
设施菜地种植年限对土壤氨氧化微生物丰度和群落结构的影响
2021-08-25 16:26王喜英卢志宏谭智勇
中国蔬菜 2021年8期
赵 辉 王喜英 卢志宏 谭智勇
(铜仁学院经济管理学院,贵州铜仁 554300)
设施蔬菜是现代农业集约化发展的一个重要分支,具有集约化程度高、收益大等特点,在许多地区已经成为支柱产业(王学霞 等,2018)。截止到2018 年,我国设施蔬菜面积达到了400 万hm2(6 000万亩)左右,位居世界第一,占蔬菜种植总面积的19.1%(王学霞 等,2021)。然而,设施蔬菜具有种植指数高、农业投入(如杀菌剂、化肥、塑料薄膜)大、封闭或半封闭环境导致棚内温湿度较高且无雨水淋溶等特点。因此,设施蔬菜长期种植已经引起了人们对土壤质量退化、土壤和蔬菜潜在污染以及对人类健康的负面影响的担忧(宁德富 等,2016;Sun et al.,2016)。氮(N)是作物生长的主要限制因子,土壤氮有效性在决定作物氮素吸收和产量方面起着重要作用(李生秀,2008)。鉴于氮素对作物生长的显著贡献,设施栽培中大量氮肥被施入土壤,氮肥大量施用虽然提高了设施蔬菜的产量,但也造成了土壤养分氮的大量积累,加剧了土壤次生盐渍化和酸化,导致病原菌不断积累,土壤质量逐渐退化,温室气体排放加剧等问题,蔬菜品质下降,严重制约了设施蔬菜高效可持续发展(王倩姿,2019)。
硝化作用是自然界土壤氮素循环中的一个重要环节,土壤氨态氮在微生物作用下转化为亚硝酸盐氮()和硝酸盐氮()的过程,同时连接固氮作用、氨化作用和反硝化作用,直接影响氮素利用和环境质量(Vitousek et al.,1997)。硝化过程可以分为两步完成,第一步为氨态氮在亚硝化微生物下氧化成亚硝酸盐氮()的过程,也称为氨氧化过程;第二步为亚硝酸盐氮在亚硝酸盐氧化酶下氧化为硝酸盐氮()。在硝化过程中,亚硝态氮不稳定,易被氧化为硝态氮;因此,氨氧化过程的限速步骤,主要由氨氧化细菌(AOB)和氨氧化古菌(AOA)催化完成。研究表明,土壤pH、水分、有机质含量、施氮量和土地利用方式等对AOB、AOA 数量和群落结构有影响(Chen et al.,2014;Zhong et al.,2016)。可知,随着设施蔬菜种植年限的延长,土壤理化性质的变化将不可避免地导致硝化微生物数量和群落结构发生变化(方明 等,2019)。
前人研究认为,在氮充足的草地和农田生态系统中,硝化作用主要由AOB 驱动(Di et al.,2009;Jia & Conrad,2009)。Gubry-Rangin 等(2010)研究认为,在酸性低营养农田土壤中,硝化作用主要由AOA 驱动。Zhong 等(2016)研究表明,设施蔬菜土壤中AOB 和AOA 数量随施氮量增加呈现下降趋势,且AOB 群落结构变化较大。然而,关于设施蔬菜长期种植对土壤硝化微生物数量和群落结构的研究鲜见报道。
为此,本试验运用Illumina MiSeq 高通量测序技术和荧光定量PCR 技术对不同种植年限设施菜地土壤硝化微生物数量和群落结构进行研究,以揭示硝化微生物群落结构和丰度随设施种植年限的变化规律及其主要驱动环境因子,从硝化微生物的角度来解析土壤氮素利用效率,定向调控土壤氮素转化过程,为实现设施栽培蔬菜产业的可持续发展提供科学依据。
1 材料与方法
1.1 试验地点
试验在贵州省铜仁市碧江区和平乡农业园区(109°07′44″E,27°46′46″N)进行。试验点属于亚热带季风气候,年均温18 ℃,年降水量1 313 mm,土壤类型为黄壤。
1.2 试验设计
2017 年7 月在和平乡农业园区日光温室基地选取种植年限分别为3、5、7 a 的设施蔬菜大棚各3 个,大棚长40 m,宽8 m,以周围种植的露地蔬菜为对照(CK)。每个设施种植年限设置3 个重复,即每个大棚为1 个重复,露地蔬菜(CK)也设置3个重复。设施大棚每年蔬菜种植类型一致,轮作茄子、黄瓜、西葫芦、豇豆等,每年种植2 茬。设施蔬菜基肥长期施用氮磷钾复合肥1 200~1 500 kg ·hm-2,追施氮肥600~750 kg·hm-2。露地蔬菜主要种植茄子、南瓜、豇豆等,长期施用氮磷钾复合肥600~750 kg·hm-2,追施氮肥300~450 kg·hm-2。在整个试验处理中,除种植年限差异外,其余生产管理措施一致。
1.3 土样采集
在各样地内按“S”形(5 点法)采集0~10 cm 土层土壤样品,混合成1 个土样,用低温冰盒保存并迅速带回实验室。土样在室内去除石块和植物根系并过2 mm 筛后,分为2 份,一份-80 ℃冰箱保存,用于amoA基因群落结构和丰度分析;一份新鲜土壤用于铵态氮和硝态氮含量测定,余下部分室内风干过筛用于土壤化学指标测定。
1.4 测定项目
1.4.1 土壤化学指标测定 土壤化学性质采用鲍士旦(2000)的方法进行测定:土壤pH 值采用电位法测定,有机碳(SOC)含量采用重铬酸钾氧化法测定,全氮(TN)含量采用凯氏定氮法测定,铵态氮含量采用靛酚蓝比色法测定,硝态氮含量采用酚二磺酸比色法测定。
1.4.2 土壤DNA 提取及荧光定量PCR 称取0.5 g 土壤,按照E.Z.N.A.®Soil DNA Kit(Omega,GA,USA)试剂盒操作步骤提取土壤DNA。用1%琼脂糖凝胶电泳检测DNA 的完整性,用核酸定量仪(Nanodrop-NC2000)检测DNA 浓度和纯度。PCR 产物纯化回收后,将其连接至pMD18-T 载体,转化至大肠杆菌DH5α 感受态中进行培养,筛选阳性克隆,提取amoA基因重组质粒,质粒浓度经核酸定量仪测定后,计算基因拷贝数,按照10倍梯度稀释至103~108拷贝数,制备标准曲线。荧光定量PCR 反应在ABI7500 荧光定量PCR 仪上进行,反应体系为20 μL:10 μL 2×SYBR Green,上下游引物(10 μmol·L-1)各0.5 μL,ROX 0.5 μL,DNA 模板2 μL(1~10 ng),最后用ddH2O补至20 μL。荧光定量PCR 反应条件为:95 ℃ 30 s;95℃ 5 s,60 ℃ 40 s,72 ℃ 30 s,40 个循环。
1.4.3 高通量测定 AOA 扩增引物为amoA-23F(5′-ATGGTCTGGYTWAGACG-3′)和amoA-616R(5′-GCCATCCATCTGTATGTCCA-3′)。AOB 扩增引物为amoA-1F(5′-GGGGTTTCTACTGGTGG T-3′)、amoA-2R(5′-CCCCTCKGSAAAGC CTTCTTC-3′)(Wessén et al.,2011)。PCR体系25 μL:5×Ex Taq缓冲液5.0 μL,dNTP(2.5 μmol ·L-1)2.0 μL,上下游引物(10 μmol·L-1)各1.0 μL,DNA 模 板2.0 μL(1~10 ng),Ex Taq(5 U ·μL-1)0.25 μL,最后用ddH2O 补至25 μL。PCR 反应条件为:95 ℃ 5 min;95 ℃ 30 s,55 ℃ 30 s,72℃ 1 min,27 个循环;72 ℃ 10 min。以2%琼脂糖凝胶电泳检测PCR 产物。样品送至上海派森诺生物科技股份有限公司,运用Illumina MiSeq 测序平台进行测序。测序样品数据下机后,根据barcode标签序列和前引物序列筛选出有效序列,去除接头和barcode 序列,利用FLASH 软件对通过质量初筛的双端序列根据重叠碱基进行配对连接,同时利用USEARCH 软件检查并剔除嵌合体序列,从而获得高质量有效序列。使用QIIME 软件调用UCLUST 序列比对工具,按照97%的序列相似度进行OTU(Operational Taxonomic Unit)划分和归并,并选取丰度最高的序列作为该OTU 的代表序列。利用QIIME 软件将OTU 的代表序列与功能基因数据库(FunGene)进行比对,获取每个OTU 对应的分类学信息。
1.5 数据处理
采用SPSS 21.0 软件进行土壤化学性质、氨氧化微生物α 多样性指数、丰度和群落组成相对丰度的差异显著性分析(P<0.05)和相关性分析。采用R 软件进行主成分分析(principal component analysis,PCA)和冗余分析(redundancy analysis,RDA)。
2 结果与分析
2.1 种植年限对土壤化学性质的影响
由表1 可知,设施蔬菜长期种植对土壤理化性质有显著影响(P<0.05)。不同设施菜地种植年限土壤pH 值、有机碳含量和碳氮比均显著小于CK,且随种植年限延长逐渐降低。种植3、5、7 a土壤全氮、铵态氮和硝态氮含量均高于CK,且随种植年限延长逐渐增加。由此可知,设施蔬菜长期种植土壤酸性增强,土壤铵态氮和硝态氮含量积累。

表1 不同种植年限设施菜地土壤理化性质
2.2 种植年限对氨氧化菌amoA 基因丰度的影响
由图1 可知,不同设施菜地种植年限土壤AOA-amoA和AOB-amoA基因拷贝数变化范围分别为2.02 × 107~3.28 × 107个·g-1(干土)和0.53 ×106~1.91 × 106个·g-1(干土)。AOA-amoA拷贝数随种植年限延长逐渐下降,种植3 a 和5 a 处理土壤AOA-amoA拷贝数均显著高于7 a 和CK(P<0.05),其中种植3 a 和5 a 处理之间差异不显著;种植7 a 土壤AOA-amoA拷贝数低于CK,但差异不显著(P>0.05)。AOA-amoA和AOB-amoA拷贝数在各处理中变化趋势相同,且AOA-amoA拷贝数在不同处理中均高于AOB-amoA。AOA/AOB值为17.33~39.16,大小顺序表现为CK >7 a >5 a >3 a,且7 a 处理分别与3 a 和5 a 处理间差异显著(P<0.05)。
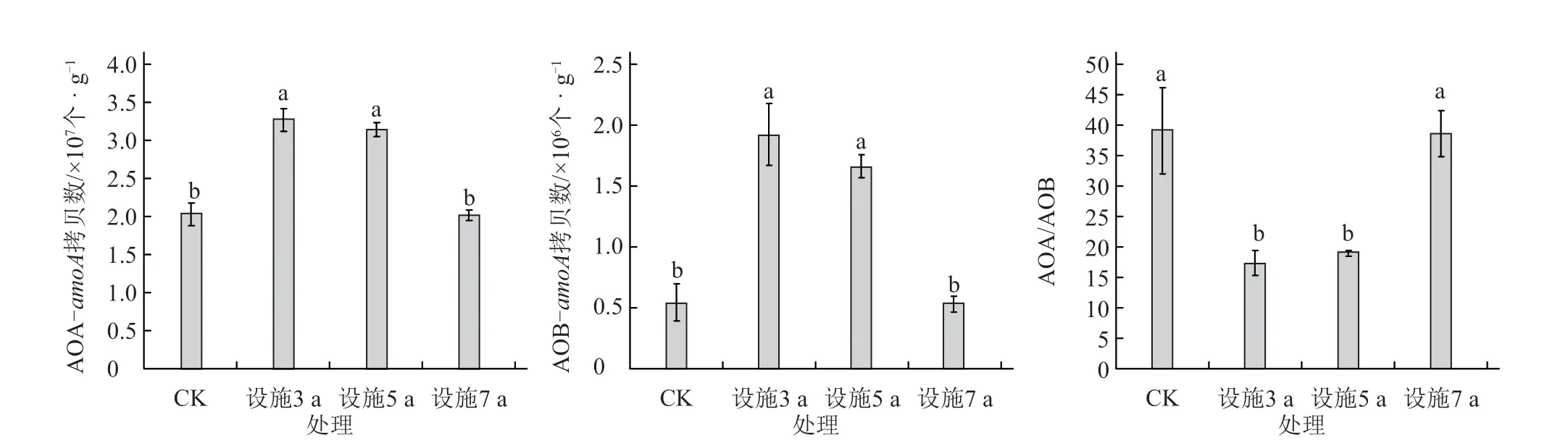
图1 不同种植年限设施土壤AOA-amoA 和AOB-amoA 基因丰度及AOA/AOB
2.3 种植年限对氨氧化微生物α 多样性的影响
种植年限对AOA 群落的Chao1 指数、ACE指数和Simpson 指数有极显著影响(P<0.01);对Shannon 指数有显著影响(P<0.05)(表2)。Chao1 指数和ACE 指数范围分别为97.65~221.13和98.80~228.65,其中种植3 a 处理显著高于其他处理。Shannon 指数和Simpson 指数范围为2.87~4.26 和0.83~0.94,大小顺序为3 a >5 a >CK >7 a(表3)。
种植年限对AOB 群落的ACE 指数和Simpson指数有显著影响(P<0.05)(表2)。Chao1 指数 和ACE指数范围分别为192.15~415.53 和195.63~424.68,其中3 a 和5 a 处理分别是CK 的2.16、2.03 倍和2.17、2.04 倍。Shannon 指数范围为3.38~4.60,3 a 和5 a 处理分别是CK 的1.11 倍和1.08 倍。Simpson 指数范围为0.68~0.88,大小顺序为5 a >CK >3 a >7 a,7 a 分别与CK、3 a和5 a 处理间差异显著(表3)。

表2 设施栽培年限对AOA 和AOB 多样性指数的影响方差分析

表3 不同种植年限设施土壤AOA 和AOB 群落ɑ 多样性指数的影响
2.4 种植年限对AOA 和AOB 群落的影响
在门水平上,AOA 群落由奇古菌门(Thaumarchaeota)、变形菌门(Proteobacteria)、unidentified 组成(图2-A)。奇古菌门为主要优势类群,各处理相对丰度分别为55.03%~71.90%;其中种植3 a 处理相对丰度为55.06%,极显著低于其他处理。变形菌门相对丰度为6.36%~24.26%;其中种植3 a 处理最高,随设施种植年限延长逐渐减低,且处理间差异显著。在属水平上,共检测到6个类群(图2-B),分别为Candidatus Nitrosotalea、亚硝化暖菌属(Nitrososphaera)、亚硝化单胞菌(Nitrosomonas)、Archaea-unidentified、硝化螺旋菌属(Nitrospira)和气单胞菌属(Aeromonas)。其中Candidatus Nitrosotalea、亚硝化暖菌属和亚硝化单胞菌属为优势类群,各处理相对丰度分 别 为30.23%~58.76%、12.12%~33.54% 和3.68%~18.91%。Candidatus Nitrosotalea相对丰度大小顺序为CK >5 a >3 a >7 a,说明设施年限延长对其有抑制作用。亚硝化暖菌属、亚硝化单胞菌属相对丰度在3、5 a 和7 a 处理中均显著高于CK,说明设施栽培对其有促进作用。硝化螺旋菌属在5 a 处理中相对丰度为5.33%,高于其他处理,但各处理间差异不显著。Archaea-unidentified在CK 中相对丰度为11.48%,高于其他处理。气单胞菌属在5 a 处理中相对丰度最低,但与其他处理间差异不显著。
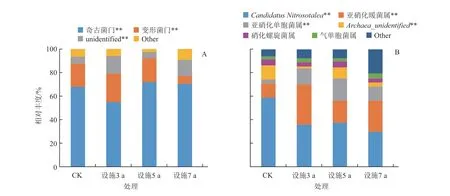
图2 设施种植年限土壤氨氧化古菌门和属水平组成
AOB 群落在门水平上共获得3 个类群(图3-A),分别为变形菌门(Proteobacteria)、奇古菌门(Thaumarchaeota)和硝化螺旋菌门(Nitrospirae)。变形菌门为主要优势类群,相对丰度范围为73.57%~87.55%,3 a 和5 a 处理分别是CK 的1.12倍和1.03 倍。奇古菌门相对丰度在CK 中最低,随设施种植年限延长呈先增加后降低趋势。硝化螺旋菌门相对丰度在CK 最高,随设施种植年限延长表现出先降低后增加趋势。在属水平上,共检测到5 个类群(图3-B),分别为亚硝化螺菌属(Nitrosospira)、Candidatus Nitrosotalea、亚硝化单胞菌属(Nitrosomonas)、硝化螺旋菌属(Nitrospira)和亚硝化暖菌属(Nitrososphaer)。亚硝化螺菌属为优势类群,相对丰度范围为77.21%~86.55%。亚硝化螺菌属的相对丰度在3 a 和5 a 处理中较高,分别是CK 的1.10 倍和1.05 倍,随设施种植年限延长逐渐降低。Candidatus Nitrosotalea相对丰度在7 a 处理中最低,显著低于其他处理。亚硝化单胞菌属和硝化螺旋菌属相对丰度在CK 中最高。亚硝化暖菌属相对丰度在3 a 处理最高,随设施种植年限延长表现出先减少后增加的趋势。

图3 不同种植年限设施土壤氨氧化细菌门和属水平组成
2.5 氨氧化微生物群落结构及其与土壤化学性质的关系
通过对AOA 和AOB 群落进行主成分分析可知(图4),不同设施种植年限土壤AOA 和AOB群落结构差异明显。AOA 的PC1 和PC2 分别为56.79%和18.60%,二者累计贡献率达到75.39%。3、5 a 和7 a 处理分别与CK 在PC1 和PC2 上分离都较大,说明设施蔬菜种植年限导致AOA 群落结构变化较大;3 a 和7 a 处理相聚较近,群落相似度较大。AOB 的PC1 和PC2 分别为55.30%和27.38%。3、5 a 和7 a 处理在PC1 和PC2 上分离都较大,说明设施蔬菜种植年限对AOB 群落结构影响较大。总体看来,种植年限对AOA 群落的影响强于AOB。
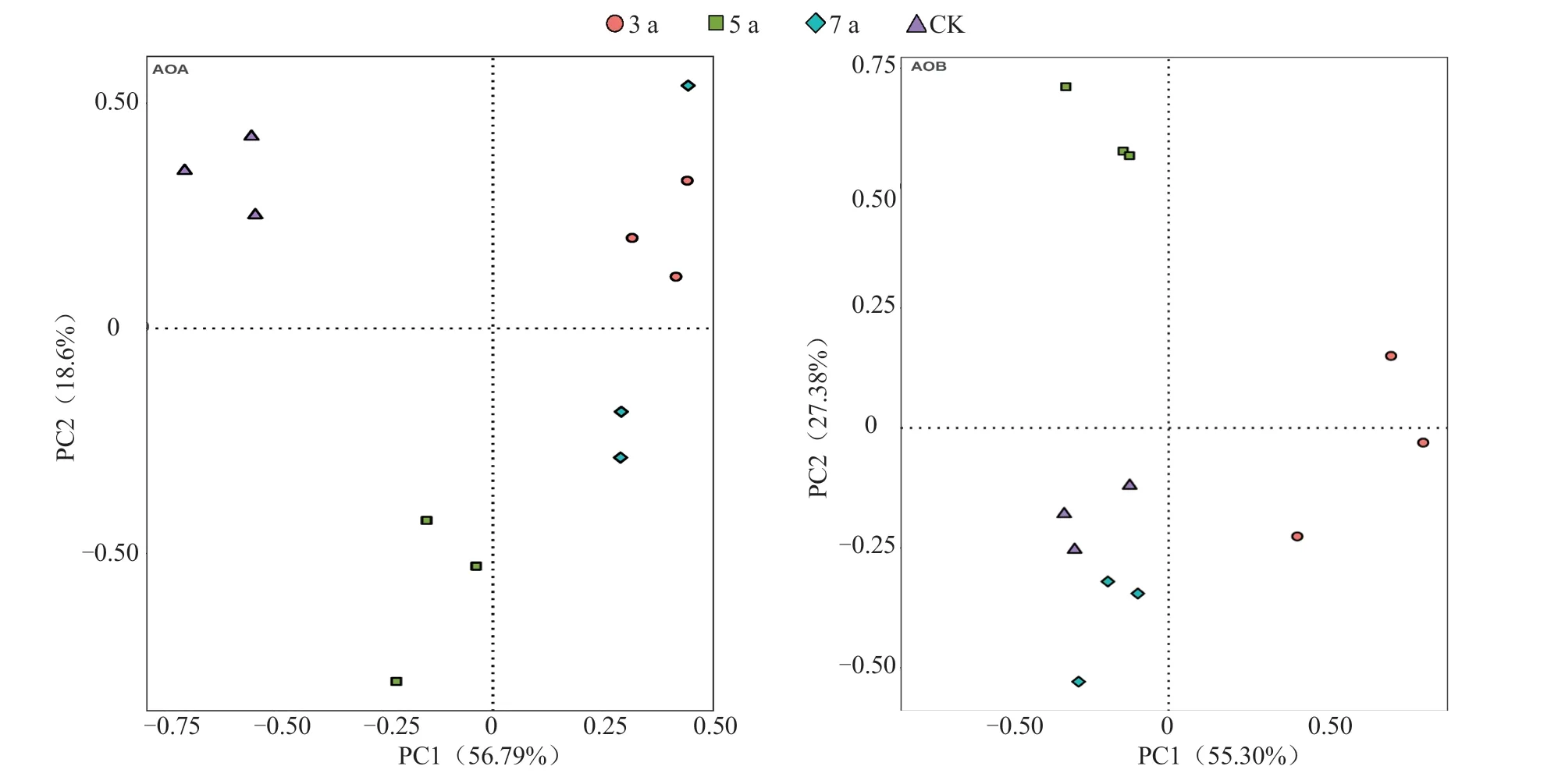
图4 不同种植年限设施土壤AOA 和AOB 群落主成分分析
RDA 反应基于属分类水平上土壤化学性质对AOA 和AOB 群落结构的影响(图5)。AOA 的RDA1 和RDA2 轴分别为61.29%和19.59%。土壤pH 对AOA 群落结构有显著影响(P<0.05),铵态氮和硝态氮对AOA 群落结构有极显著影响(P<0.01)。AOB 的RDA1 和RDA2 轴 分 别 为39.64%和25.24%。土壤铵态氮和硝态氮对AOB群落结构有显著影响(P<0.05)。

图5 AOA、AOB 与土壤化学性质的冗余分析
3 讨论
3.1 种植年限对设施土壤AOA 和AOB 数量的影响
AOA 和AOB 是氨氧化作用的主要承担者,在土壤氮转化过程中具有重要作用,影响植物对氮素的吸收利用,且与过量施肥导致土壤酸化、硝酸盐淋失和温室气体排放等关系密切(Wang et al.,2017)。本试验中不同处理土壤AOA 数量高于AOB,AOA/AOB 值为17.33~39.16,与其他研究者结果相符(Xu et al.,2012;张苗苗 等,2015),进一步证实了酸性土壤中AOA 占主导优势(杨亚东 等,2017)。设施蔬菜土壤中AOA 和AOB 数量均高于CK,可能与设施土壤氮含量较高有关。Di等(2010)研究表明,无机氮作为氨氧化细菌的能量来源,可促进土壤氨氧化细菌生长。
相关研究表明,AOA 和AOB 数量受多种土壤环境因子影响(Yao et al.,2013)。Pernes-Debuyser和Tessier(2004)研究认为,长期氮肥添加通过氨氧化作用可导致土壤中质子积累,从而降低土壤pH。Chen 等(2013)研究认为,土壤pH 与AOB数量显著负相关,与AOA 数量没有显著相关关系。然而,杨亚东等(2017)研究认为,土壤pH 与AOA 数量显著正相关,与AOB 数量没有显著相关关系。Chen 等(2011)研究也认为,土壤pH 是影响AOA 种群数量变化的主要因素,较低pH 不适合AOB 生长。AOB 利用的底物为氨分子(NH3),土壤pH 降低将导致NH3转变为,导致底物NH3浓度降低。本试验中,随种植年限延长,土壤pH 值逐渐降低,驱动NH3转化为-N,可能是造成AOB 数量减少的主要因素(Macqueen & Gubry-Rangin,2016)。然而,AOA 对底物氨分子(NH3)的亲和力强于AOB(Willm et al.,2009),且适应较低pH 环境(He et al.,2012)。由此可知,在设施蔬菜长期种植中AOA 可能在氨氧化过程发挥重要作用。
然而,本试验中AOA 和AOB 数量级低于杨亚东(2018)在农田土壤的研究结果,设施蔬菜长期种植降低了AOA 和AOB 数量。AOA 和AOB 数量变化趋势一致,随设施种植年限延长逐渐降低,均高于CK。AOA 数量在不同种植年限中有显著变化,与陈秋会(2014)研究结果相反。Shen 等(2011)通过研究不同氮水平下我国菜地土壤AOB 数量的变化,发现氮肥显著提高菜地土壤AOB 数量。以上研究结果的差异,可能与研究区域施肥组成、施肥水平、取样时间和立地条件等因素有关。由此可知,AOA 和AOB 数量对不同土壤环境因子的差异响应,将导致AOA 和AOB 生态位发生分离。
3.2 AOA 和AOB 群落及其与土壤化学性质的关系
种植年限3 a 和5 a 的AOA 和AOB 群落丰富度和多样性指数均高于CK,但种植7 a 的AOA 和AOB 群落丰富度和多样性指数低于CK。由此可知,适宜种植年限有利于增加AOA 和AOB 群落多样性,长期种植将降低AOA和AOB群落多样性;可能由于长期施用氮肥,特别是铵态氮是引起土壤AOA 和AOB 群落结构发生变化的主要因素(Willm et al.,2009)。本试验中,种植年限对AOA 群落的影响最大,与Chen 等(2013)的研究结果相反。然而,Wang 等(2009)和Shen 等(2011)分别在水稻土壤和半干旱温带草原土壤中发现,施氮对AOA 群落组成没有显著影响。研究结果的差异原因可能包含:①本试验对象为设施蔬菜,具有独特的生态环境,不同于农田(水稻)和草原生态系统。② AOA 生长极其缓慢,需要经过长期处理后才可检测到变化(Könneke et al.,2005)。Wang 等(2009)仅在施氮89 d 后就开始对AOA 数量和群落结构进行检测,而本试验是设施蔬菜种植多年后才对其进行检测。
在门分类水平上,AOA 群落中奇古菌门为优势类群,与杜颖等(2014)研究表明浑善达克沙地的优势类群结果一致。与种植3 a 相比,种植5 a和7 a 显著增加了奇古菌门的相对丰度,表明种植年限对其影响较大。AOB 群落中变形菌门的相对丰度占比较高,平均相对丰度达到整个AOB 群落门水平的79.89%。种植3 a 和5 a 的AOA 和AOB群落的变形菌门相对丰度高于7 a 处理,进一步证实了变形菌门具有嗜营养的特点,但有其生态阈值(Fazi et al.,2005);可知,随设施蔬菜种植年限延长,土壤养分过量富集,超过变形菌门需要的阈值,将对其有抑制作用。土壤pH、铵态氮和硝态氮是土壤AOA 群落结构的主要因素,与杨亚东(2018)研究结果一致。说明设施蔬菜种植过程中,土壤酸化和氮肥积累直接影响土壤AOA 群落变化。
土壤可利用性氮直接影响AOB 群落结构变化(Hynes & Germida,2012)。土壤铵态氮和硝态氮对AOB 群落结构影响较大,说明设施栽培通过改变铵态氮和硝态氮等指标直接或者间接来影响AOB 群落组成结构变化。为准确评价设施蔬菜种植中土壤氨氧化作用发生规律,需要进一步开展长期的试验研究。
4 结论
本试验结果表明,AOA 和AOB 数量、群落的Chao1 指数、ACE 指数和Shannon 指数,随设施种植年限延长逐渐降低。门水平上,奇古菌门和变形菌门分别为AOA、AOB 群落的优势类群。设施蔬菜长期种植中,土壤pH 是一个重要指标,土壤pH 与AOA 群落结构有显著关系,低pH 值可限制硝化微生物生长,不利于硝化过程进行。在设施蔬菜酸性土壤中,硝化作用可能主要由AOA 驱动完成。
猜你喜欢
土壤学报(2022年2期)2022-06-09
水土保持学报(2022年3期)2022-05-26
建材发展导向(2022年6期)2022-04-18
河南师范大学学报(自然科学版)(2022年3期)2022-04-14
科技创新与应用(2022年6期)2022-03-24
中国生态农业学报(中英文)(2022年1期)2022-01-15
环境与发展(2019年12期)2019-02-12
江苏农业科学(2016年10期)2017-02-05
江苏农业科学(2016年5期)2016-07-23
农家顾问(2016年5期)2016-05-14
