
蛋白质组学技术解析急性免疫应激影响肉仔鸡肉品质的机理
2021-08-23 07:10张安荣吴正可陈志敏常文环蔡辉益刘国华郑爱娟
畜牧兽医学报 2021年8期
张安荣,吴正可,陈志敏,常文环,蔡辉益,2,刘国华,郑爱娟*
(1.中国农业科学院饲料研究所 农业农村部饲料生物技术重点开放实验室,北京 100081;2.生物饲料开发国家工程研究中心,北京 100081)
在现代化畜牧业发展过程中,集约化养殖早已被视作高效生产、优质产品的保证,但大规模的集中饲养模式所暴露的问题也日益突显。集约化饲养导致肉鸡长期处于高密度、多因素应激源的环境中,例如高温、惊吓、争食、运输、季节变化、屠宰等诸多因素[1-2],对动物生理和心理健康都造成不同程度的影响。其中,免疫应激会导致机体营养代谢消耗增加、生长发育受阻以及肉品质下降,严重时还会引发各种疾病,给家禽业带来严重的经济损失。研究表明,LPS诱导的免疫应激不但降低肉鸡生产性能、改变血液指标[3]、损害肉鸡肠道结构的完整性,还会促进肠黏膜核因子κB、白介素IL-1β、IL-6、肿瘤坏死因子α等促炎细胞因子积累,最终造成炎症的发生[4-6]。同时,免疫应激也会引起肉鸡机体氧化损伤和抗氧化机能下降,进而导致肉鸡肉品质显著下降。无论是免疫亢进还是免疫抑制,均会影响宰后鸡肉的品质[7]。目前,对肉鸡抗免疫应激的营养调控,主要是通过在日粮中添加硒[8]、维生素E[9]、丁酸钠[10]、α-生育酚琥珀酸酯[11]和抗菌肽[12]来缓解损伤,但是其效果并不理想,其原因是,免疫应激影响宰后鸡肉品质的分子机理仍不清楚。
蛋白质是生命活动的物质基础和生理功能的直接参与者和执行者,生物机体的生理变化通常是由多个蛋白协同或是作为功能整体来完成[13]。蛋白质组学是实现对基因组所编码的全部蛋白质及其相互作用关系的研究,对有机体或细胞中的蛋白质进行全方位研究,以整体的视角阐述生命现象的本质和活动规律。近年来,以质谱和生物信息学为技术系统的蛋白质组学以其高分辨率、高灵敏度、高通量和规模化的优势,广泛应用于动物营养原理与分子调控机制的研究。尤其是在肉品质研究方面,该技术主要用于研究肉品质形成的机制、肉品质的改善和肉质检测等方面。付睿琦[14]利用蛋白质组学技术研究了鸡生长期胸肌蛋白表达谱,结果证实,随着周龄的增长,差异蛋白数减少,但是与出雏时相比,差异蛋白数达总蛋白的2/3。刘志红等[15]通过蛋白质组学技术对肉鸡胸肌和腿肌进行了蛋白质鉴定和定量分析,发现胸肌与腿肌间共有98个差异表达蛋白,并且腿肌中高表达蛋白多富集于运动相关功能,而胸肌中高表达蛋白则富集于肌肉收缩相关功能。彭梦玲等[16]研究了肉鸡胚胎发育过程中肝组织中蛋白表达的变化,发现14胚龄和出壳1日龄肉鸡肝出现10个核心差异蛋白,分别调控脂肪酸降解和糖异生生物过程。邹波等[17]研究了屠宰前不同驱赶方式对生猪造成应激的影响,通过蛋白质组学分析发现,与能量代谢和肌肉收缩相关的46个蛋白发生差异化表达。Jia等[18]研究发现,在活体和新鲜屠宰后的牛胸部嫩肉中有大量蛋白质Peroxiredoxin-6存在,并建议将此蛋白作为检测肉质嫩度的指标蛋白。
因此,本试验采用蛋白质组学技术研究免疫应激对肉仔鸡胸肌肉品质和蛋白质组的影响,解析免疫应激导致肉仔鸡胸肌物质代谢变化的分子机理,为合理制定缓解生产中免疫应激措施和提高肉鸡生产水平提供理论依据。
1 材料与方法
1.1 主要仪器设备和试剂
主要仪器设备:组织破碎仪、探头超声仪(中国南京电子计量仪器公司);纳升级高效液相色谱系统(Q-Exactive,美国Thermo Fisher Scientific公司);质谱仪、蛋白定量分光光度计(美国Thermo Fisher Scientific公司);台式低温离心机(美国Beckman Coulter公司);低温蒸发浓缩仪(德国Marin Christ公司);Milli-QGradient超纯水机(美国Millipore公司);超低温冰箱(日本SANYO公司);pH计和电子分析天平(德国Mettler Toledo公司)。
主要试剂:尿素(Solarbio公司);硫脲、CHAPS、Tris碱、二硫苏糖醇(Amresco公司);碘乙酰胺(Merk公司);丙酮、三氟乙酸(J.T.Baker公司);蛋白酶抑制剂(Roche Base公司);Bradford法蛋白质定量试剂盒(普利莱公司);碳酸氢铵(Sigma Aldrich公司);Trypsin酶(Promega);甲酸(MREDA Technology公司);乙腈(Fisher公司);脂多糖(大肠杆菌血清型O55:B5,Sigma公司)。
1.2 试验动物及试验设计
健康1日龄AA肉公鸡购自北京正大有限公司,饲喂玉米-豆粕型饲粮(表1),随机分为两组,即对照组和免疫应激组,每组6个重复,每个重复10只鸡。对照组和免疫应激组分别于36、38、40日龄,腹腔分别注射1 mL生理盐水(对照组)或5.0 mg·kg-1体重的LPS溶液(免疫应激组)。在42日龄,每个重复随机取2只鸡,采集胸肌组织样本,-80 ℃冰箱保存备用。
1.3 指标测定
1.3.1 蒸煮损失的测定 将样品沿着与肌肉自然走向(肌肉的轴)垂直的方向切成2.5 cm厚的肉块,并去除样品表面的结缔组织、脂肪和肌膜,使其表面平整。记录肉样的初始重量。将肉块放入塑料蒸煮袋中,将温度计探头由上而下插入肉块中心,记录肉块的初始温度,将蒸煮袋口密封起来。将包装的肉块放入72 ℃水浴中,水浴液面需完全浸没肉块,袋口不得浸入水中。当肉块中心温度达到70 ℃时,立即取出肉样,冷却称重。肉块蒸煮前后的质量损失即为蒸煮损失。

表1 基础饲粮组成及营养水平(风干基础)
1.3.2 pH测定 测定肌肉pH的样品取自肉鸡右胸肌。将校准后的便携式pH计金属头直接插入肉样中,待pH计读数稳定后,读取数值。每个样品测3次,取平均值。在屠宰后45 min记录初始pH(pHi),于冷却24 h后再次记录pH(pH24 h),于48 h 后记录最终pH(pH48 h)[19]。
1.3.3 滴水损失的测定 测定肌肉滴水损失的样品取自肉鸡左胸肌。记录肉样的初始重量,然后将样品放入密封的真空聚乙烯袋中,并在4 ℃下储存。24 h后,将样品从袋子中取出,用吸水纸吸干,并重新称重[20]。肉块处理前后的质量损失即为滴水损失。
1.3.4 IMP含量的测定 参照Li等[21]的方法,通过高效液相色谱(HLPC)测定肉鸡胸肌IMP含量。
1.3.5 胸肌肌纤维横截面周长与面积的测定 在ImageJ分析软件下,打开已采集的不同处理胸肌肌纤维横截面染色图像。在image菜单中选择Type为8bit,在plugins中选择demos为inverter,在image下adjust中选择threshed调节需要测量的区域,在analyze下的tools选项中选择ROI manager,然后用选图工具勾勒出测定区域并加入至ROI manager待测框中。将视野中的肌纤维横截面选中后点击measure选项获得测定数据,测得胸肌肌纤维横截面积与周长。
1.4 胸肌组织蛋白质的提取与酶解
1.4.1 胸肌组织蛋白质提取 蛋白质提取按照Zheng等[22]的方法,每30 mg胸肌样品加入300 μL样品裂解液(8 mol·L-1尿素,2 mol·L-1硫脲,4% CHAPS,20 mmol·L-1Tris-碱,30 mmol·L-1二硫苏糖醇(DTT))及蛋白酶抑制剂,冰浴充分研磨,超声,使样品完全溶解。4 ℃,15 000×g,离心20 min。 避开脂肪层取上清液,加入3倍体积预冷的丙酮溶液,冰浴沉淀30 min。4 ℃,15 000×g,离心20 min。弃上清。开口2~3 min让丙酮挥发,然后将沉淀重新溶解于100 μL的5 mol·L-1尿素中,再加入400 μL 40 mmol·L-1NH4HCO3。采用Bradford方法测定所提取蛋白质的浓度。
1.4.2 胸肌组织蛋白质液内酶解 每个样品加入50 μL 100 mmol·L- 1DTT,4 ℃放置1 h,加入250 μL 100 mmol·L-1碘乙酰胺(IAA),避光放置1 h。按照酶∶蛋白=1∶50(W/W)比例加入Trypsin胰蛋白酶,37 ℃下进行消化反应24 h,后加入1 μL甲酸终止酶切。4 ℃,14 000×g,离心15 min,取500 μL上清液转移至新管,待测。
1.5 胸肌样品质谱分析
用100 μL 0.1%甲酸溶解干燥样品,4 ℃,14 000×g离心15 min,取50 μL上清于上样管中,同时避免有气泡;将上样管放入色谱仪样品槽,每针上样体积8 μL,每个样品做3针重复。采用纳升级液相色谱系统EASY-nLC 1000,通过纳升电喷雾源与质谱Q-Exactive串联,通过纳升ESI源将洗脱的肽段注入质谱仪,上样流速250 nL·min-1。流动相A(0.1%甲酸和2%乙腈),流动相 B(0.1%甲酸 & 80% ACN);梯度:3%~8% B 10 min,8%~12% B 100 min,12%~20% B 70 min,20%~30% B 30 min,30%~90% B 20 min和90% B 10 min。
参数设置如下:喷雾电压:2.3 kV;毛细管温度:250 ℃;S-lens:55%;碰撞能量:27% HCD;分辨率设置:一级70 000@m/z 200;二级17 500@m/z 200;母离子扫描范围:m/z 300~2 000;子离子扫描范围:从m/z 100开始。通过Xcalibur软件(版本2.2,Thermo Fisher Scientific)收集MS/MS数据并保存为Raw文件。
1.6 胸肌组织全蛋白质组定性及定量分析
原始文件使用PEAKS软件进行Label-free定性和定量分析,搜索鸡种属蛋白质数据库(Gallusgallusdatabase)。检索结果通过严格标准进行筛选(peptide FDR & protein FDR≤1%)。数据库搜索时的参数设置:前体离子质量偏差:15 ppm;碎片离子质量偏差:20 mmu;固定修饰:半胱氨酸(Cysteine)烷基化(+57.021 u);动态修饰:甲硫氨酸(Methionine)氧化(+15.995 u);天冬酰胺和谷氨酰胺(Asparagine & Glutamine)脱氨基化(+0.984 u);酶:trypsin;漏切位点:2。
运用PEAKS Q板块对搜库结果进行定量,参数如下:保留时间(retentiontime shift tolerance):0.5 min;质量误差范围(mass error tolerance):30 ppm; 特有肽段(unique peptide)≥1;电荷范围(charge between):2~8;不同处理间蛋白质的表达量差异倍数(fold change)≥1.5,P≤0.05判定为差异显著。
1.7 蛋白质组生物信息学分析
利用Cytoscape3.8.0软件中的Cluego插件,分别用基因本体论数据库(Gene Ontology database,GO)和京都基因和基因组百科全书(Kyoto encyclopedia of genes and genomes,KEGG)数据库对差异蛋白质进行功能富集分析和代谢通路富集分析。使用检索相互作用基因/蛋白质(STRING)软件分析差异蛋白相互间的作用以筛选出核心差异因子。
1.8 试验数据统计分析
肉鸡胸肌肉品质部分数据以“平均值±标准差(mean±SD)”表示。采用SPSS 21.0软件中的独立样本t检验进行数据分析处理。以P<0.05为差异显著的判断标准,P<0.01为差异极显著的判断标准。
2 结 果
2.1 免疫应激对肉仔鸡胸肌肉品质的影响
免疫应激对肉鸡胸肌滴水损失、蒸煮损失和肌苷酸(IMP)含量的影响如表2所示。与对照组相比,免疫应激显著增加了宰后肉鸡胸肌的滴水损失及蒸煮损失(P<0.05),并且免疫应激组肉鸡胸肌IMP含量显著低于对照组(P<0.01)。表3显示,屠宰后24和48 h的肉鸡胸肌pH,与对照组相比,免疫应激对屠宰后24和48 h的肉鸡胸肌pH无显著影响(P>0.05)。如表4显示,与对照组相比,免疫应激组肉鸡胸肌纤维的横截面周长和面积均极显著增加(P<0.01)。

表2 免疫应激对肉鸡胸肌蒸煮损失、滴水损失及肌苷酸含量的影响

表3 免疫应激对宰后静置不同时间肉鸡胸肌pH的影响

表4 免疫应激对肉鸡胸肌肌纤维横截面的周长与面积的影响
2.2 肉鸡胸肌蛋白质组定性分析
2.2.1 肉鸡胸肌蛋白质鉴定 本试验在肉鸡胸肌组织中共鉴定到1 163个蛋白质,对照组表达702个蛋白质(共468个组),免疫应激组表达975个蛋白质(620组),对照组和免疫应激组共表达514个蛋白质,如图1所示。

图1 对照组和免疫应激组肉鸡胸肌表达蛋白质数目维恩图Fig.1 Venn diagram of the number of proteins expressed in the PM muscle of broilers in control group and immunological stress group
2.2.2 对照组特异表达蛋白质GO和KEGG分析 如图2所示,对对照组中特异表达的蛋白质进行了KEGG途径分析,结果显示,特异表达蛋白质未富集到任何通路。
GO分析显示,对照组特异表达蛋白质主要富集到了能量产生和代谢相关的生物学过程:细胞呼吸(cellular respiration)、有氧呼吸(aerobic respiration)、己糖代谢与生物合成过程(hexose metabolic and biosynthetic process)、单糖生物合成过程(monosaccharide biosynthetic process)、葡萄糖代谢过程(glucose metabolic process)、糖异生(gluconeogenesis)。
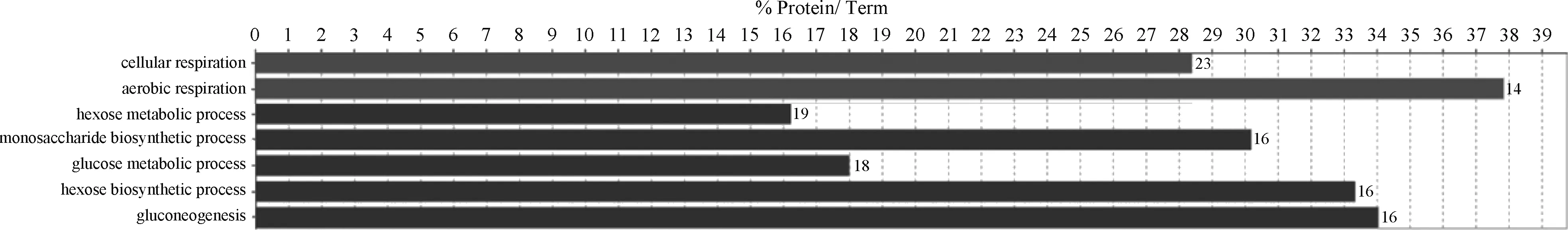
图2 对照组肉鸡胸肌特异表达蛋白质GO功能注释Fig.2 GO annotation of unique proteins specially expressed in the PM of broilers in the control group
2.2.3 免疫应激组特异表达蛋白质GO和KEGG分析 如图3所示,KEGG分析结果显示,免疫应激组特异表达蛋白质富集到了紧密连接(tight junction)通路。免疫应激组差异表达蛋白GO富集分析结果显示,所富集的GO条目主要涉及以下生物学过程:大分子复合物分解(macromolecular complex disassembly)、蛋白质分解与解聚(cellular & protein complex disassembly, protein depolymerization)、核酸合成和代谢(purine nucleoside & ribonucleoside triphosphate metabolic process, ATP metabolic process, nucleoside metabolic process, purine nucleoside & ribonucleoside monophosphate biosynthetic and metabolic process等)、糖基化合物合成与代谢(glycosyl compound biosynthetic and metabolic process)、含碱基小分子物质代谢(nucleobase-containing small molecule metabolic process)。
2.3 肉鸡胸肌蛋白质组定量分析
2.3.1 免疫应激组肉鸡胸肌差异表达蛋白质数目 定量分析结果表明,免疫应激组和对照组有21个 蛋白质发生差异化表达,其中15个蛋白质在免疫应激组上调表达,6个蛋白质在免疫应激组下调表达,如表5所示。
免疫应激组上调表达的蛋白有3类。第一类是参与碳水化合物代谢和能量产生相关的蛋白,包括:AMP脱氨酶1 X1亚型(AMPD1)、甘油-3-磷酸脱氢酶1样蛋白(GPD1L2)、L-乳酸脱氢酶A链X1型(LDHA)、线粒体X2型细胞色素b-c1复合体亚基2(UQCRC2);第二类是肌肉收缩相关的蛋白,包括:肌球蛋白重链1C(MYH1C)、肌球蛋白重链1A(MYH1A)、钙调素-2(CASQ2)、三聚体细胞内阳离子通道A(TMEM38A);第三类是参与免疫反应保护细胞免受损伤相关蛋白,包括:高迁移率蛋白B1X1亚型(HMGB1)、蛋白磷酸酶1调控亚基1A(PPP1R1A)、E3 sumo蛋白连接酶ranbp2(LOC101747971)、过氧化物酶1(PRDX1)和ATP结合盒亚家族F成员2 X1亚型(ABCF2)。
免疫应激组下调表达的蛋白质有:血红蛋白β亚基(HBBA)、线粒体柠檬酸合成酶X3型(CS)、ATP合酶亚基α(ATP5A1WL)、热激蛋白90(HSP90B1)、ATP结合盒B家族成员6(ABCB6)和X1型突触小泡磷酸酶2(SYNJ2)。
2.3.2 免疫应激组肉鸡胸肌差异表达蛋白质网络互作分析 利用string在线分析软件对免疫应激组差异表达蛋白质进行网络互作功能分析,结果如图4所示,有15个差异蛋白富集到网络中。在网络互作图中,通过聚类分析发现,具有显著互作关系的功能模块主要为能量代谢(energy metabolism)和肌肉收缩(muscle contraction)子网络。每个子网络有多个节点蛋白质互作,每个节点均代表关键差异蛋白,具体信息见表5。其中,涉及能量代谢的关键节点蛋白质主要有CS、LDHA、ATP5A1WL、UQCRC2。涉及肌肉收缩的关键节点蛋白质主要有MYH1A、MYH1C、CASQ2、TMEM38A。
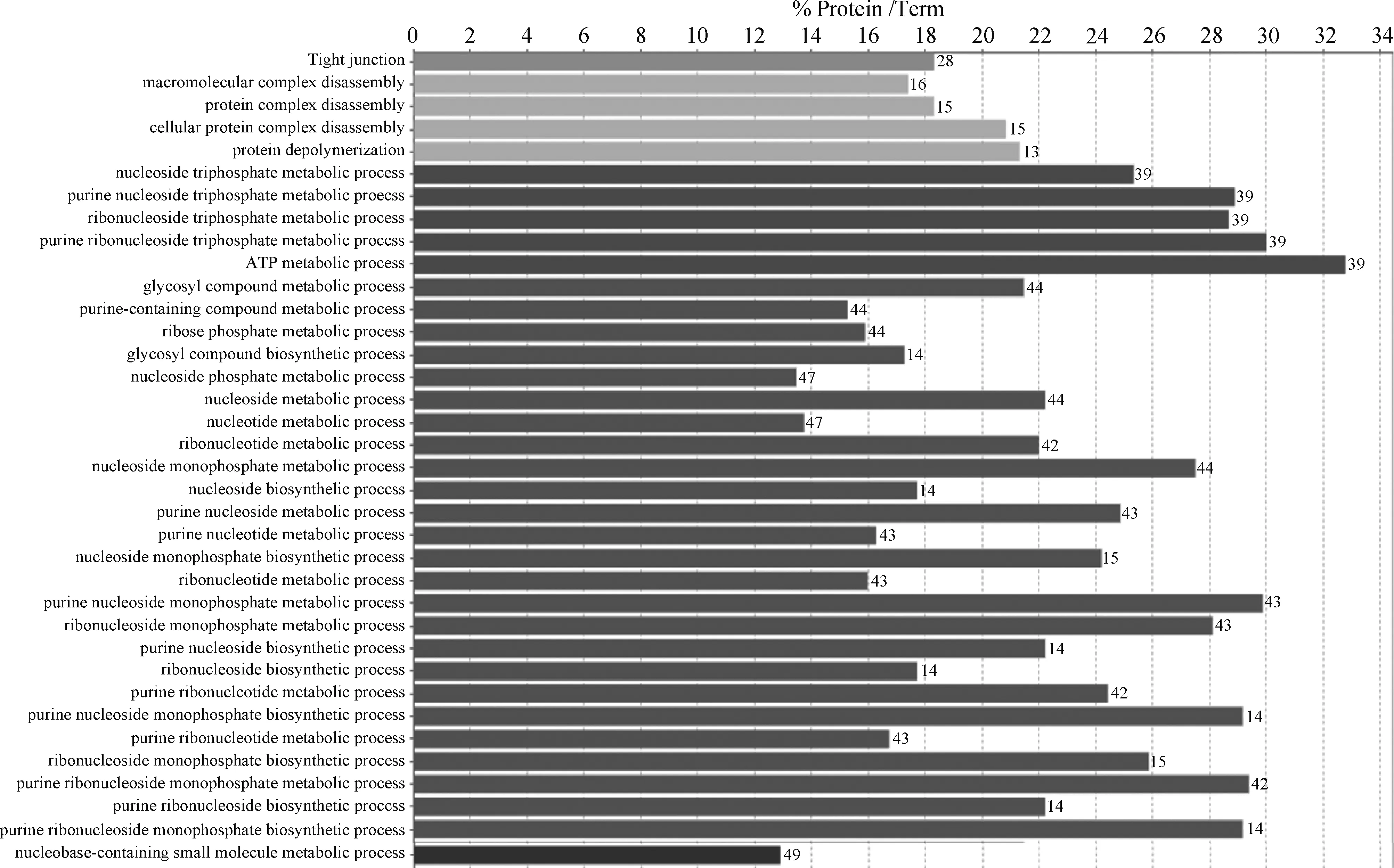
大写字母开头为KEGG注释,小写字母开头为GO注释Terms that begin with uppercase or lowercase letters are KEGG or GO annotation, respectively图3 免疫应激组肉鸡胸肌特异表达蛋白质GO和KEGG功能注释Fig.3 GO and KEGG annotation of unique proteins specially expressed in the PM of broilers in the immunological stress group
3 讨 论
国内外报道均表明,蛋白质组成的改变会引起肌肉品质的变化[23-25]。本试验定性蛋白质组结果显示,免疫应激组和对照组特异表达蛋白质分别富集到了不同功能模块,这表明免疫应激导致肌肉中的蛋白质表达种类发生了变化。同时,定量蛋白质组学结果显示,有21个蛋白质表达量发生了变化。因此,免疫应激导致肉品质下降的分子基础是组成肌肉中与肉品质相关蛋白质的质与量的改变。
研究表明,肌肉纤维的直径、类型等因素对肌肉的嫩度有重要的影响,I型肌纤维直径较细,密度较大,嫩度较高,而IIb型肌纤维则相反[26]。肌肉的总重量从出生到屠宰大约增长50~100倍,这个差异主要由肌纤维的“肥大”造成的。研究发现,在肌肉肥大的过程中,收缩蛋白的合成速率大大高于其分解速率,从而导致肌纤维中粗、细肌丝数目增加。另外,提供能量的合成酶也显著增加。本试验结果显示,免疫应激组肉鸡胸肌肌纤维横截面直径和面积均显著增加,显著降低了肉的嫩度。其分子机制可能是肌肉收缩相关蛋白的高表达(包括MYH1C、MYH1A、CASQ2、TMEM38A),同时与之相对应的为其提供能量的蛋白质的高表达共同作用的结果,这其中与能量产生相关的蛋白质有AMPD1、GPD1L2、LDHA、UQCRC2。
pH是决定肉品质好坏的重要因素[27],宰后糖酵解代谢速率直接影响肌肉pH的下降程度。研究表明,LPS诱导的肉鸡免疫应激显著降低了宰后24 h 胸肌的pH(P<0.05)[28]。本试验结果显示,免疫应激组肌肉pH没有显著变化,但从数值上看,免疫应激组pH下降程度大。宰后肌肉肌糖原和ATP被快速消耗,导致细胞由有氧呼吸向无氧呼吸转变,肌肉内乳酸累积导致pH下降[29]。Zheng等[30]研究证实,葡萄糖磷酸变位酶1、线粒体肌酸激酶、果糖-1,6-二磷酸酶、L-乳酸脱氢酶A链、甘油醛-3-磷酸脱氢酶、磷酸甘油酸变位酶1,磷酸甘油酸激酶的上调表达与宰后肌肉pH下降相关。本试验结果也显示,免疫应激导致L-乳酸脱氢酶A链表达量升高,可能会改变肌糖原的消耗速率,从而导致肌肉pH下降。

表5 免疫应激组AA肉鸡胸肌差异表达蛋白质信息
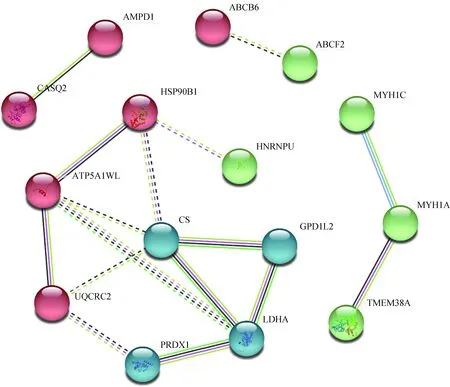
每个球体代表节点蛋白,相同颜色的球体代表不同节点蛋白聚在同一个子网络。图中实线表示两个蛋白间的互作得分大于0.5(虚线表示小于0.5)。不同颜色的实线代表蛋白间不同互作关系的证据:红线表示融合证据,绿线表示邻域证据,蓝线表示共现证据,紫线表示试验证据,黄线表示数据挖掘证据,浅蓝线表示数据库证据,黑线表示共表达证据Each ball represents node protein, the same color balls represent node proteins are clustered in the same sub network. The solid line indicates that the interaction score between the two proteins is more than 0.5 (the dotted line indicates that the score is less than 0.5). Different color solid lines between proteins represent evidence of association: Red lines indicate fusion evidence, green lines indicate neighborhood evidence, blue lines indicate co-occurrence evidence, purple lines indicate experimental evidence, yellow lines indicate text mining evidence, light blue lines indicate database evidence, and black lines indicate co-expression evidence图4 免疫应激组肉鸡胸肌差异表达蛋白质及其相互作用网络Fig.4 The interaction network of differentially expressed proteins in the PM of broiler chickens in the immunological stress group
持水力是肉品质的一个重要指标,通过滴水损失和蒸煮损失来反映。本试验结果显示,免疫应激组肌肉滴水损失和蒸煮损失均显著增加。这可能与免疫应激导致的CS、ATP5A1WL、HSP90B1下调表达有关。线粒体柠檬酸合成酶X3型,催化来自糖酵解或其它异化反应的乙酰CoA与草酰乙酸缩合合成柠檬酸反应的酶。趋于生成柠檬酸的方向,控制三羧酸循环的入口,成为三羧酸循环的限速步骤。ATP合酶是线粒体氧化磷酸化的关键酶。这两个酶的下调表达可能会导致细胞能量代谢障碍。另外,热休克蛋白表达增高,使动物组织和细胞可以更有效的抵抗各种应激反应。在蛋白质变性之初,热休克蛋白与肌原纤维蛋白结合,能够起到保护的作用,阻止各种酶类分解肌原纤维。热休克蛋白能够有效延缓细胞膜的破裂,从而阻止内源性蛋白酶的释放,使得肌肉的水分损失减少[24]。本试验中,免疫应激导致热激蛋白90的下调表达,使蛋白质无法维持其细胞的完整性和某些蛋白(如肌间线蛋白,肌浆和肌原纤维)的修复功能,从而引起细胞的持水力下降。Kristensen和Purslow[31]认为,宰后骨架蛋白如肌动蛋白等不降解,在肌肉发生僵直时会导致肌原纤维网格结构收缩形成汁液流失通道而增加汁液流失。在本试验中,免疫应激导致细胞骨架蛋白肌球蛋白重链(MYH1C和MYH1A)高表达,可能与其降低持水力有关。因此,免疫应激可能引起了细胞结构和功能的损伤,导致肌肉细胞的持水力下降,增加了滴水损失和蒸煮损失。
氨基酸含量和组成与肉的鲜味高度相关。其中,与鲜味有关的最主要氨基酸是谷氨酸,肌苷酸与谷氨酸钠协同影响肉的鲜味[32]。国内外研究表明,鸡肉质鲜味特性的主要物质基础是由肌苷酸(IMP)决定的[33-35]。本试验结果显示,免疫应激组肉鸡肌肉中IMP含量极显著降低,这可能是由于免疫应激导致免疫相关蛋白高表达,而与风味物质相关的蛋白低表达所致。如HMGB1、PPP1R1A 和PRDX1等参与免疫反应,保护细胞免受损伤相关蛋白发生了高表达。高迁移率蛋白B1X1亚型(HMGB1)是多功能蛋白,促进宿主对感染性信号的炎症反应,并参与先天和适应性免疫反应的协调和整合。在组织损伤时,能够放大免疫反应信号,促进LPS绑定因子的释放,并激活细胞表面受体参与免疫反应。蛋白磷酸酶1调控亚基1A(PPP1R1A)具有抗原蛋白的绑定分子功能,促进微管相关蛋白TAU/MAPT的去磷酸化;过氧化物酶1(PRDX1)通过解毒过氧化物和作为过氧化氢介导的信号传感器,在抗氧化应激中发挥保护细胞的作用。ATP结合盒亚家族F成员2 X1亚型在巨噬细胞的脂质代谢和神经发育中起作用。
4 结 论
本试验对比研究了对照组和免疫应激组肉鸡胸肌肉品质和蛋白质组的差异,发现免疫应激组肉鸡胸肌肉品质下降的分子机理:1)免疫应激可能通过提高肌肉收缩相关蛋白和与之相对应的为其提供能量的蛋白质的高表达,显著增加肉鸡胸肌肌纤维横截面直径和面积,显著降低肉的嫩度;2)免疫应激导致L-乳酸脱氢酶表达量升高,可能会改变肌糖元的消耗速率,从而导致肌肉pH下降。3)免疫应激通过下调表达CS、ATP5A1WL、HSP90B1,引起细胞结构和功能的损伤,导致肌肉细胞的持水力下降,增加滴水损失和蒸煮损失。4)免疫应激可能通过提高免疫反应相关蛋白的表达,降低与风味相关物质的合成,降低肉质风味。
猜你喜欢
肝博士(2022年3期)2022-06-30
今日农业(2021年7期)2021-11-27
海外星云(2021年9期)2021-10-14
今日农业(2020年18期)2020-12-14
中成药(2017年8期)2017-11-22
中国现代医学杂志(2015年26期)2015-12-23
少儿科学周刊·少年版(2015年11期)2015-12-17
少儿科学周刊·儿童版(2015年11期)2015-12-17
医学研究杂志(2015年7期)2015-06-22
西安交通大学学报(医学版)(2015年2期)2015-02-28
