
实验性免疫性不育雄性小鼠滤泡辅助性T细胞相关细胞因子水平分析①
2021-08-23 03:06曹晓丹魏任雄张晓霞浙江中医药大学附属宁波中医院男科实验室宁波315010
中国免疫学杂志 2021年12期
曹晓丹 魏任雄 张晓霞 周 俊 (浙江中医药大学附属宁波中医院男科实验室,宁波315010)
不育症日益受到社会的广泛关注,我国约有15%~20%的育龄夫妇不能生育,男性因素约占50%[1-4]。免疫性不育是在某些外因或内因的诱发下,自身免疫耐受状态被打破,持续迁延的自身免疫对自身抗原产生异常的免疫应答,导致免疫调节失衡,进而导致疾病发生。滤泡辅助性T(T follicular helper,Tfh)细胞由 IL-21 等诱导 Th0 分化产生,是真正意义上辅助B 细胞的一群新的CD4+Th 细胞亚群[5]。Tfh 细胞的产生和功能表达受多种因素调控。Bcl-6 等被认为是Tfh 细胞产生和分化的关键分子,IL-4、IL-21等细胞因子是Tfh细胞和B细胞状态和功能的关键指标。本研究通过建立免疫性不育小鼠模型,分析免疫性不育小鼠与健康对照小鼠脾脏淋巴细胞中滤泡辅助性T细胞相关细胞因子变化,探讨滤泡辅助性T细胞在免疫性不育发病过程中的意义。
1 材料与方法
1.1 材料
1.1.1 实验动物 SPF 级BALB/c小鼠,雄性,35只,6~8 周龄,体重16~18 g,购自上海斯莱克实验动物有限责任公司。对BALB/c 小鼠昼夜12 h 交替光照,自由食水,于室温(23±1)℃条件下饲养。
1.1.2 仪器与试剂 IL-4、IL-12、IL-17、IL-21 ELISA试剂盒(美国BioLegend公司);Trizol试剂、反转录试剂盒、RT-PCR 试剂盒(日本TaKaRa 公司);APC 标记抗小鼠CD4 单克隆抗体、PerCP/Cy5.5 标记抗小鼠CXCR5 单克隆抗体、FITC 标记抗小鼠ICOS 单克隆抗体(美国eBioscience 公司);小鼠抗精子抗体检测试剂盒(上海恒远生物科技有限公司);iMark 酶标仪(美国Bio-Rad 公司);ABI 7500 PCR 仪(美国ABI 公司);FACSCanto Ⅱ流式细胞计数仪(美国BD公司)。qPCR 特异引物由Primer5 软件设计(表1),上海桑尼生物科技有限公司合成。

表1 Bcl-6、IL-4、IL-12、IL-17、IL-21基因引物序列Tab.1 Primer sequences of Bcl-6,IL-4,IL-12,IL-17,IL-21
1.2 方法
1.2.1 免疫性不育小鼠模型的建立 每次免疫前取雄性BALB/c小鼠5只,共3次,CO2安乐死后取双侧睾丸组织,用生理盐水冲洗,置于M199 获能液中,剪碎,37℃ 静置15 min,尼龙网(孔径200µm)过滤,37℃ 培育 1.5 h,调整精子数为 2.0×106个/ml,加入完全弗氏佐剂,研磨混匀作为抗原。
取雄性BALB/c 小鼠20 只,随机分为正常组和实验组(10 只/组)。实验组双侧腹股沟颈背部脊柱两侧皮下多点注射0.5 ml 抗原作为首次免疫,10 d后于上述相同部位加强免疫。正常组注射生理盐水。分别于初免前及每次加强免疫后眼眶静脉采血,分离血清,ELISA 检测血清抗精子抗体(anti-sperm antibody,AsAb)。第 3 次加强免疫后检测小鼠血清AsAb 水平,筛选AsAb 阳性的小鼠进行后续研究。
1.2.2 小鼠脾脏淋巴细胞的分离 小鼠经CO2安乐死后,用75%的乙醇浸泡2~3 min,减少毛发造成的污染。将小鼠移入超净工作台,在无菌条件下,以仰卧位固定于解剖盘。采用眼科镊子夹起腹股沟中线处皮肤,眼科剪做一横切口。镊子捏住切口两端皮肤,向小鼠的尾部和头部纵向撕开皮肤,暴露腹腔膜壁,乙醇消毒。切开腹腔壁膜,采用镊子夹住脾脏,剪刀剪去与其相连的组织。将分离得到的脾脏组织浸泡于干净的PBS 溶液;在培养皿中加入适量PBS溶液,放置细胞筛,将脾脏取出置于细胞筛。取干净的注射器,用其按压处末端碾压组织。膜内细胞会慢慢游离出来,经过细胞筛后,悬浮于培养皿溶液。洗涤细胞筛,收集细胞筛外PBS 至50 ml 离心管,400 g 离心 15 min,去上清;加红细胞裂解液 2 ml,重悬,37℃ 静置 2 min 后,400 g 离心15 min,加入 5 ml PBS 洗涤,400 g 离心,去上清;加入5 ml PBS重悬,除去其中不可溶的组织纤维,悬浮液取10 µl细胞计数,调整细胞密度为1×106个/ml。
1.2.3 流式细胞术检测小鼠脾淋巴细胞内Tfh 细胞的比例[6]取1×106个上述方法分离获得的脾淋巴细胞,PBS 洗涤 2 次,4℃、1 000 r/min 离心 5 min。取100µl PBS 重悬细胞,分别加入APC 标记抗小鼠CD4 单克隆抗体、PerCP/Cy5.5 标记抗小鼠CXCR5单克隆抗体、FITC 标记抗小鼠ICOS 单克隆抗体各5 µl,混匀后4℃ 避光孵育30 min。4℃、1 000 r/min离心 5 min,弃上清,PBS 洗涤后重悬细胞,PBS 稀释至300~500µl,流式细胞仪检测。
1.2.4 qRT-PCR 检测免疫性不育小鼠脾淋巴细胞中 Bcl-6、IL-4、IL-12、IL-17、IL-21 水平 按照 Trizol试剂盒说明书提取脾淋巴细胞总RNA,并进行去除基因组 DNA 及反转录反应。Bcl-6、IL-4、IL-12、IL-17、IL-21 和内参基因GAPDH 的核酸序列通过基因库(Gene Bank)获得。反应体系采用SYBR Green 荧光染料法,避光操作,每个反应重复3 孔,在ABI 7500 PCR 仪上对转录因子和不同细胞因子转录水平进行检测,反应条件为:95℃ 30 s,95℃ 5 s,60℃34 s,共进行个循环。
1.2.5 ELISA 检测小鼠血清中 IL-4、IL-12、IL-17、IL-21水平 小鼠眼眶静脉采血,离心收集血清。待测样品孔中每孔分别加入待测血清100µl,所有样品均作复孔,反应板于37℃下孵育120 min。用300 µl 1×Wash Buffer 洗板 4~6 次,每个测试孔中分别加入IL-4、IL-12、IL-17、IL-21一抗工作液各50 µl,将反应板充分混匀后于37℃孵育60 min。洗板后,各测试孔加酶标抗体工作液100 µl,反应板于37℃孵育60 min。孵育结束后洗板,之后每孔加入底物液100 µl,置暗处反应5~10 min。每孔加入50 µl 终止液混匀。酶标仪检测450 nm 处每个孔的吸光值,根据标准品计算样品的浓度。
1.3 统计学分析 采用Graphpad Prism 5软件进行统计学分析。所有数据以表示,组间比较采用t检验,以P<0.05为差异有统计学意义。
2 结果
2.1 免疫性不育小鼠模型的建立 对第3 次加强免疫后的小鼠血清进行AsAb 检测,结果显示,实验组小鼠血清吸光度值为0.45±0.04,对照组小鼠血清吸光度值为0.18±0.01。实验组10 只小鼠AsAb吸光度/正常对照组≥2.1,即为AsAb(+)。
2.2 小鼠脾脏淋巴细胞中CD4+CXCR5+ICOS+T 细胞表达水平 将分离的脾脏淋巴细胞进行多色免疫荧光标记,以CD4+CXCR5+ICOS+作为Tfh 的标志进行分析。免疫性不育小鼠脾脏淋巴细胞中CD4+CXCR5+ICOS+T 细胞比例明显高于正常对照组(P<0.001,图1)。
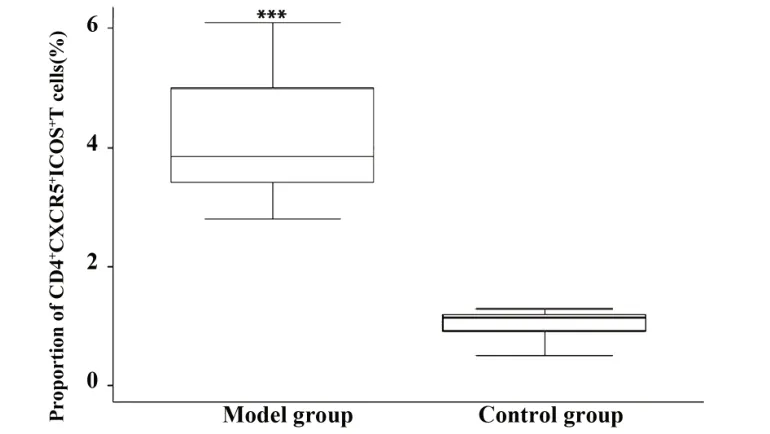
图1 小鼠脾脏淋巴细胞中CD4+CXCR5+ICOS+T 细胞的比例分析Fig.1 Analysis of proportion of CD4+CXCR5+ICOS+T cells in spleen lymphocytes of mice
2.3 小鼠脾脏淋巴细胞中 Bcl-6、IL-4、IL-12、IL-17、IL-21 mRNA 相对表达量 与对照组相比,免疫性不育小鼠脾脏淋巴细胞Bcl-6、IL-4、IL-12、IL-21 mRNA 含量升高,差异有统计学意义(P<0.05),IL-17 mRNA 含量变化无统计学差异(P>0.05)。见图2。
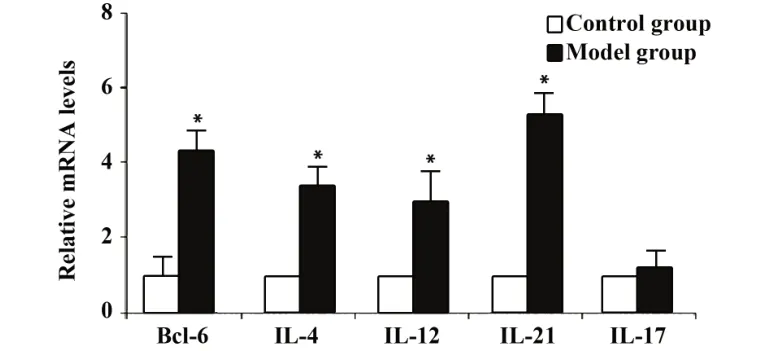
图2 小鼠脾脏淋巴细胞中Bcl-6、IL-4、IL-12、IL-17、IL-21 mRNA相对表达量Fig.2 mRNA relative expression levels of Bcl-6,IL-4,IL-12,IL-17,IL-21 in spleen lymphocytes of mice
2.4 小鼠血清中IL-4、IL-12、IL-17、IL-21 水平 与对照组相比,免疫性不育小鼠血清中IL-4、IL-12、IL-21 的含量升高(P<0.05),IL-17 含量变化差异无统计学意义(P>0.05,图3)。
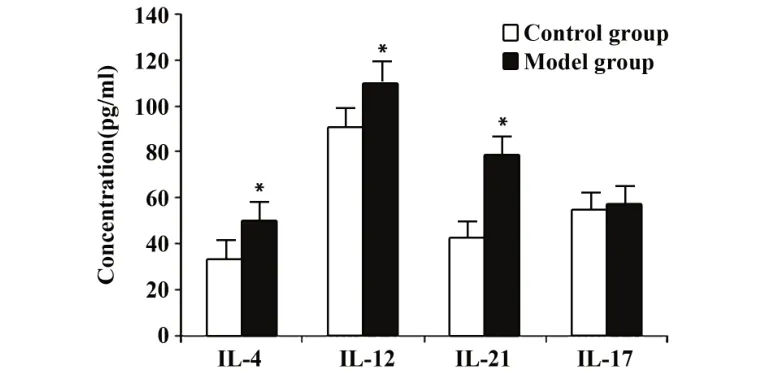
图3 小鼠血清中IL-4、IL-12、IL-17、IL-21水平Fig.3 Serum levels of IL-4,IL-12,IL-17,IL-21 in mice
3 讨论
前期研究认为Th1/Th2 细胞亚群的失衡和免疫性不育的发生有关[7]。在免疫系统中,初始CD4+T 淋巴细胞(Th0)在局部微环境中受不同细胞因子调控向不同方向分化,其分化方向决定免疫应答的类型。IL-21 和 IFN-γ 等可诱导 Th0 向 Th1 分化,主要介导细胞免疫应答。IL-4 等可诱导Th0 向Th2 分化,主要介导体液免疫应答。TGF-β 和IL-2可诱导Th0 向Treg 分化,主要发挥免疫抑制和免疫调节作用。TGF-β 和 IL-6(小鼠)或 IL-1β 和 IL-6(人)等可诱导Th0 向Th17 分化,主要通过分泌IL-17 刺激多种细胞参与机体的免疫防御,在固有免疫中发挥重要作用。由于Th2细胞分泌的IL-4等细胞因子能够诱导B 细胞活化、增殖及抗体产生,因此Th2 细胞一直被认为是辅助B 细胞的主要辅助性T细胞,但实验发现IL-4 基因敲除小鼠并不影响抗体的产生[8]。Tfh 细胞由 IL-21 等诱导 Th0 分化产生,是真正意义上辅助B 细胞的一群新的CD4+Th 细胞亚群。Tfh 细胞辅助B 细胞在生发中心存活、增殖,促进B细胞向浆细胞分化、抗体类别转换,在自身免疫性疾病中扮演重要角色[9]。
Tfh 细胞的产生和功能表达受多种因素调控。转录因子B 细胞淋巴瘤6(B cell lymphoma 6,Bcl-6)是Tfh 细胞特异性的转录因子,是区别于Th1、Th2、Th17 等辅助性T细胞的特征之一,在Tfh 的分化、特异性表面分子的表达及T细胞依赖的生发中心免疫应答中起关键作用[10]。研究显示,Bcl-6 缺陷的小鼠体内不能形成生发中心,Tfh 细胞发育障碍,且不能对胸腺依赖的抗原产生免疫应答[11]。本研究采用RT-PCR 方法检测免疫性不育小鼠脾脏淋巴细胞中Bcl-6 的mRNA 水平,结果显示,在免疫性不育小鼠中,Bcl-6 的表达显著高于健康对照小鼠。流式检测结果显示,免疫性不育小鼠脾脏淋巴细胞中CD4+CXCR5+ICOS+Tfh 细胞比例明显高于正常对照组。实验结果提示免疫性不育小鼠脾脏淋巴细胞中Bcl-6 的升高可诱导初始T 细胞向Tfh 分化,从而增加免疫性不育小鼠中Tfh 的数量。
免疫微环境中的相关细胞因子,如IL-12、IL-21等可促进初始 CD4+T 细胞分化为 Tfh 细胞[12-13]。IL-21 是Tfh 细胞的重要功能性细胞因子,具有强大的辅助诱导功能,IL-21 可通过与B 细胞表面受体结合促进B 细胞高频突变、Ig 类别转化及浆细胞形成[14-15]。IL-21受体(IL-21 receptor,IL-21R)表达在B细胞、T 细胞和NK 细胞表面,其中B 细胞是其作用的最主要的靶细胞。通过自分泌的方式,IL-21 能促进Tfh 自身的分化,同时影响Tfh 细胞转录因子Bcl-6的表达[16]。高表达IL-21 的小鼠浆细胞数目显著上升,而在IL-21 缺失或IL-21R 缺陷的小鼠中,T细胞依赖型抗体的分泌发生障碍,提示IL-21 为B细胞分化及类别转换的重要细胞因子,在B 细胞对T 细胞依赖抗原的免疫应答及体液免疫的长期维持中发挥重要作用[17]。在本研究中,免疫性不育小鼠脾脏淋巴细胞中IL-4、IL-12、IL-21 mRNA 的表达显著升高,两组间IL-17 的表达差异无统计学意义。ELISA 法检测血清中 IL-4、IL-12、IL-21、IL-17 的水平,得到了同样的结果。推测在免疫性不育的发生过程中,大量炎症细胞及免疫细胞被活化,产生大量 IL-4、IL-12 及 IL-21,导致免疫微环境紊乱,使CD4+Th 细胞向Tfh 细胞分化、增殖。本研究阐明了Tfh 细胞相关细胞因子在免疫性不育小鼠发病中的表达特征及意义,为后续Tfh 细胞在辅助免疫性不育机体B细胞分泌自身抗体过程中具体作用及机制的研究奠定基础。
猜你喜欢
昆明医科大学学报(2022年4期)2022-05-23
中国民间疗法(2021年13期)2021-08-30
天津医科大学学报(2021年2期)2021-03-29
现代临床医学(2019年6期)2019-12-07
临床医药文献杂志(电子版)(2017年11期)2017-05-17
当代医药论丛(2017年22期)2017-04-12
中国卫生标准管理(2015年15期)2016-01-15
现代检验医学杂志(2015年6期)2015-02-06
当代畜禽养殖业(2014年11期)2014-02-27
西南军医(2014年1期)2014-02-03
