
茶叶香气物质响应胁迫机制与功能研究进展
2021-08-31 03:30:28王登良
食品科学 2021年15期
闫 振,王登良,2,*
(1.广东德高信茶业控股有限公司,广东 英德 513000;2.华南农业大学园艺学院,广东 广州 510642)
茶叶作为世界三大饮料之一,以其独特的风味品质、药理功能和文化价值成为仅次于水的普遍饮品。茶叶独特的风味品质取决于其丰富的内含物质,茶叶香气是茶叶感官审评的重要因子,是茶叶品质的重要贡献部分。茶叶香气的主体芳香物质(也称挥发性香气组分)按照合成途径的不同主要分为挥发性萜类 化合物(volatile terpenes,VTs)、挥发性脂肪酸分解产物(volatile fatty acid decompositions,VFADs)和挥发性苯丙酸/苯类化合物(volatile phenylpropanoid/benzenoid,VPBs)[1]。成品茶叶香气物质种类繁多、组成复杂,其组分含量与茶树品种、地域环境、栽培条件和加工工艺等因素密切相关,其中茶叶采前的 生长环境条件和采后的加工工艺,尤其是后者,通常对茶叶香气品质具有决定性作用。
根据胁迫因素的性质可将胁迫分为生物胁迫(食叶害虫、病原菌等)和非生物胁迫(光照、温度、湿度、机械损伤等)两大类[2-3]。植物胁迫响应是当植物受到环境胁迫因素刺激时,植物通过识别防御信号启动体内的防御系统并通过一系列的生理反应以抗衡和降低胁迫伤害的一种自我保护反应。食叶害虫取食和病原菌侵染不仅会严重影响茶叶的产量和外观品质,同时也会诱导茶叶形成并释放大量的香气物质[4]。采后加工过程是茶叶香气品质形成的关键阶段,加工工艺产生的机械损伤、失水和高温等非生物胁迫对香气物质的合成与释放有重要影响[1]。本文总结了生物胁迫和非生物胁迫对茶叶香气物质代谢的调控机制以及茶叶香气响应胁迫在实际生产中的应用价值,以期为改善茶叶香气品质和增强茶树病虫害防御提供理论依据。
1 茶叶香气物质
1.1 茶叶中的香气化合物
相比于茶多酚和氨基酸等物质,茶叶香气物质在茶叶中的含量很低,仅占茶叶干质量的0.03%左右,但茶叶香气物质种类繁多,利用气相色谱-质谱联用(gas chromatography-mass spectrometry,GC-MS)和核磁共振(nuclear magnetic resonance,NMR)技术已从茶叶中分离鉴定到700余种香气物质[5]。另一方面,由于大多数茶叶香气物质的阈值较高而无法直接被人体感知,茶叶中重要的香气成分仅有几十种低阈值香气物质[6]。如Schuh等[7]在全发酵红茶中鉴定出600余种挥发性香气组分,仅发现41 种对红茶香气品质有突出贡献;Kumazawa等[8]从不发酵绿茶中鉴定到200余种挥发性香气组分,仅发现 30 种对绿茶香气品质有贡献。茶叶中的香气物质根据代谢来源的不同可分为:VTs类,包括芳樟醇、香叶醇、橙花叔醇、法尼烯和罗勒烯等;VFADs类,包括(Z)-3-己烯-1-醇、(Z)-3-己烯基乙酸、(E)-2-己烯醛和(E)-2-己烯酸等;VPBs类,包括苄基腈、苯甲醛和吲哚等[9-10]。
1.2 香气化合物在茶叶中的合成与释放
多数香气物质是通过从头途径(de novo)合成的,而酶在细胞内的定位和与底物接触的有效性是决定香气产物形成的关键因素,如细胞质萜类合成酶(terpenoid synthase,TPS)由于底物法尼烯焦磷酸的有效性(存在于细胞质)主要参与倍半萜的生物合成,而定位于质体中的TPS由于底物香叶基焦磷酸的有效性(存在于质体中)则主要参与单萜的生物合成[11]。如茶树芳樟醇合成酶基因CsLIS/NES存在CsLIS/NES-1和CsLIS/NES-2两个亚型,CsLIS/NES-1定位于叶绿体,主要催化香叶基焦磷酸合成芳樟醇;CsLIS/NES-2定位于细胞质,主要催化法尼烯焦磷酸合成橙花叔醇[12]。茶叶香气物质在茶树体内的存在方式主要有游离态(free volatile compounds,FVCs)和糖苷结合态(glycosidically bound volatile compounds,GBVs)两种。GBVs的水溶性和化学稳定性较FVCs更佳,且不会对植物产生毒害作用,因此大多数香气物质主要是以GBVs的方式存在于茶叶中,这也是茶鲜叶在加工前只散发出轻微的青气而没有丰富香气的主要原因。茶叶香气物质从游离态到糖苷态的转变是在尿苷二磷酸糖基转移酶(uridine diphosphate-glycosyltransferases,UGTs)的作用下进行[13-14]。从茶树基因组中已发现300多个UGTs[15-16], 而参与香气糖苷合成的只有少数UGTs已被验证功能,如UGT85K11和UGT94P1分别参与香叶醇β-葡萄糖苷和香叶醇β-樱草糖苷的形成[17-19],UGT74AF3参与呋喃的糖基化反应[20]等。茶叶香气物质在糖苷水解酶的催化作用下由糖苷态水解成游离态[21-22]。茶叶中起关键作用的糖苷水解酶主要有β-樱草糖苷酶、β-葡萄糖苷酶和β-半乳糖苷酶3 种[23-24]。目前,从茶叶中已分离鉴定出1 种β-樱草糖苷酶和3 种β-葡萄糖苷酶[25-27](表1)。GBVs主要存在于液泡,而糖苷水解酶定位于细胞壁[28],因此在细胞完整状态下,由于腔室分隔作用,两者无法接触而不会发生GBVs的水解。加工过程的失水和机械损伤导致细胞破碎或细胞膜透性增加,从而使GBVs与糖苷水解酶相互接触并释放FVCs[27],这也是香气物质种类和含量在茶鲜叶中较少而在成品茶中丰富的主要原因。

表1 茶叶香气物质糖基转移酶和糖苷水解酶Table 1 Glycosyltransferases and glycosidases hydrolyzing tea aroma compounds
2 生物胁迫对茶叶香气物质生物合成及调控的影响
小绿叶蝉、茶尺蠖、茶蚜虫、黑刺粉虱和茶卷叶蛾等是危害茶树生长的常见茶园害虫[30];茶炭疽病、茶云纹叶枯病、茶轮斑病、茎点霉真菌、茶褐枯病、茶饼病、茶赤叶斑病等是普发性的茶叶病害[31-33]。病原菌侵染可诱导茶叶芳樟醇、香叶醇、苯乙醇、(Z)-3-己烯醇和水杨酸甲酯等香气物质的释放[34];红蜘蛛取食可诱导茶叶释放大量的罗勒烯和α-法尼烯[3];茶卷叶蛾可促使茶叶释放(Z)-3-己烯-1-醇、芳樟醇、α-法尼烯、苄腈、吲哚、橙花叔醇和罗勒烯等;茶蚜虫可诱导茶叶释放(E)-2-己烯醛、 罗勒烯、(Z)-3-乙酸己烯酯、(Z)-3-己烯-1-醇、丁酸-3-己烯酯、芳樟醇、香叶醇、吲哚、正辛醇等挥发性物质[35]。
2.1 食叶害虫取食对茶叶香气物质的影响
食叶害虫取食茶树叶片对茶叶香气的影响方式主要有取食方式造成的机械损伤和取食过程分泌的诱导因子两种。一方面,食叶害虫取食方式(如咀嚼式和刺吸式等)直接对茶叶造成的机械损伤可影响茶叶香气等挥发性物质的合成与释放。害虫对植物的机械损伤往往是通过激活叶片内部与胁迫信号传导相关的丝裂原活化蛋白激酶(mitogen-activated protein kinases,MAPKs)和钙依赖蛋白激酶的活性从而提高茉莉酸、水杨酸、茉莉酸-异亮氨酸衍生物和乙烯等植物激素的表达水平,这些植物激素特别是茉莉酸可以激活多种挥发性物质合成酶基因的表达并最终导致挥发性物质的合成与释放[36-37]。另一方面,食叶害虫特别是咀嚼式害虫在取食茶叶的同时往往分泌一些诱导因子如β-葡萄糖苷酶、脂肪酸-氨基酸共轭衍生物和一种热不稳定化合物等,这些诱导因子可通过与茶叶内部化合物发生酶促反应促进茶叶香气等挥发性物质的合成与释放[38-40]。
茶卷叶蛾、红蜘蛛、茶尺蠖、小绿叶蝉取食茶叶时均会诱导茶叶释放大量的α-法尼烯[4]。茶叶被小绿叶蝉取食后,作为信号物质的茶叶内源茉莉酸大量生成,并上调α-法尼烯合成酶基因CsAFS的表达,从而促进α-法尼烯的合成与释放[41]。脂肪酸过氧化氢裂解酶(hydroperoxide lyase,HPL)是植物挥发性醛和醇的重要合成酶,由HPL催化形成的挥发性醛和相应的还原醇是水果、蔬菜等挥发性气味的主要贡献成分。茶尺蠖咀嚼茶叶会显著诱导茶树脂肪酸HPL基因CsHPL表达上调,并促进不饱和脂肪酸氢过氧化物分解为醛和含氧酸,而这些醛类化合物特别是(Z)-3-己烯醛是茶叶香气的重要组成成分[42]。除此之外,被茶尺蠖取食茶叶中的脂氧合酶(lipoxygenase,LOΧ)基因如CsLOX2、CsLOX3、CsLOX4、CsLOX6、CsLOX7的表达水平显著上调,CsLOX2、CsLOX3可分别催化9/13-过氧羟基-十八碳三烯酸(9/13-hydroperoxy-octadecatrienoic acid,9/13-HPOT)和13-HPOT的形成,而9/13-HPOT和13-HPOT均为HPL代谢途径形成茶叶香气物质醛和醇的相应底物[43]。
小绿叶蝉取食茶叶会促进茶叶香气物质萜类化合物如(S)-芳樟醇的释放,而取食造成的持续机械性损伤是诱导(S)-芳樟醇大量形成并释放的关键因素。一方面,小绿叶蝉取食可诱导茶叶(S)-芳樟醇合成酶基因CsLIS1和CsLIS2的表达水平上调,继而催化香叶基焦磷酸释放(S)-芳樟醇;另一方面,小绿叶蝉取食能够诱导上调β-樱草糖苷转移酶和β-樱草糖苷水解酶的表达水平,前者促进(S)-芳樟醇β-樱草糖苷的合成,而后者则促进(S)-芳樟醇β-樱草糖苷的水解并释放(S)-芳樟醇[44]。除了(S)-芳樟醇,小绿叶蝉取食茶叶还会促进香叶醇和其他一些单萜物质的释放,但香叶醇的释放并不是来源于机械损伤 引发的茶叶内部的响应调控途径,而是小绿叶蝉分泌物中的香叶醇合成酶催化茶叶香叶醇的合成与释放。此外,在小绿叶蝉取食的茶叶中还会形成一种特殊的香气成分即双烯雌酚I,而其他食叶害虫,如茶蚜虫、茶尺蠖、茶卷叶蛾、红蜘蛛取食的茶叶中均未发现双烯雌酚I的释放,这暗示在小绿叶蝉分泌物中应存在一种可催化双烯雌酚I合成的独特诱导因子[1]。
2.2 病原菌侵染对茶叶香气物质的影响
茶叶感染茶炭疽杆菌后,一种12-氧植物二烯酸还原酶的表达水平显著上升,并将12-氧植物二烯酸最终转化为茉莉酸,导致茉莉酸含量在感病茶叶中增加[45]。此外,茶炭疽病侵染可以激活茶叶中苯丙氨酸解氨酶的活性以及LOS途径并促进VFADs、VPBs等挥发性物质的释放[46]。茶云纹叶枯病也称作茶树叶枯病,王瑾等[47]研究指出当茶叶受到茶云纹叶枯病致病菌侵染时,GBVs水解释放出游离态挥发性糖苷元抑制病菌的发展。Zhang Zhengzhu等[34]发现茶炭疽病菌、茶云纹叶枯病菌、茶轮斑病菌和茶赤叶斑病菌4 种致病菌侵染可在染病初期不同程度地诱导上调茶树内源β-樱草糖苷酶、β-木糖苷酶、β-葡萄糖苷酶I、β-葡萄糖苷酶II和β-葡萄糖基转移酶基因的表达,并释放香叶醇、苯甲醇、苯乙醇、(Z)-3-己烯醇、水杨酸甲酯、芳樟醇和芳樟醇氧化物等主要的茶叶FVCs。茶饼病是一种低温高湿型病害[48],茶叶感染茶饼病时,包括萜烯类、芳香族化合物、含硫化合物和绿叶挥发物等在内的挥发性物质组分和含量显著增加,其中顺-3-己烯基乙酸酯和2-乙基己醇在感病茶梢中的含量大幅增加[49],而这些化合物对病原菌的侵染具有一定的抑制作用[50]。采前茶叶香气与胁迫响应的关系如图1所示。
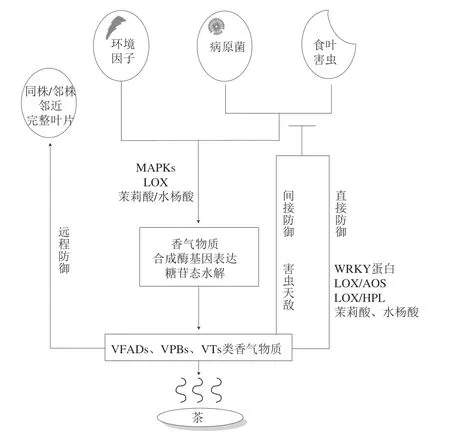
图1 采前茶叶香气物质与胁迫响应关系Fig.1 Relationship between aroma components and stress response in preharvest tea leaves
3 非生物胁迫对茶叶香气物质生物合成及调控的影响
3.1 环境胁迫因子对茶叶香气物质的影响
温度、海拔、光照等条件是影响茶鲜叶中芳香物质形成的重要环境因子,多数芳香物质在茶叶中的含量与环境温度变化呈现一定的规律性,如清香型的戊烯醇、己烯醇等香气物质在春茶中的含量高于夏茶;花果香型的苯乙醇、苯乙醛等香气物质在秋茶中的含量较高;秋冬茶香高馥郁,低温胁迫是其形成的重要原因之一;高山茶香气高的特点与高山茶园夜晚温度较低有关[51-52]。曹藩荣等[52]对岭头单枞茶树品种施加不同程度的低温处理,发现低温胁迫可显著促进茶鲜叶中芳香物质的合成,且10~15 ℃低温对岭头单枞茶叶香气成分的种类和含量影响最大。Zhou Hanchen等[53]发现冷害、盐胁迫和干旱等胁迫因素能够显著抑制多数茶树香气物质TPS基因CsTPS的表达,少数CsTPS基因响应胁迫而增强表达。Zhu Junyan等[43]研究发现短期低温胁迫可诱导茶叶LOΧ基因CsLOX1、CsLOX6和CsLOX7的表达,并促进香气物质醛和醇的合成与释放,而长期低温胁迫则不利于茶叶LOΧ基因的表达。
茶园遮荫管理是调控茶树生长期间光照条件的常用方法。茶树遮荫有利于改善茶叶天然产物如氨基酸、类黄酮等的积累并提高茶叶品质[54-55],同时也有助于促进茶叶香气物质的形成。Yang Ziyin等[56]发现遮荫处理可显著促进茶叶VFADs和VPBs等挥发性物质合成与释放,遮荫处理诱导茶叶香气物质的形成与GBVs的水解没有直接关系,而主要与其上游代谢途径有关。如L-苯丙氨酸是大多数VPBs合成的关键前体物质,L-苯丙氨酸的脱氨反应是莽草酸途径中VPBs合成的第一步,遮荫处理后的茶叶L-苯丙氨酸与其他芳香类氨基酸如L-酪氨酸和L-色氨酸的含量均显著增加,表明遮荫处理对茶叶香气的形成有一定的促进作用。除此之外,单一波长光对茶叶香气的形成也具有一定的促进作用。Fu Χiumin等[57]发现与自然光照或黑暗条件相比,蓝光和红光可以通过上调茶树LOΧ基因(如CsLOX2、CsLOX3、CsLOX4、CsLOX9和CsLOX13)、苯丙氨酸解氨酶基因和TPS基因 (如CsTPS1、CsTPS2和CsTPS3)的表达而诱导采前茶叶挥发性香气成分的合成与积累。与遮荫处理相比,自然光照和红蓝光则不能显著诱导L-苯丙氨酸含量增加,L-苯丙氨酸在苯丙氨酸解氨酶的催化下可以转化为肉桂酸,而苯丙氨酸解氨酶的表达水平显著受红蓝光的诱导,这可能是红蓝光辐射下苯甲醇和苯甲醛含量增加的原因[57]。
茉莉酸是植物响应机械伤害、病虫害、低温以及盐胁迫等逆境的重要信号物质[58]。芳樟醇及其氧化物和香叶醇分别对红茶甜香味和花香味的形成具有贡献,对茶鲜叶施加外源茉莉酸甲酯(methyl jasmonate,MeJA)处理 可提高其成品红茶的香气品质。Shi Jiang等[59]研究发现β-葡萄糖苷酶的基因表达水平和活性在MeJA诱导后的茶鲜叶中均明显降低,而β-樱草糖苷酶基因表达水平被上调,萜烯醇类如香叶醇、芳樟醇及其氧化物和萜烯类如石竹烯氧化产物、α-法尼烯和δ-杜松烯等的含量明显提高。此外,在MeJA处理诱导茶鲜叶中生成了3 个新组分——古巴烯、荜澄茄油烯醇和吲哚,古巴烯具有花香,荜澄茄油烯醇具有清新的香气,而吲哚具有辛辣的香气。Zhou Hanchen等[53]发现CsTPS基因中的27 个的表达水平在茶叶经MeJA处理后显著上调,12 个则被明显抑制,表明MeJA可显著改变茶鲜叶中CsTPS基因的表达水平,从而影响香气物质的代谢。
3.2 加工工艺对茶叶香气物质的影响
在白茶、红茶或乌龙茶的加工过程中,萎凋不仅有助于降低茶叶的苦味,还有助于(Z)-3-己烯醇等青草气成分的挥发,并促进芳香物质的释放,从而改善茶叶香气品质[60]。红茶发酵过程伴随着氨基酸的脱羧反应并形成醛类化合物;类胡萝卜素发生初级氧化和次级氧化,并产生β-紫罗兰酮、茶螺烯酮、二氢海葵内酯等;不饱和脂肪酸发生分解反应并形成大量的醇和醛类;这些代谢产物共同贡献于红茶的香气品质。Wang Lifei等[61]指出(E)-2-己烯醛、苯甲醛、6-甲基-5-庚烯-2-酮、水杨酸甲酯和吲哚5 种挥发性物质是区分不发酵茶和发酵茶的关键香气成分,(E)-2-己烯醛和水杨酸甲酯也是半发酵茶和全发酵茶的特征性香气挥发性成分。绿茶的香气成分相对于红茶和乌龙茶要少得多,不同杀青和干燥工艺对绿茶香气特征形成的影响不同,如蒸青绿茶含有较多鲜爽型的芳樟醇及其氧化物,青草香较明显,而炒青绿茶则含有较多苯甲醇、香叶醇、吡嗪、吡咯等物质,香型以焦糖香和栗香为主[5]。乌龙茶的香气成分主要来源于萜烯类和芳香醇配体糖苷的水解,以及脂肪酸的氧化裂解和类胡萝卜素的氧化降解产物[62]。与绿茶、红茶等茶类相比,乌龙茶的加工工艺相对复杂,使其具有特殊的蜜香和花果香。由于加工工艺的特殊性,茶叶细胞完整度和细胞活性在乌龙茶加工过程中保持的最长久,并使酶促反应在采后茶鲜叶中得以充分地进行。同时,复杂的加工过程使茶叶受到多种胁迫,如采青产生的机械性伤害、晒青引发的失水和紫外线辐射、晾青导致的失水反应以及摇青造成的持续性机械损伤等[63]。乌龙茶生产过程的综合胁迫响应成为研究采后加工过程茶叶香气形成的良好模型[1,27]。Baldermann等[64]对38 种不同品种、生产工艺和产地的绿茶、乌龙茶和红茶的挥发性成分进行比较,发现吲哚、茉莉酸内酯和(E)-橙花叔醇在乌龙茶摇青阶段大量形成并释放,这些香气成分协同芳樟醇、香兰素、呋喃酮、β-紫罗兰酮等共同贡献于乌龙茶的特征香气[29,65]。乌龙茶加工过程导致(E)-橙花叔醇的大量形成与释放,(E)-橙花叔醇合成酶基因CsNES的表达水平和(E)-橙花叔醇的含量在摇青阶段,特别是在第3~5次摇青过程中显著升高,而摇青工艺对茶叶造成的持续性机械损伤是诱导CsNES基因表达并释放(E)-橙花叔醇的关键因素[27]。莽草酸途径色氨酸合成酶亚基蛋白CsTSA和CsTSB2的复合体是催化吲哚-3-甘油磷酸酯合成吲哚的关键单元,CsTSB2蛋白水平在乌龙茶摇青过程中大量增加,而摇青工艺产生的持续性机械损伤是诱导CsTSB2基因表达并使吲哚大量积累的主要原因[66]。虽然红茶加工的揉捻工艺也会产生机械损伤,但红茶中的吲哚含量比乌龙茶要少得多,红茶揉捻过程对茶叶细胞的破坏并没有促使吲哚的转化,而是导致了吲哚合成的终止。
胁迫响应与植物挥发性物质关系的研究表明,多重胁迫对植物挥发性物质的形成与释放具有叠加的效果[10]。 Ai Zeyi等[67]研究单一光波长对红茶加工萎凋过程中品质形成的影响发现,萎凋过程辅加黄、橙、红光能够不同程度地影响茶叶儿茶素、氨基酸、茶黄素和香气成分的含量,具有改善茶叶香气和滋味的良好作用,但绿光辐射则会诱导茶叶产生大量的青草气,从而破坏茶叶香气和滋味。采摘后的茶鲜叶施加15 ℃低温处理对CsNES基因表达水平和(E)-橙花叔醇含量的影响较单独采摘造成的机械损伤更大[11,68]。茉莉酸内酯具有类似茉莉花的香气和果香味,是茶叶香气的重要成分之一。单一采摘产生的短暂性机械损伤、室温或低温胁迫对茶鲜叶中的茉莉酸内酯含量影响均不显著,而持续性机械损伤和低温胁迫协同短暂性机械损伤均能够通过诱导茶树LOΧ基因,特别是CsLOX1的表达,并催化茉莉酸内酯前体物质亚油酸和α-亚麻酸形成,从而促进茉莉酸内酯在茶叶中的积累或释放[68-69]。采后茶叶香气与胁迫响应的关系如图2所示。

图2 采后茶叶香气物质与胁迫响应关系Fig.2 Relationship between aroma components and stress response in postharvest tea leaves
4 茶叶香气物质响应胁迫的功能
4.1 改善茶叶香气品质
基因工程、传统育种和逆境响应利用是目前用于改良茶叶天然风味的主要方法,相比于基因工程和传统育种,逆境响应利用更加生态安全、短时有效。在茶鲜叶生长和加工过程中对茶叶进行施加模拟胁迫可以提高茶叶香气组分含量。
利用病虫害和环境因子胁迫来促进茶叶在采前自然生长阶段香气物质的合成与积累。茶园害虫取食能够诱导茶叶释放大量的挥发性香气物质,“东方美人”乌龙茶是典型的利用小绿叶蝉取食后的茶叶加工而成的茶制品,这一特殊工艺赋予了“东方美人”乌龙茶特有的挥发性单萜类化合物,并使其成品茶具有特殊的蜜香和花果香[4,9]。 在采前的茶叶自然生长阶段,遮荫处理和特定波长光辐射,如红光和蓝光均可显著提高茶叶内源香气挥发性物质的含量[56],中短期遮荫处理对茶叶香气的形成有一定的促进作用,但长期遮荫则会严重影响茶叶光合作用并抑制其营养生长加速生殖生长,从而导致茶叶内含物减少。特定波长光辐射比遮荫处理在保证茶叶产量和提升香气方面更加具有优势[57],单一波长光、发光二极管以其波长特异性好、能量转换效率高等优点被广泛用于大多数农作物的采前生长与采后加工生产中。已有报道指出,蓝光和红光能够显著促进包括茶鲜叶中VFADs、VPBs和VTs等大多数挥发性物质的形成与释放[57]。
加工工艺是影响茶叶采后形成茶叶香气的主要因素。在茶叶加工过程中,茶叶细胞状态、环境温度、湿度、光照条件、机械损伤等因素往往对茶叶香气的形成起着关键性作用。有研究指出,新采茶鲜叶在3.0 μmol/(m2·s) UV-B处理2 h后其β-葡萄糖苷水解酶和β-樱草糖苷水解酶的基因表达水平均被显著上调,并释放大量的葡萄糖苷态挥发性物质和樱草糖苷态挥发性物质[70];在萎凋过程中辅加黄光、橙光或红光照射处理显著影响茶叶香气成分的含量,具有改善茶叶香气和滋味的良好效果[67];在茶叶遭受机械损伤的同时施加低温胁迫可促进挥发性脂肪酸衍生物的形成与释放。因此,在传统加工工艺上,合理辅加模拟逆境胁迫能够在一定程度上丰富茶叶香气物质、提高茶叶香气品质。
4.2 增强茶叶病虫害防御
植物防御响应机制包括直接防御和间接防御。植物可以通过挥发性物质直接防御食叶动物和病原菌的侵害[71], 如大豆叶片遭受线虫取食会释放大量的α-法尼烯来阻止线虫的进一步伤害[72];香气物质单萜、倍半萜或叶片混合挥发性物质可通过降低多种鳞翅目雌虫的产卵率来阻止害虫对自身的侵害[73-74];茶炭疽病菌、茶云纹叶枯病菌、茶轮斑病菌和茶饼病菌等病原菌侵染茶树叶片时 可诱导茶叶中GBVs大量水解释放出游离态香气组分并可通过诱导病原菌细胞凋亡来降低感病率[38,44,49]。Chen Shenglong等[46]指出(E)-橙花叔醇对茶叶病虫害有着显著的正面调控作用,(E)-橙花叔醇通过触发MAPKs、WRKY、H2O2的激活、茉莉酸和脱落酸信号响应等植物防御的早期反应,诱导茶叶积累大量的具有广谱性抗病虫害的防御相关化学物质,从而增强茶树对小绿叶蝉和茶炭疽病等的防御能力。
间接防御主要是通过吸引害虫天敌来达到植物自我保护的目的。食叶害虫取食不仅能够诱导植物产生香气等挥发性物质,食叶害虫本身也会分泌一些化合物诱导因子,二者均可以通过引诱食叶害虫天敌间接防御害虫,以及激活植物体内或植物与植物间的信号传导[1,75]。当食叶害虫取食玉米叶片时,倍半萜合成酶基因TPS10的表达水平显著升高并释放大量倍半萜,这些倍半萜化合物往往对害虫天敌寄生蜂有着较好的引诱效果,从而实现吸引寄生蜂以抵抗食叶害虫的持续取食[71]。LOΧ途径是一种重要的间接防御途径,分别由AOS和HPL介导,LOΧ/AOS途径伴随茉莉酸的产生,LOΧ/HPL途径伴随C6醛、醇及其酯等一些绿叶挥发性物质的衍生。HPL不仅是一些挥发性物质形成的重要水解酶,并且在植物抗病和抗虫害方面扮演着重要角色,HPL催化形成的C6醛在抵抗植物抗食叶害虫、细菌和真菌毒素方面表现出积极的作用[43,76]。食叶害虫如茶尺蠖取食、低温胁迫、红蓝光和机械损伤等外部条件均能够激活茶叶内部的LOΧ途径,由此产生的香气挥发性物质在提升茶叶香气品质的同时也有利于增强茶叶自身的防御反应。
植物在胁迫条件下释放的挥发性物质除了具有间接或直接的自我保护作用外,还可以作为信号物质通过植物激素信号途径在同株未受损组织器官或邻近植物中引发防御反应[77]。Wang Χuewen等[41]研究发现,茶叶在遭受害虫取食造成的机械损伤状态下会释放大量α-法尼烯,并由此激活受损叶片抗虫相关基因如β-1,3-葡聚糖酶基因的表达来参与抗虫响应,此外,释放的α-法尼烯还可以作为信号物质激活邻近完整叶片的抗菌相关因子,通过诱导提升邻近叶片水杨酸的水平触发防御响应。 Jing Tingting等[78]研究发现,茶尺蠖侵袭茶树释放的(E)-4,8-二甲基-1,3,7-壬三烯可诱导邻近茶树的茉莉酸和茉莉酸-异亮氨酸衍生物积累并显著提高邻近茶树的虫害防御能力。茶叶病虫害对茶叶产量和品质可造成严重的负面影响,虽然这些胁迫对茶叶香气物质的形成以及茶叶病虫害防御有积极影响,但目前相关研究还在初始阶段,且茶园管理主要以病虫害的防治为主。因此,生物胁迫对改善茶叶香气和增强病虫害防御的转型利用还需进一步的研究。
5 结 语
综上所述,茶叶香气品质的形成与采前生长环境条件和采后加工过程息息相关。一方面,生物胁迫因素(如茶园病虫害)和非生物胁迫因素(如光照条件和温度等)对采前茶叶香气物质的形成与积累具有不同的影响;另一方面,加工工艺产生的非生物胁迫(如机械损伤等)对采后茶叶香气物质的形成影响显著。明确茶叶香气成分对胁迫因素的响应机制,不仅有利于优化升级加工工艺、定向改善茶叶特征香气,同时也有助于发展茶园生态管理、增强茶园病虫害防治。但目前关于胁迫响应对茶叶香气物质形成的机制研究相对较少,大多数茶叶香气物质响应胁迫的分子机制还不清晰。茶叶在采前和加工过程中通常受到双重或多重胁迫因素的共同影响,目前的研究主要集中在单因素胁迫响应,因此多重胁迫对茶叶香气物质的作用机制还需更加全面系统地研究。病虫害是茶树生长的劲敌,虽然近些年在生态防御方面取得了一定的进步,但化学杀虫剂的使用仍普遍存在,这导致了茶叶农药残留的问题,而明确茶叶香气物质对病虫害胁迫响应的关系可以为控制茶园病虫害提供一种环境友好型的管理策略。此外,随着人们食品安全意识的提高,有机食品逐渐成为消费主流。有机茶叶要求茶园全天候不使用任何农药和化肥,茶园病虫害以生态防御为主,茶叶香气物质等挥发性化合物具有积极的生态防御功能,因此研究茶叶香气物质和胁迫响应之间的关系与转化利用显得尤为重要。
猜你喜欢
分子催化(2022年1期)2022-11-02 07:11:20
广州化工(2021年19期)2021-10-25 14:03:02
皮革制作与环保科技(2020年14期)2020-03-17 07:16:06
中国调味品(2017年2期)2017-03-20 16:18:22
当代化工研究(2016年5期)2016-03-20 16:21:35
广西林业科学(2016年2期)2016-03-20 05:53:43
食品界(2016年4期)2016-02-27 07:36:47
物理化学学报(2015年7期)2015-12-30 12:13:08
经济林研究(2015年3期)2015-12-21 01:23:25
中学科技(2015年3期)2015-04-29 04:52:43
