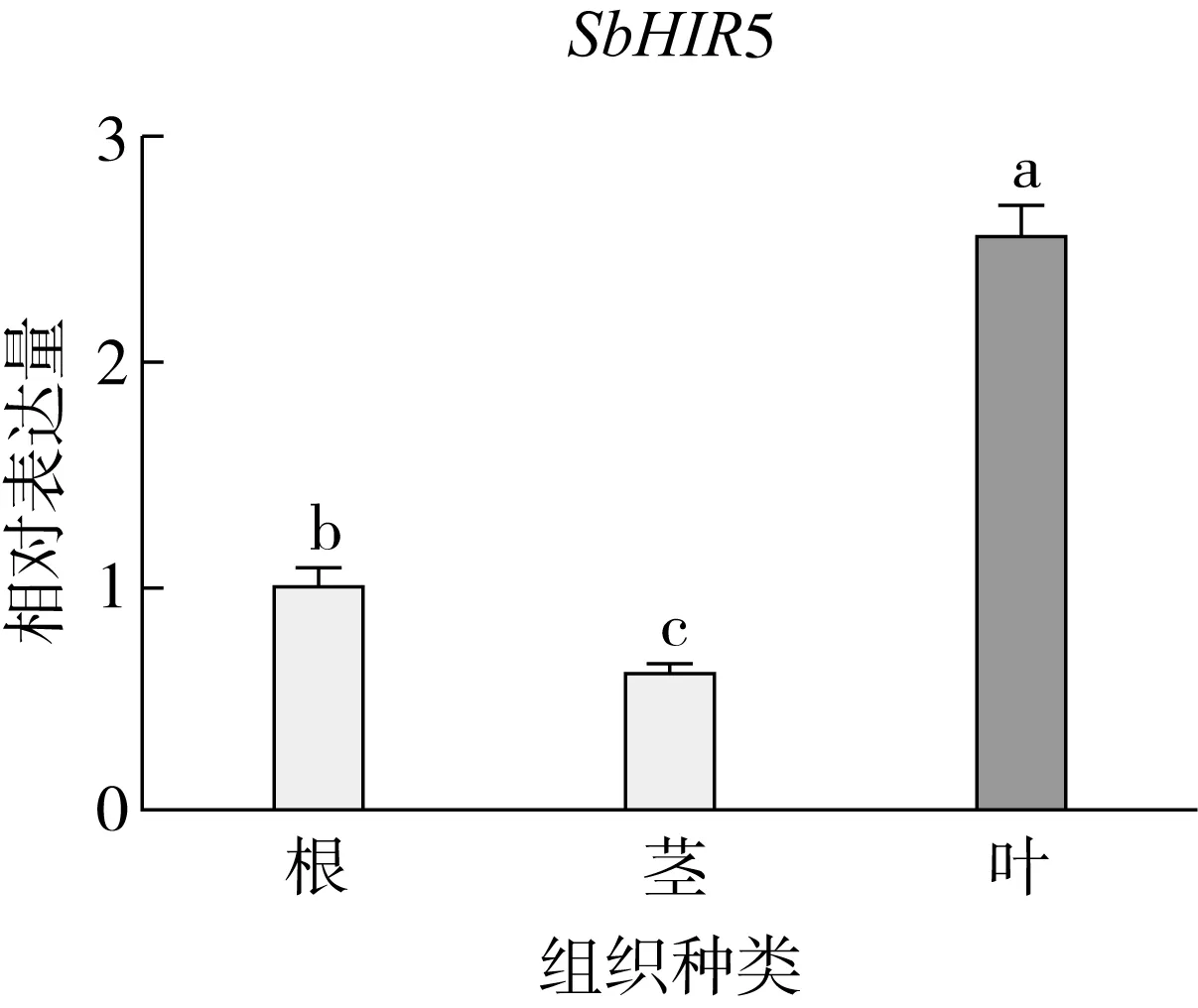
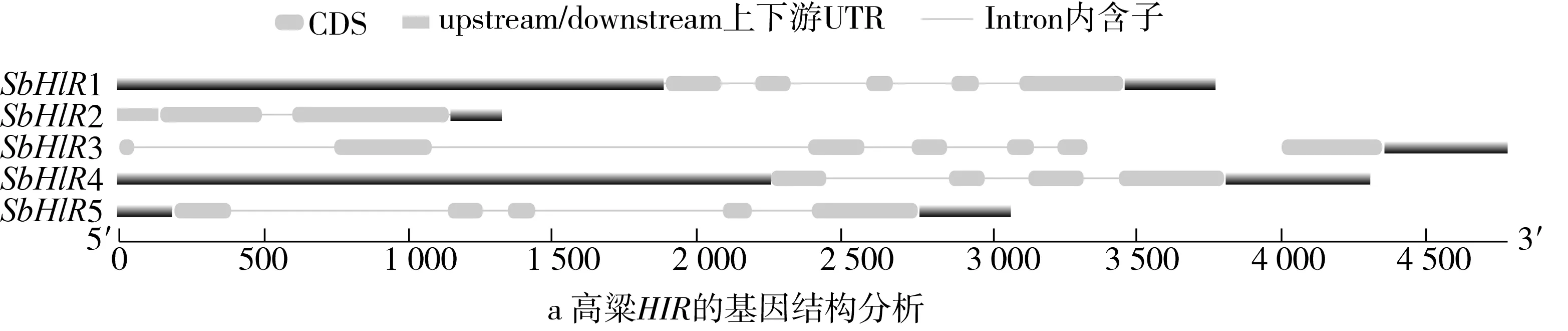
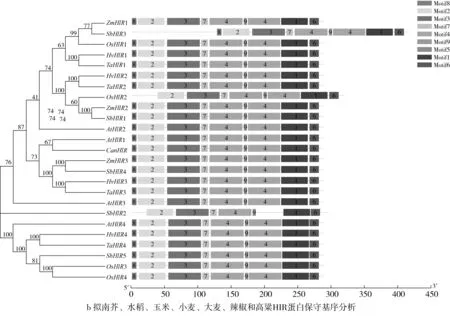
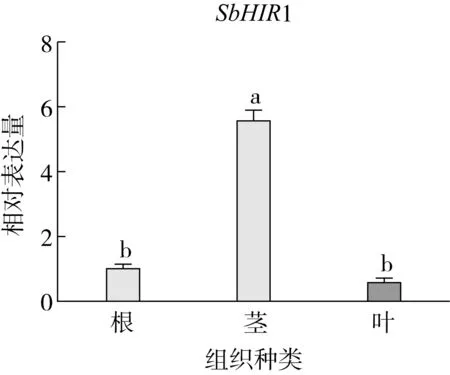
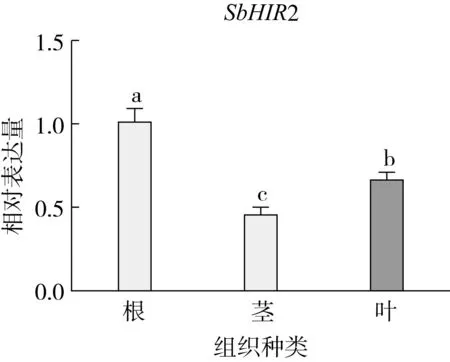
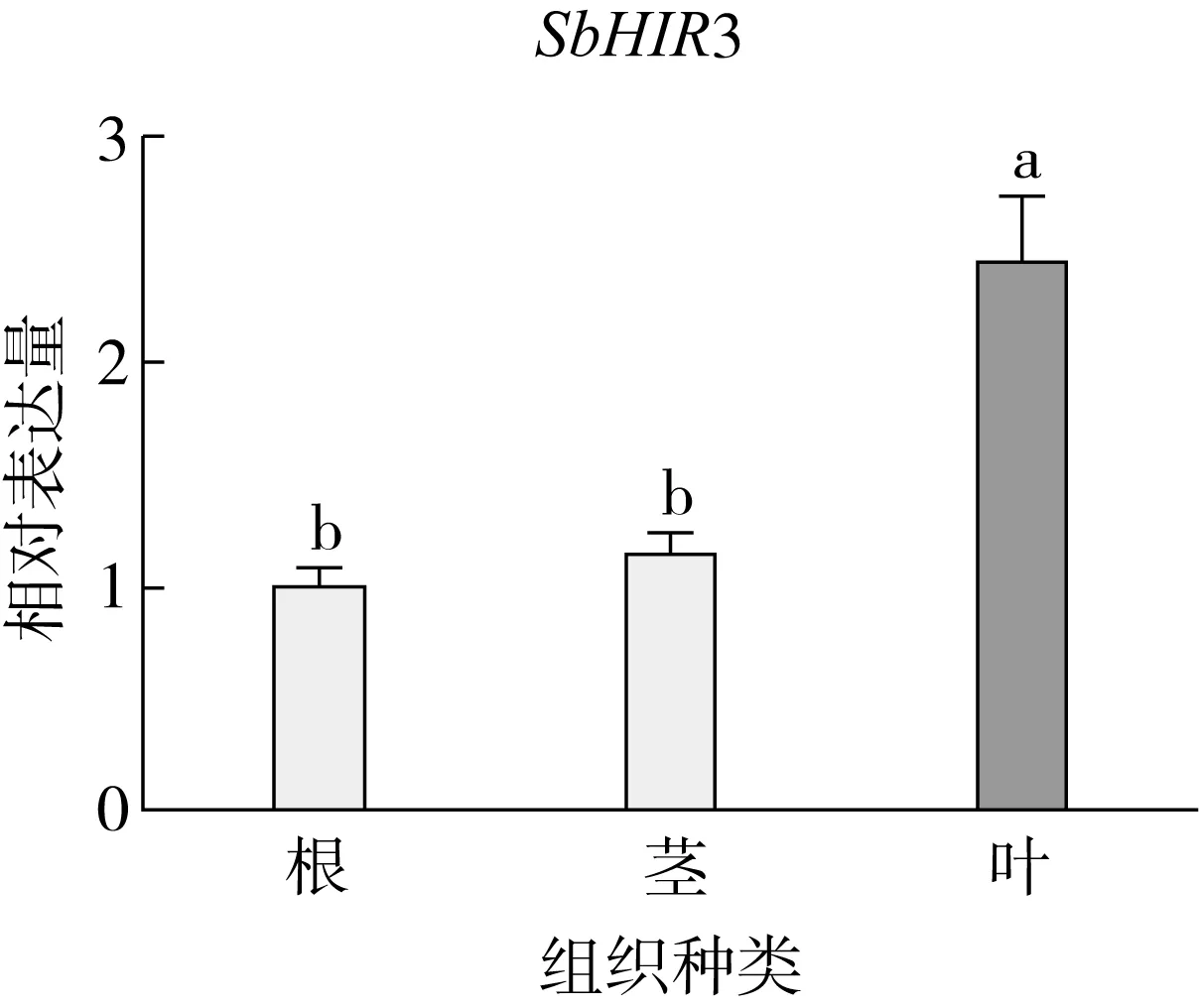
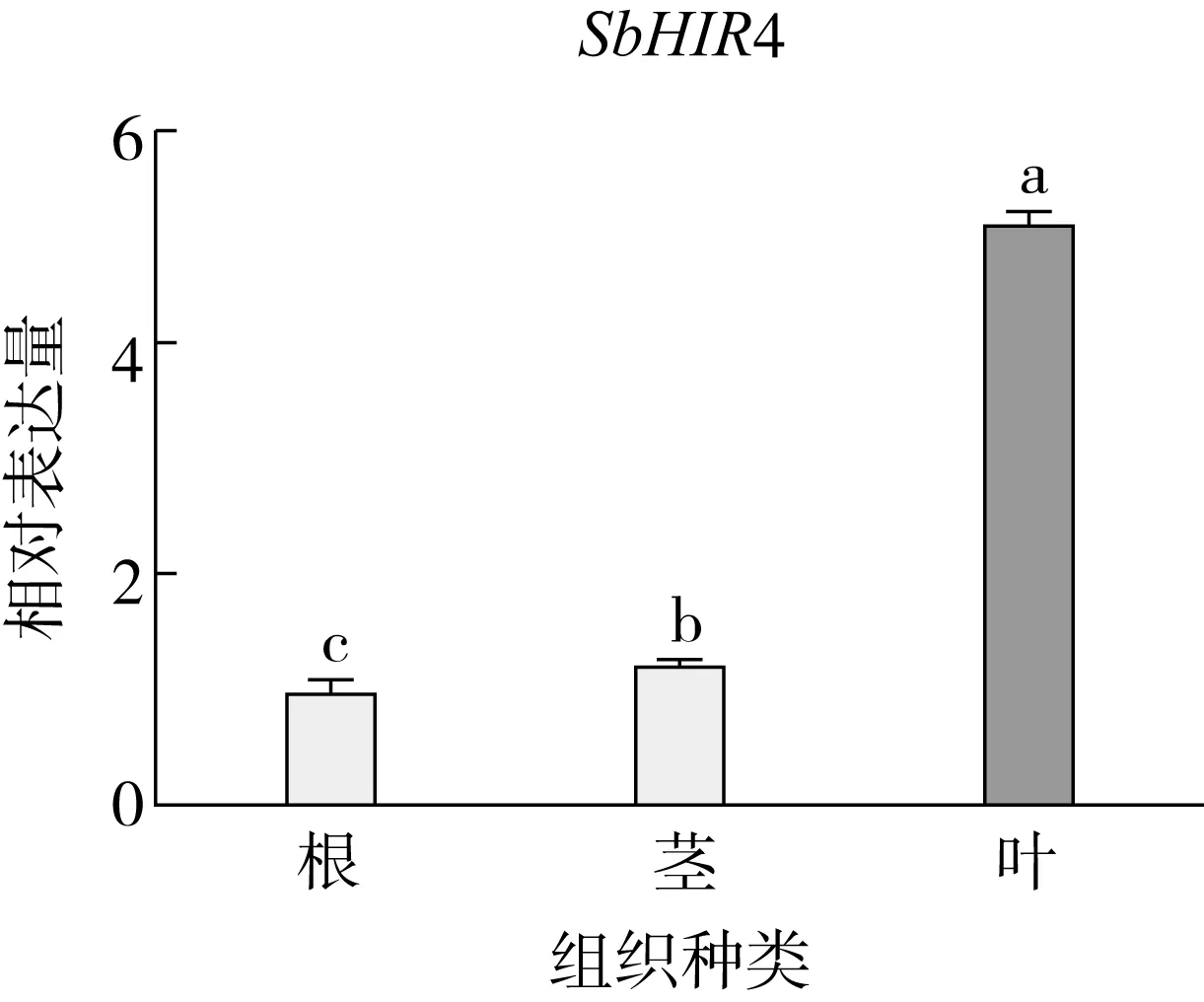
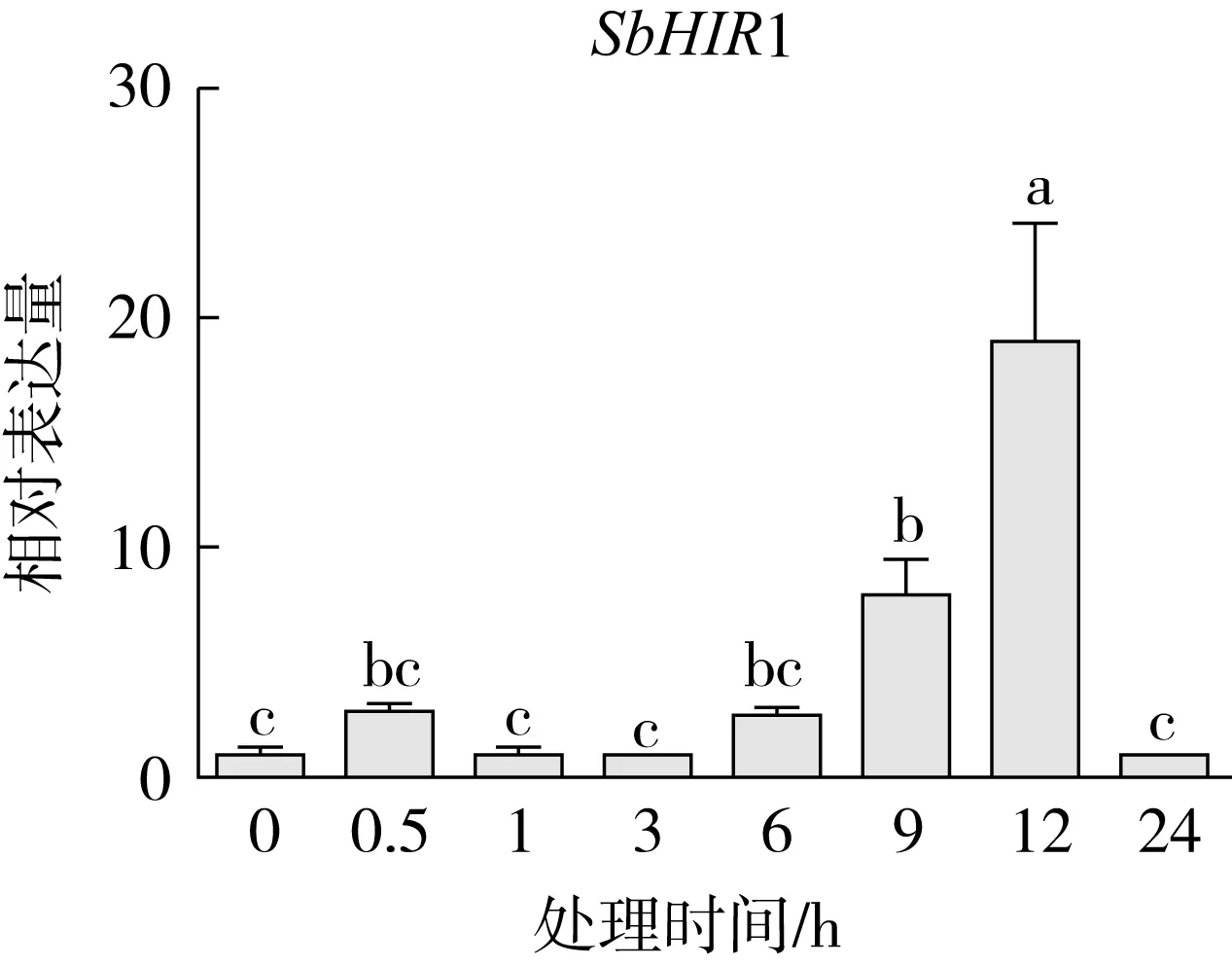
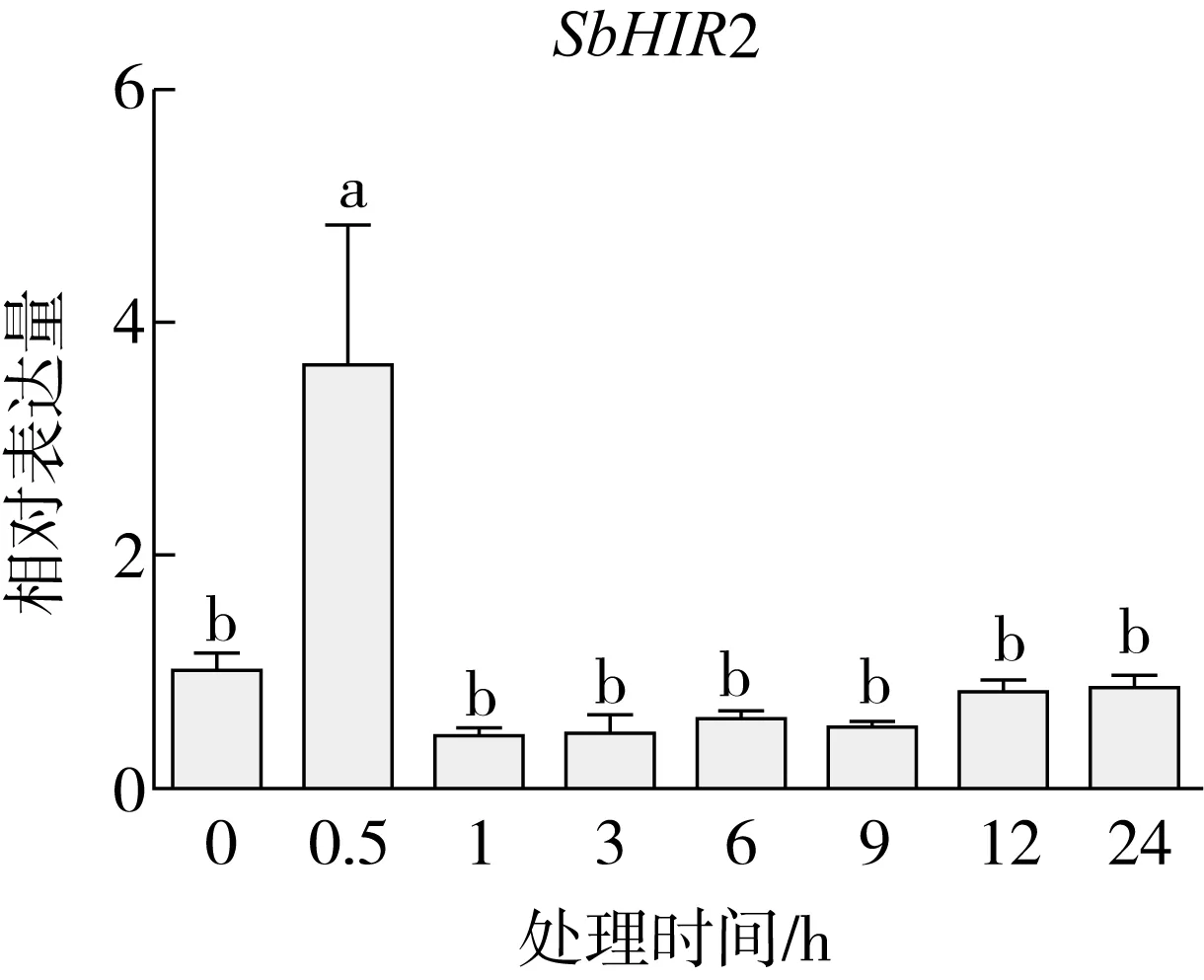
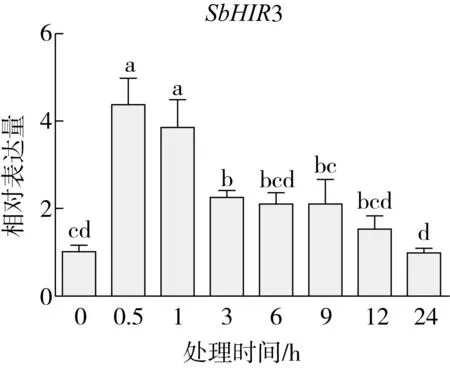
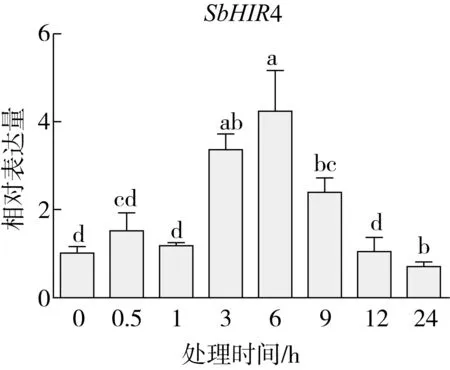
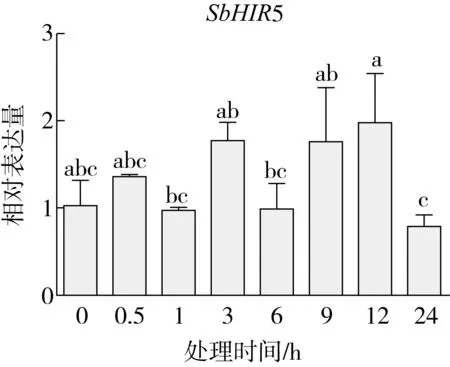
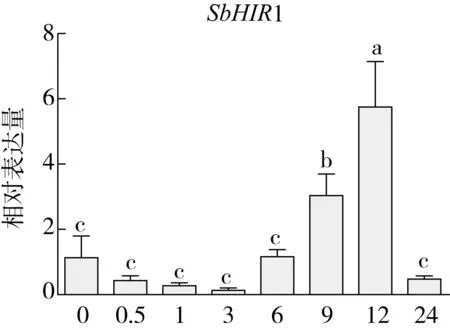
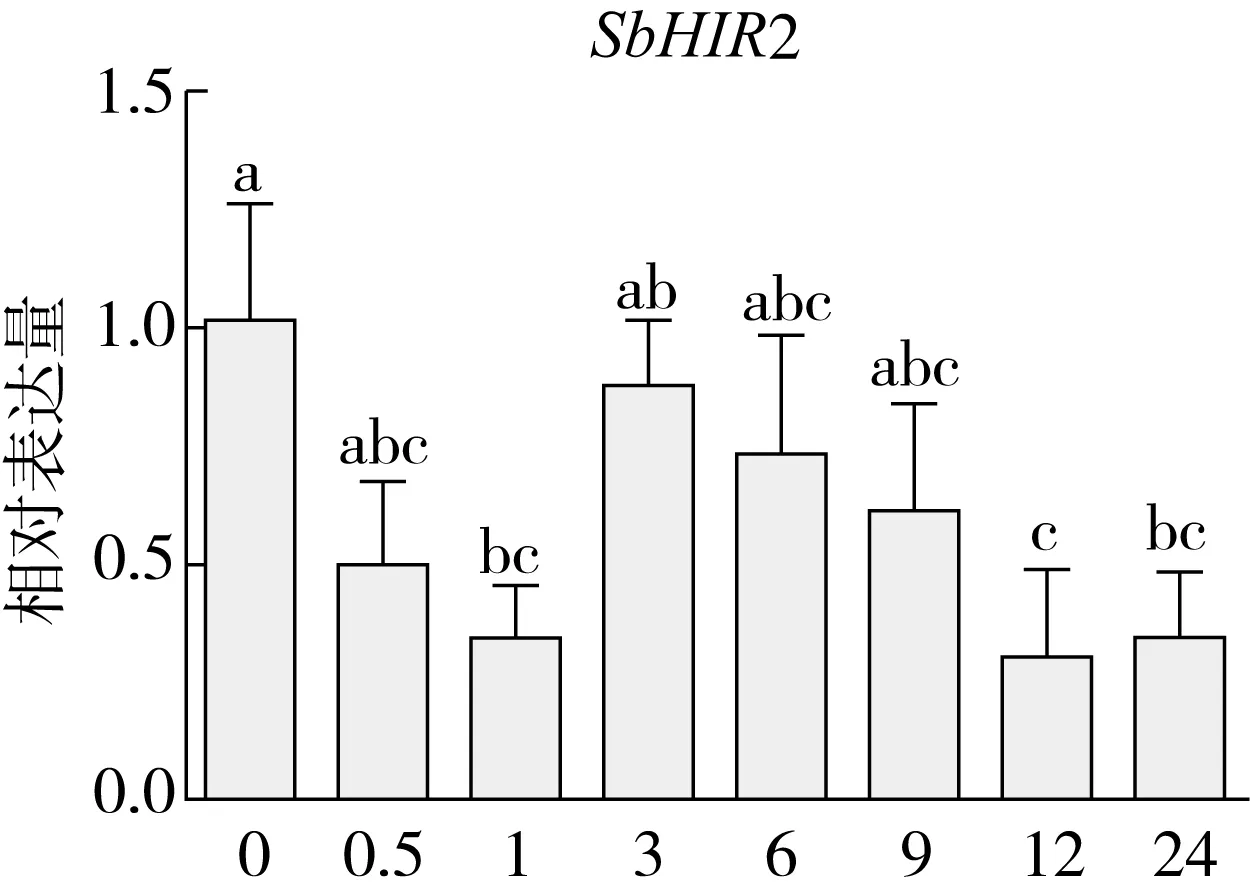
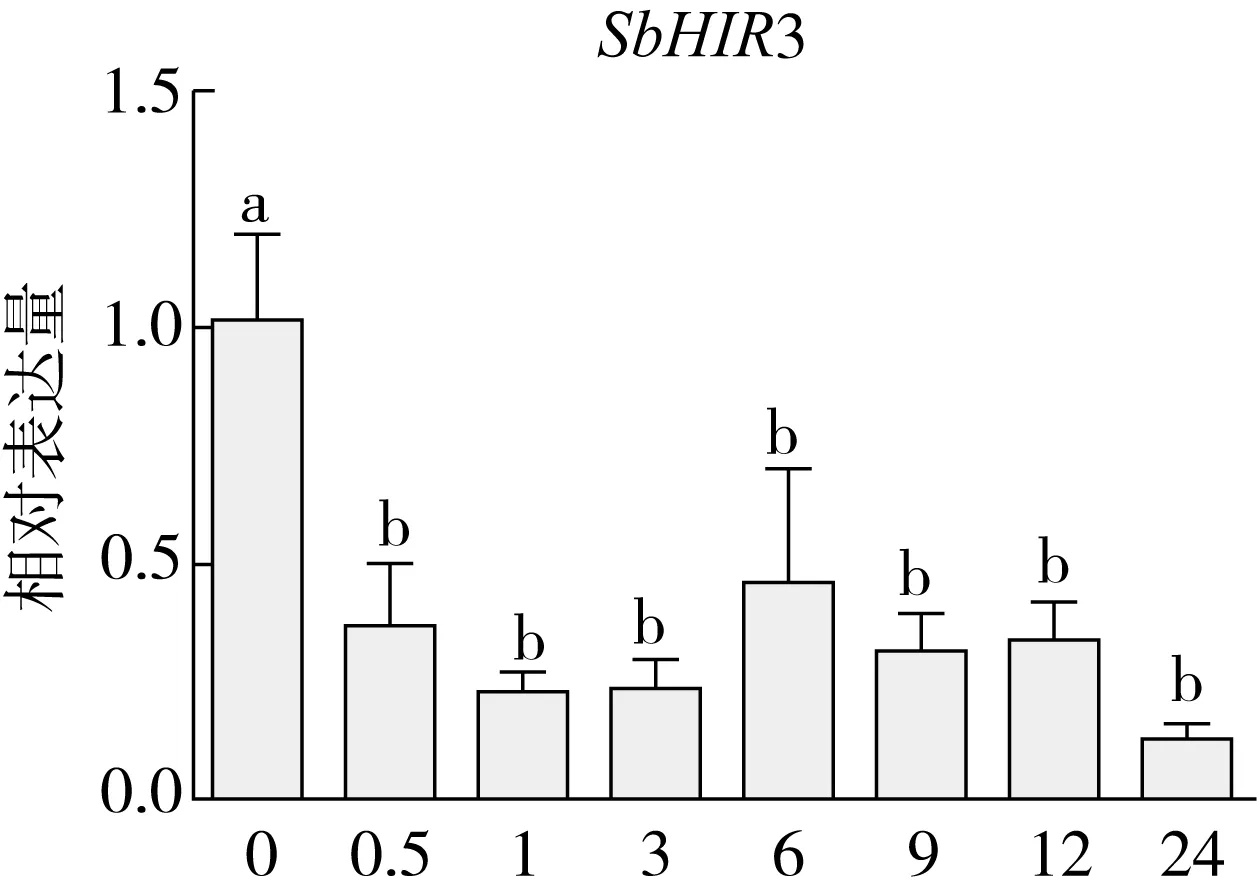
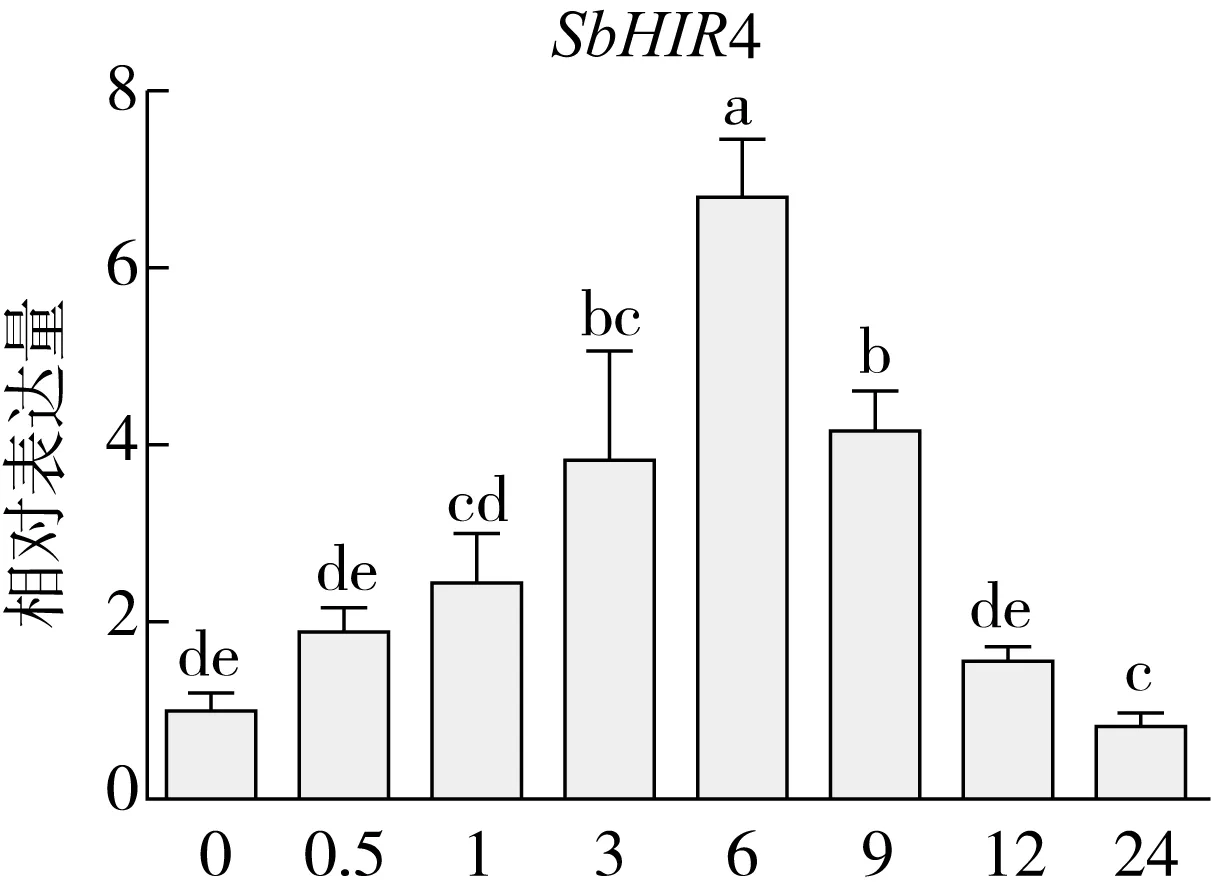
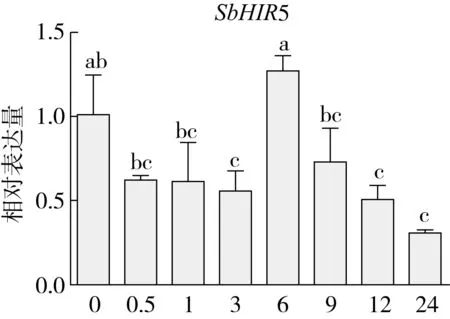
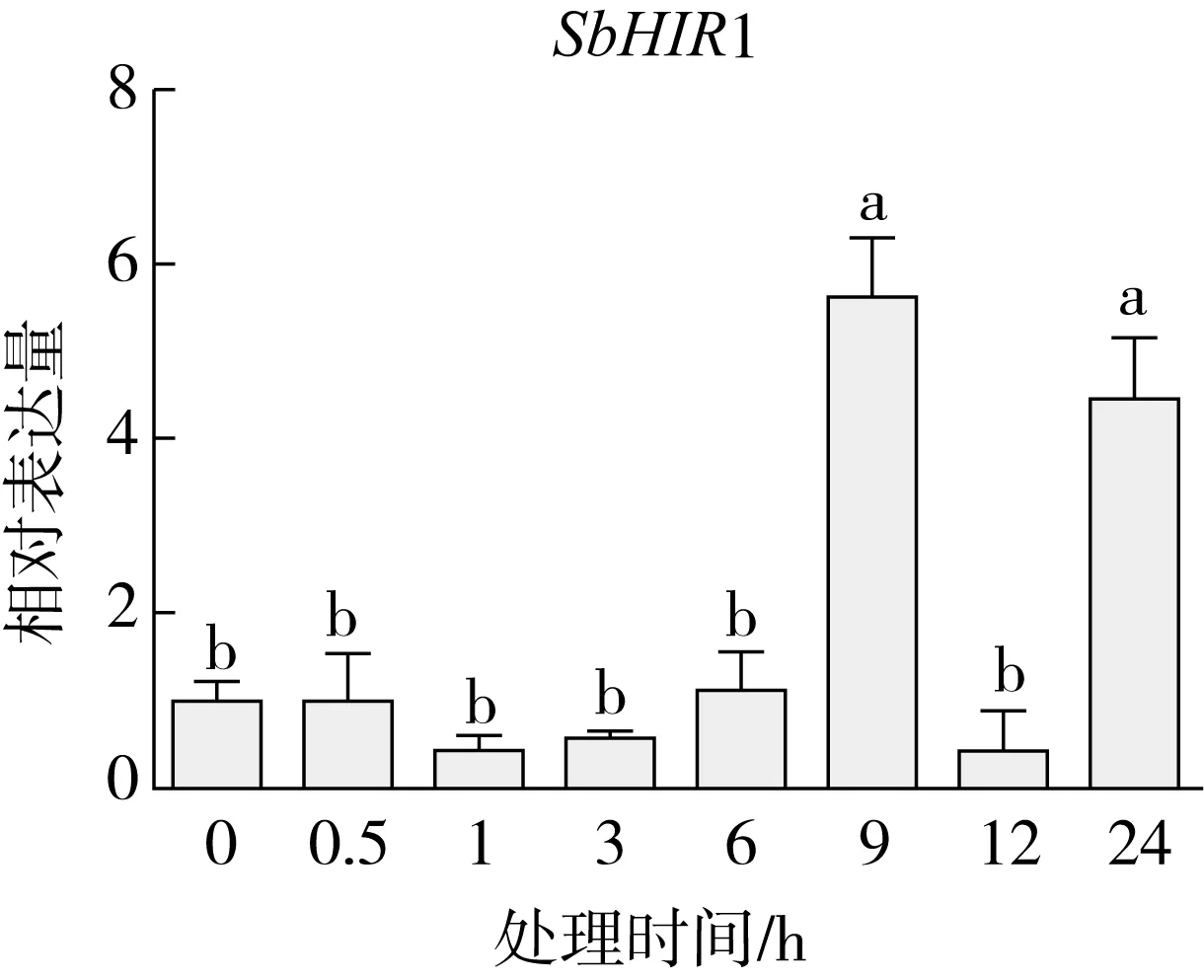
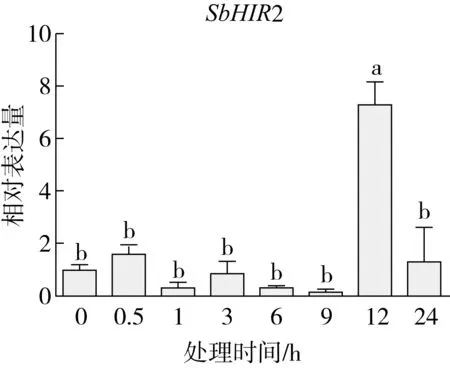
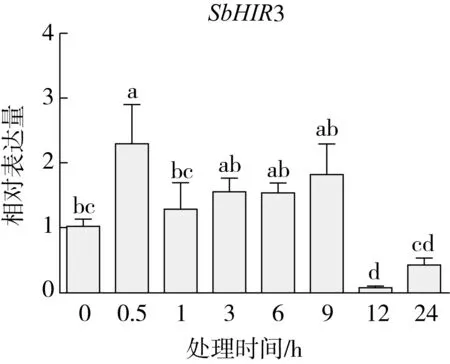
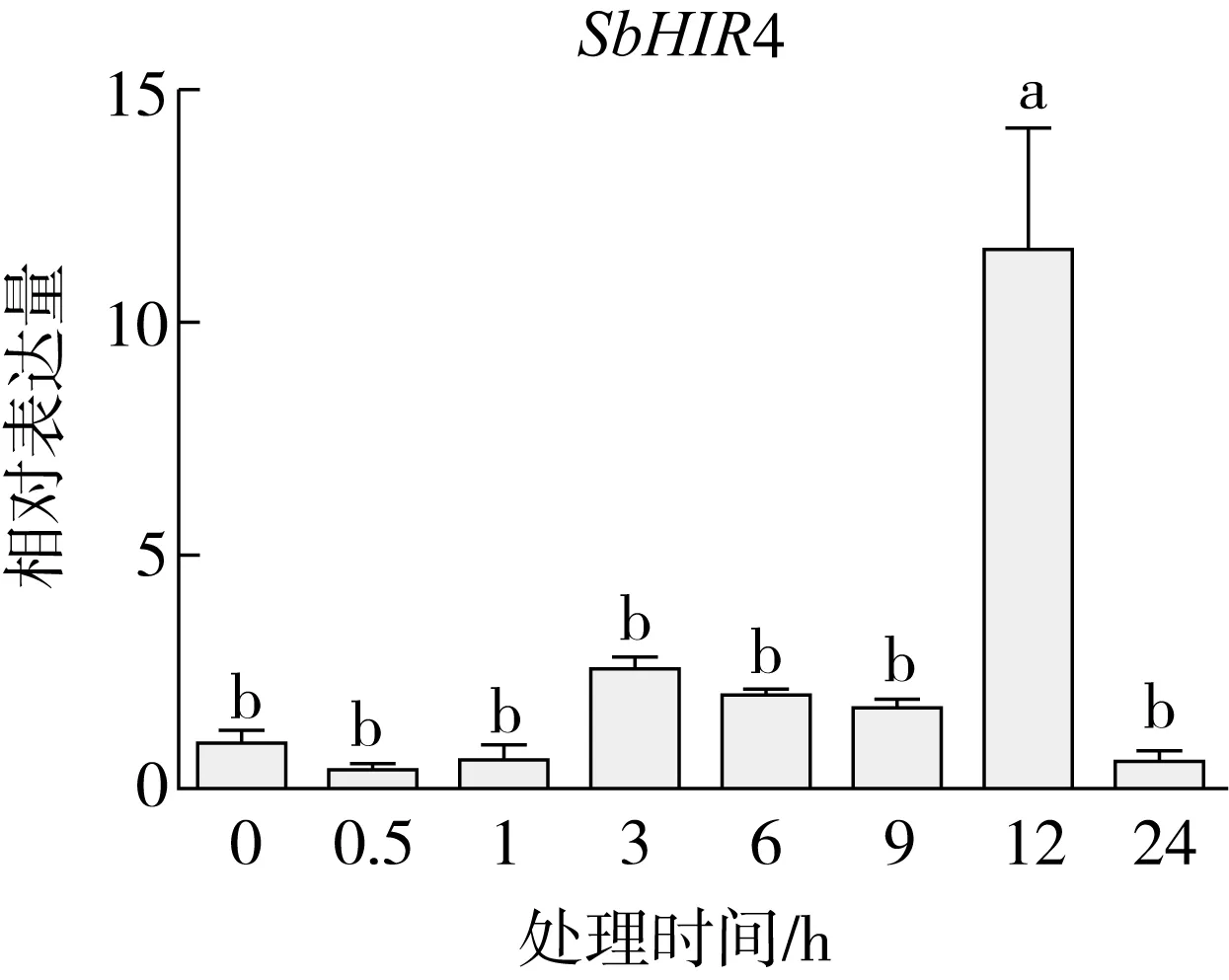
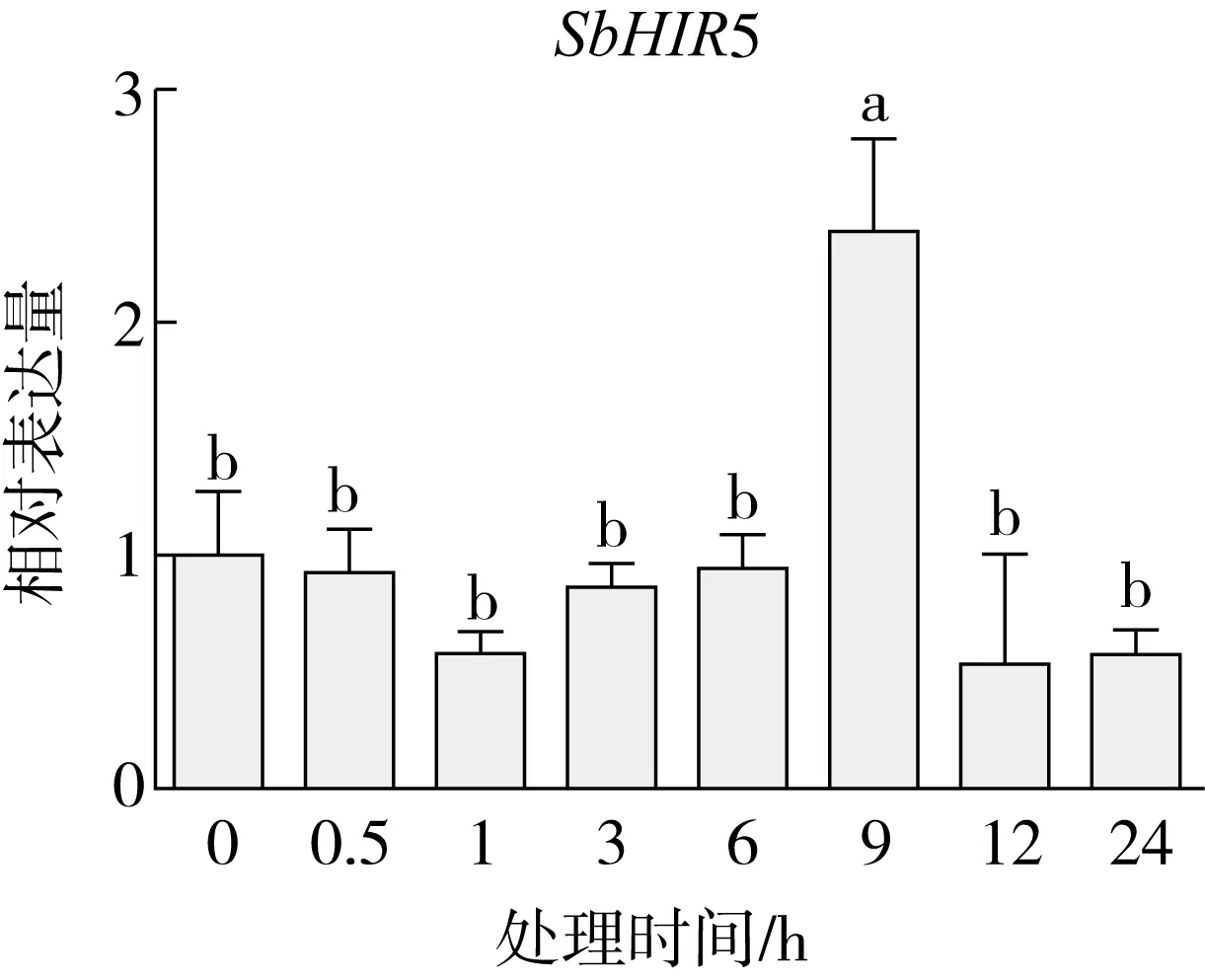
研究高粱HIR家族基因结构分布情况,及拟南芥、水稻、玉米、小麦、大麦、辣椒和高粱HIR家族成员的功能完整性,对高粱HIR家族基因结构、保守基序(Conserved motif,Motif)进行分析。使用GSDS2(http://gsds.cbi.pku.edu.cn/)在线工具对高粱HIR家族基因结构进行分析;利用MEME(http://meme-suite.org/)在线网站(参数:筛选数量为10,E-value 1.2.5 高粱HIR蛋白二级结构分析 使用在线网站NPSA-PRABI(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_seccons.html),对高粱HIR蛋白二级结构组成进行预测。 1.2.6 高粱种植和样品处理 选择健康高粱种子在清水中浸种24 h,随后用滤纸保湿48 h进行催芽,将发芽的种子播种在灭菌营养土中,在25 ℃、光照12 h/黑暗12 h的培养箱中培养到3叶期,取高粱幼苗的根、茎、叶,于-80 ℃保存备用;另一方面将高粱幼苗的根部浸泡在分别含有ABA、GA3和flg22的营养液中,其处理浓度分别为200 μmol/L、100 μmol/L、100 nmol/L,然后分别在处理后0、0.5、1、3、6、9、12、24 h取样,设置3个生物学重复,每个生物学重复处理5株,所取样品用液氮速冻,置于-80 ℃保存备用。 1.2.7 高粱HIR基因表达分析 采用qRT-PCR技术进行分析,利用qRT-PCR检测高粱SbHIR1~SbHIR5基因的表达量,其反应体系为:cDNA 4.5 μL,SYBR Premix 7.5 μL,SbHIRs基因的上下游引物各 0.3 μL,ddH2O补足15 μL;反应程序为:95 ℃ 10 min,95 ℃ 10 s,60 ℃ 30 s,40个循环,95 ℃ 10 s,60 ℃ 60 s,1个循环;设置 3 个技术性重复,其数据使用2-ΔΔCt方法进行分析,利用DPS软件进行显著性分析,其qRT-PCR引物如表1。 为研究HIR家族基因在高粱中所存在的成员数,利用Hmmer软件进行筛选。由表2和图1可知,在高粱中鉴定出5个HIR基因,根据所在高粱染色体中的位置命名为SbHIR1~SbHIR5;分别处于第2、3、7、9、10号染色体上;SbHIR1、SbHIR2、SbHIR4和SbHIR5基因所编码的氨基酸数量、相对分子质量大小和理论等电点较为接近,而SbHIR3与其他家族成员的氨基酸序列长度和蛋白分子量大小差异较大分别为413 aa、45.02 ku;高粱HIR家族蛋白均为亲水性蛋白;亚细胞定位预测显示均定位于线粒体中。 图1 高粱HIR基因在染色体上的分布 表2 SbHIRs基因编码蛋白的基本特征 构建拟南芥、水稻、玉米、小麦、大麦、辣椒和高粱的HIR家族成员系统进化树。由图2可知,该进化树一级分支分为两支,在二级树分支内双子叶植物(拟南芥、辣椒)HIR家族成员明显区分于其他几种单子叶植物(水稻、玉米、小麦、大麦和高粱),早熟禾亚科(小麦、大麦)、稻亚科(水稻)和黍亚科(高粱、玉米)的HIR成员明显分开,各自在不同的分支中,而早熟禾亚科与黍亚科HIR成员形成姐妹基因对;相对特殊的是,在同一支内,SbHIR2明显与其他单子叶HIR成员形成较大的差异(支持率为100%),单独形成一个特有支;高粱HIR成员与玉米HIR家族成员的亲缘关系较近。 图2 拟南芥、水稻、玉米、小麦、大麦、辣椒和高粱HIR成员的系统进化分析 由表3可知,不同物种间及物种内HIR基因的密码子选择性较为保守,有效密码子数在33.243~58.430之间,而SbHIR2的有效密码子数仅为33.243,说明密码子偏好较强;单子叶植物(水稻、玉米、小麦、大麦和高粱)大部分HIR基因GC和GC3s的含量大于50.00%,即水稻、玉米、小麦、大麦和高粱的HIR基因编码区对碱基的选择更偏向于G+C,且密码子偏好于以G/C结尾,且该情况在高粱SbHIR2上较为明显。而双子叶植物(拟南芥、辣椒)HIR基因GC和GC3s的含量均小于50.00%,对碱基G+C和密码子G/C结尾的偏好性较弱。不同物种间及物种内HIR基因的有效密码字数相对保守,单子叶和双子叶植物HIR基因的碱基偏好及密码子的结尾的偏好有所不同。 表3 拟南芥、水稻、玉米、小麦、大麦、辣椒和高粱HIR基因的密码子偏好分析 由图3a可知,高粱HIR基因内含子数为1~6个,基因结构相对简单,仅有SbHIR3基因缺乏上游UTR。由图3b可知,拟南芥、水稻、玉米、小麦、大麦、辣椒和高粱的HIR基因Motif均相对保守,具备9个Motif(OsHIR2和SbHIR2除外),而OsHIR2和SbHIR2均出现Motif 8的缺失,同时,SbHIR2也缺失Motif 5。 由表4可知,SbHIR1~SbHIR5二级结构均以α-螺旋为主,其次是无规卷曲、延伸链,β-转角占比最低。 表4 高粱HIR蛋白二级结构组份 研究高粱HIR基因家族的组织特异性表达和在植物激素(ABA和GA3)和flg22胁迫处理下的表达量。通过qRT-PCR分析可知,SbHIR1基因在茎中的表达量约为根和叶的4倍;SbHIR2基因在根、茎、叶中均有表达,但在根中表达量最高;SbHIR3、SbHIR4和SbHIR5基因均在叶中表达量最高(图4)。 图4 高粱HIR基因家族成员组织表达分析 在植物激素ABA处理后,高粱HIR基因家族各个成员的表达模式不同,SbHIR1和SbHIR5的表达量在12 h时达到最高;SbHIR2和SbHIR3基因在0.5 h时表达量最高,随后表达量降低;SbHIR4基因在6 h时表达量最高(图5)。 在植物激素GA3处理下,高粱SbHIR1基因的表达量呈先降后升的趋势,在3 h时表达量最低,然后又逐渐上升,在12 h时表达量达到最高;SbHIR2和SbHIR3基因在GA3处理下,其表达均受到一定程度的抑制;SbHIR4和SbHIR5基因在GA3处理下均在6 h时表达量达到最高(图6)。 注: Motif 1~9分别用数字1~9表示。图3 高粱HIR的基因结构和拟南芥、水稻、玉米、小麦、大麦、辣椒和高粱HIR蛋白保守基序分析 图5高粱HIR基因家族成员ABA处理的表达分析 图6高粱HIR基因家族成员GA3处理的表达分析 在flg22胁迫处理下,SbHIR1的表达量呈现出下降-上升-下降-上升的趋势,在9 h达到最高;SbHIR2和SbHIR4,均在12 h时表达量达到最高;而SbHIR3在各时间段内表达量均有一定程度的变化,但12 h时表达量最低;SbHIR5的表达量呈现出下降-上升-下降的趋势,在9 h达到最高(图7)。 图7高粱HIR基因家族成员flg22处理的表达分析 利用生物信息学方法和qRT-PCR技术在全基因组水平上对高粱HIR基因家族进行鉴定,共鉴定得到5个成员;5个成员之间的理化性质具有差异,且均为亲水性蛋白;系统进化分析发现高粱HIR基因与玉米的亲缘关系较近;密码子偏好分析表明,高粱的HIR基因编码区对碱基的选择更偏向于G+C,且密码子偏好于以G/C结尾;高粱HIR基因家族成员的二级结构均以α-螺旋为主;高粱HIR基因家族成员在根、茎和叶中均能够表达,且在植物激素(ABA和GA3)的处理下,其在高粱叶片中的表达量均出现了不同程度的变化,当flg22胁迫处理时其高粱HIR基因家族成员的表达量变化趋势较为明显。2 结果与分析
2.1 高粱HIR基因家族的鉴定和基本理化性质分析
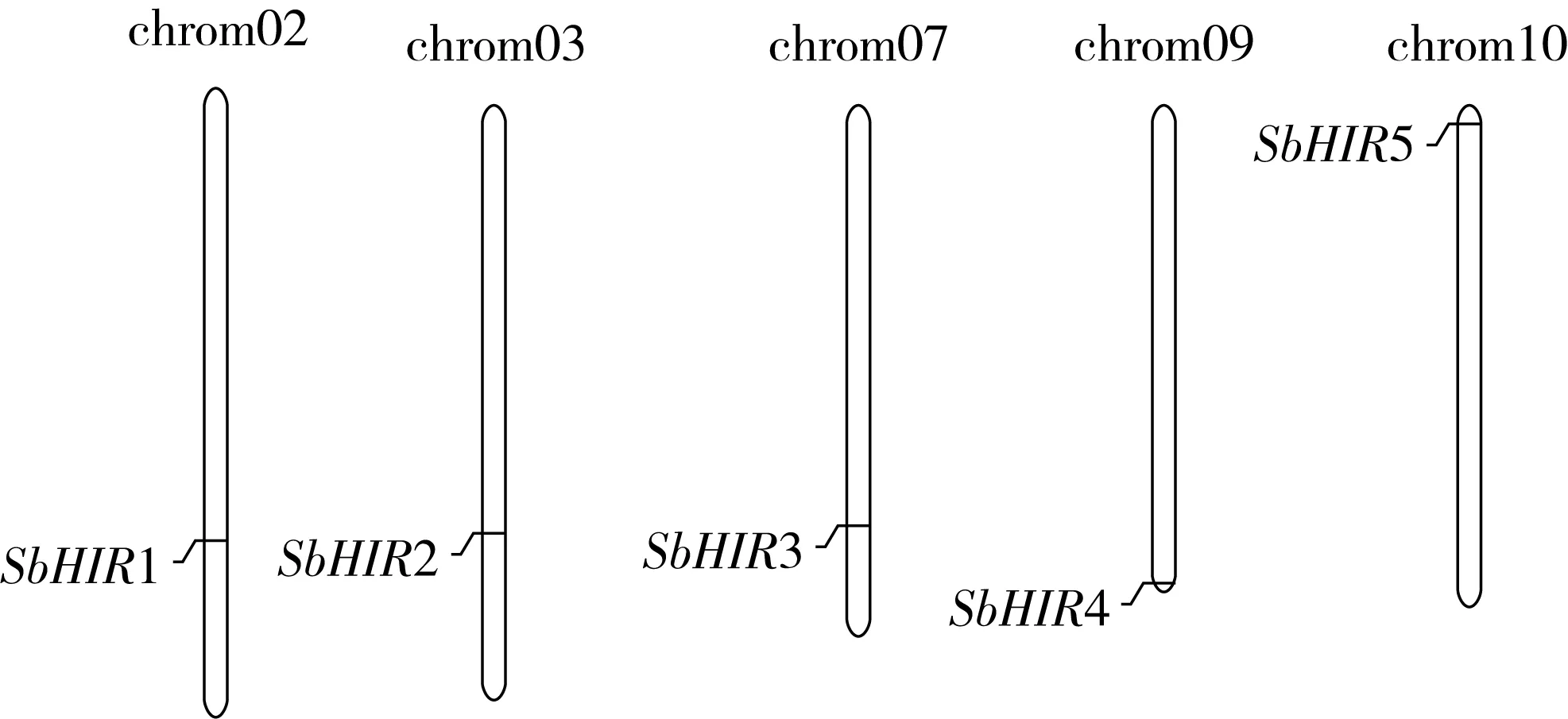
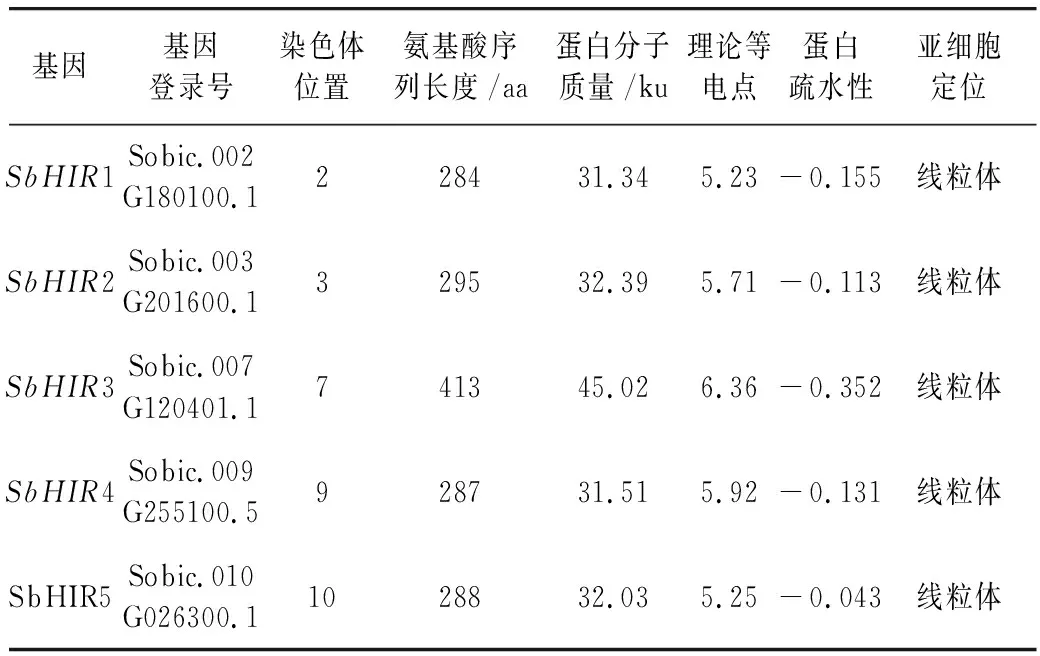
2.2 高粱HIR家族系统发育分析
2.3 HIR家族基因密码子偏好分析
2.4 高粱HIR家族基因结构和HIR家族蛋白的Motif分析
2.5 高粱HIR蛋白二级结构分析
2.6 高粱HIR基因表达分析
3 结论
猜你喜欢
作文小学高年级(2022年5期)2022-06-16
环境卫生工程(2021年4期)2021-10-13
疯狂英语·新读写(2021年2期)2021-02-25
生物学通报(2020年11期)2020-10-22
国际放射医学核医学杂志(2020年4期)2020-07-27
中成药(2018年7期)2018-08-04
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年3期)2017-04-10
华人时刊(2016年19期)2016-04-05
植物营养与肥料学报(2014年1期)2014-03-11
