
不同载畜率对短花针茅荒漠草原优势物种间关系及其空间分布特征的影响
2021-08-21 02:32张峰孙嘉伟孙宇郑佳华乔荠瑢赵萌莉
草业学报 2021年8期
张峰,孙嘉伟,孙宇,郑佳华,乔荠瑢,赵萌莉*
(1.内蒙古农业大学草原与资源环境学院,内蒙古 呼和浩特010019;2.内蒙古农业大学草地资源教育部重点实验室,内蒙古呼和浩特010019)
种间关系是决定草地群落组成结构、动态变化的重要因素[1],是形成草地群落空间格局的主要动力[2]。目前,对草地群落物种间关系的研究较多[3-4],但在反映物种在群落中的相对重要性及物种间亲和或竞争的相对关系时,却无法直观反映种群在空间上的动态变化规律[5]。借助地统计学对不同草地类型的不同指标测度进行分析[6-8],能反映植物种群空间分布格局的动态变化,并能模拟还原物种分布及其种间关系。近年来,由于过度放牧造成各草地类型出现不同程度的退化,使得群落盖度、多样性及生产力降低[9];而荒漠草原是我国北方天然的生态屏障,是以放牧为主的畜牧业基地[10],其气候干燥、生境脆弱且对干扰响应敏感[11]。但目前对荒漠草原放牧干扰的相关研究主要集中于群落结构、物种多样性、生产力、性状属性及土壤理化性质等方面[12-13],对多物种间空间分布关系的研究较少;而张峰等[14]对大针茅(Stipa grandis)典型草原空间分布关系的研究发现,放牧会导致种间关系改变;而Callaway等[15]和Armas等[16]的研究表明,植物的相互作用影响着种群动态。
以短花针茅(Stipa breviflora)荒漠草原为对象,结合常规分析方法与地统计学方法,分析建群种短花针茅与优势种无芒隐子草(Cleistogenes songorica)和冷蒿(Artimisia frigida)种间关系及空间分布特征,阐明优势物种的种间关系及其作用强度,并结合空间模拟揭示物种间的动态变化,为草地合理利用提供理论基础。本研究旨在回答以下3个问题:1)优势种无芒隐子草和冷蒿与建群种短花针茅之间是竞争关系还是亲和关系?不同载畜率是否会改变物种之间的关系?2)物种之间的作用强度是怎样的?不同载畜率是否会改变物种间的作用强度?3)不同载畜率下优势物种间的空间分布特征是怎样的?对其进行空间模拟是否具有实际意义?
1 材料与方法
1.1 研究区概况
试验区位于内蒙古农牧业科学院综合试验示范中心四子王旗基地(41°46′44″N,111°53′42″E),海拔1456 m;典型的中温带大陆性季风气候,春季干旱多风,夏季炎热,而冬季寒冷漫长。>10℃的年积温为2200~2500℃,气温平均日差为13~14℃;平均无霜期约为108 d。在2004-2018年期间,年均温3.4℃,最冷月在1月,均温约为-16.03℃,最热月在7月,均温约为20.82℃,极值为-20.38和23.73℃;年均降水量为234 mm,降水主要集中在4-9月;年均风速为4~5 m·s-1。地带性植被类型为短花针茅荒漠草原,建群种为短花针茅,主要优势种为无芒隐子草和冷蒿,伴生种为银灰旋花(Convolvulus ammanni)、阿尔泰狗娃花(Heteropappus altaicus)、小叶锦鸡儿(Caragana microphylla)、木地肤(Kochia prostrata),其他常见植物有糙苏(Phlomis umbrosa)、驼绒藜(Ceratoideslatens)等,还有一、二年生植物地锦(Euphorbia humifusa)、刺穗藜(Chenopodum aristatum)等;植被稀疏低矮,群落高度为8~10 cm,盖度为17%~20%。土壤为淡栗钙土,土层厚度约为1 cm,较贫瘠,有机质含量较低,约为14 g·kg-1。
1.2 试验设计
试验区于2004年进行围建,设置对照(control,CK)、轻度放牧(light grazing,LG)、中度放牧(moderate grazing,MG)和重度放牧(heavy grazing,HG)4个载畜率水平,放牧家畜为当地成年蒙古羯绵羊,分别为0、4、8、12只,载畜率分别为0(CK)、0.15(LG)、0.30(MG)和0.45(HG)羊单位·hm-2·月-1;3次重复,共12个小区,各小区面积均为4.4 hm2;在围建前,草地放养绵羊且草地利用强度较大,但围建时各小区草地及土壤状况较一致。自2004年起,放牧期于6月初开始并于11月底结束。
由于12个小区均呈现南北长、东西窄的特点,本试验于2020年7-9月在每个小区分别设置1个25 m×100 m的大样方,均等划分为50个5 m×10 m的中样方(南北长10 m,东西宽5 m)(图1),在每个中样方东南角设置1个1 m×1 m的小样方;各小区小样方均为50个,共计600个。以西南角作为起点(0,0),自西向东为X轴,自南向北为Y轴,最西南角坐标为(0,0),最东北角坐标为(20,90);记录各小样方中短花针茅、无芒隐子草及冷蒿的株丛数(多度)。
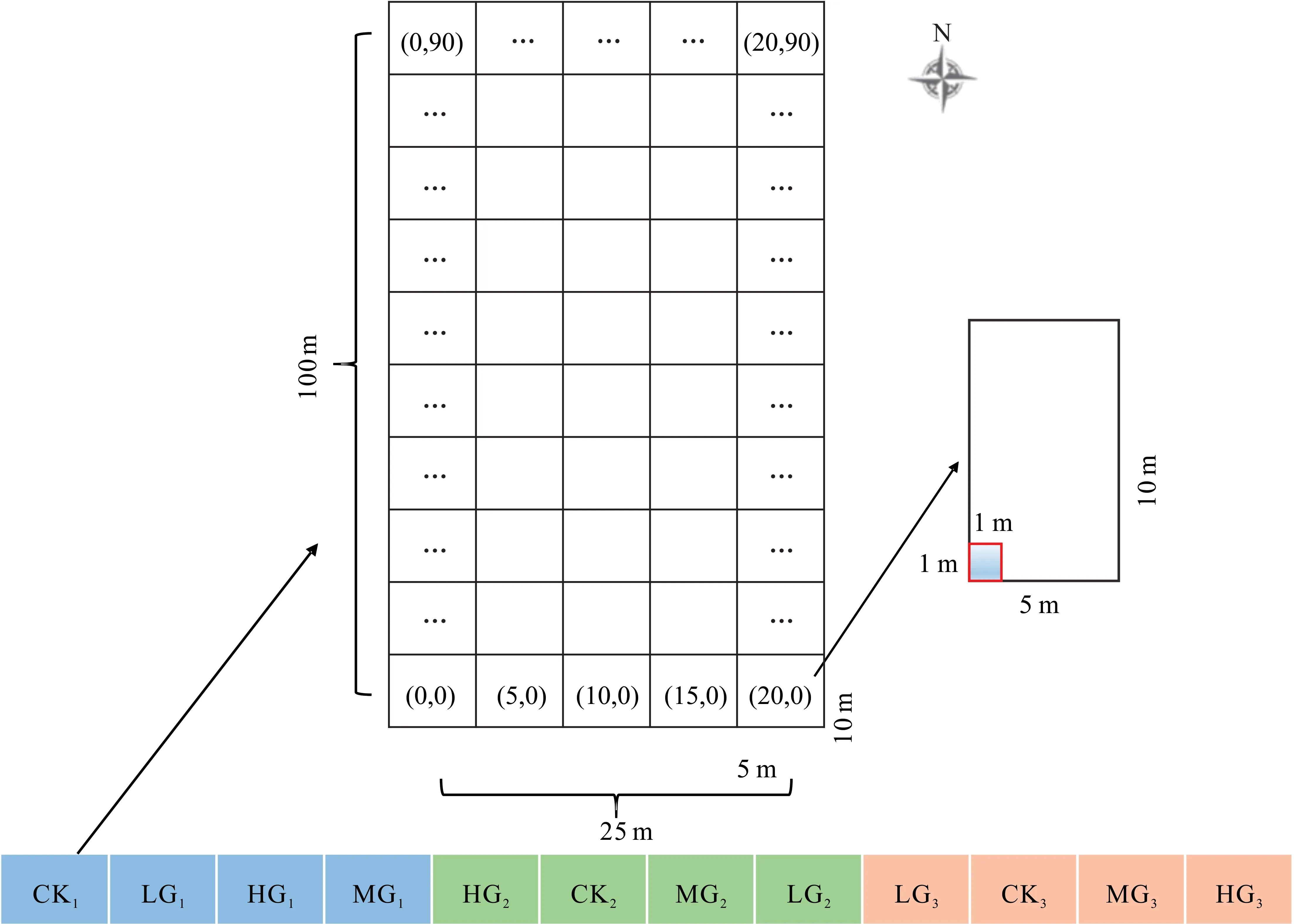
图1 样地及取样示意图Fig.1 Schematic diagram of plot and sampling
1.3 数据处理
首先,对建群种短花针茅与优势种无芒隐子草和冷蒿种群出现频率及多度进行处理及月份下双因素方差分析;其次,以建群种短花针茅为自变量,以优势种无芒隐子草和冷蒿为因变量分别进行回归分析;再者,对优势种两两进行物种对同分布指数分析,得出6个物种对的物种作用强度;最后,将优势种无芒隐子草(X)和冷蒿(Y)种群多度定义为有序的坐标变量,分析建群种短花针茅(Z)受优势种无芒隐子草和冷蒿种群的影响特点和程度。物种多度数据均经过正态转化,符合正态分布后使用地统计学软件GS+9.0进行半方差函数分析,得到最优模型及优势种群间空间分布关系的分形维数后通过克里格(Kriging)插值法绘制其3D立体图,通过立体图峰谷之间的过渡及凹凸情况来判断优势种群间分布关系情况。所有数据分析基于SPSS 19.0软件,所有图表制作基于SigmaPlot 14.0。显著性水平为α=0.05。此外,频率(frequency,f)及同分布指数公式(co-occurrence index,CO)[17]如下:

式中:N i代表物种出现的样方数;N为总样方数;O1,2代表两个物种同时出现的样方数;O1代表只有物种1出现的样方数。如果CO1,2值很大,说明物种2在物种1出现的大多数样方都出现,物种1受到物种2影响的可能性比较大;如果CO1,2值很小,说明在物种1分布的大多数样方内都没有物种2个体,二者发生作用的可能性很小。
2 结果与分析
2.1 优势种群密度
对建群种短花针茅及优势种无芒隐子草和冷蒿种群进行分析表明,不同放牧处理、月份及其交互作用均对短花针茅和无芒隐子草出现频率无显著影响(P>0.05);而冷蒿出现频率仅在处理间差异显著(P<0.05)。然而,不同放牧处理、月份及其交互作用对短花针茅、无芒隐子草及冷蒿种群多度均存在显著影响(P<0.05)(表1)。

表1 优势种群频度及多度的双因素方差分析Table 1 Two-way analysis of variance for frequency and abundance of dominant population
2.2 优势种群出现频率
不同放牧处理及月份下,短花针茅及无芒隐子草种群出现频率分别为91.67%~100.00%和85.33%~100.00%(P>0.05)。不同放牧处理下冷蒿种群出现频率存在显著差异(P<0.05);与CK相比,LG仅在7月时显著降低了50.54%(P<0.05),而MG与HG在7-9月均显著降低(P<0.05),MG在7-9月分别降低了76.92%、77.78%和78.57%,HG则分别降低了100.00%、92.59%和100.00%;但各放牧处理在不同月份间无显著差异(P>0.05)(图2)。
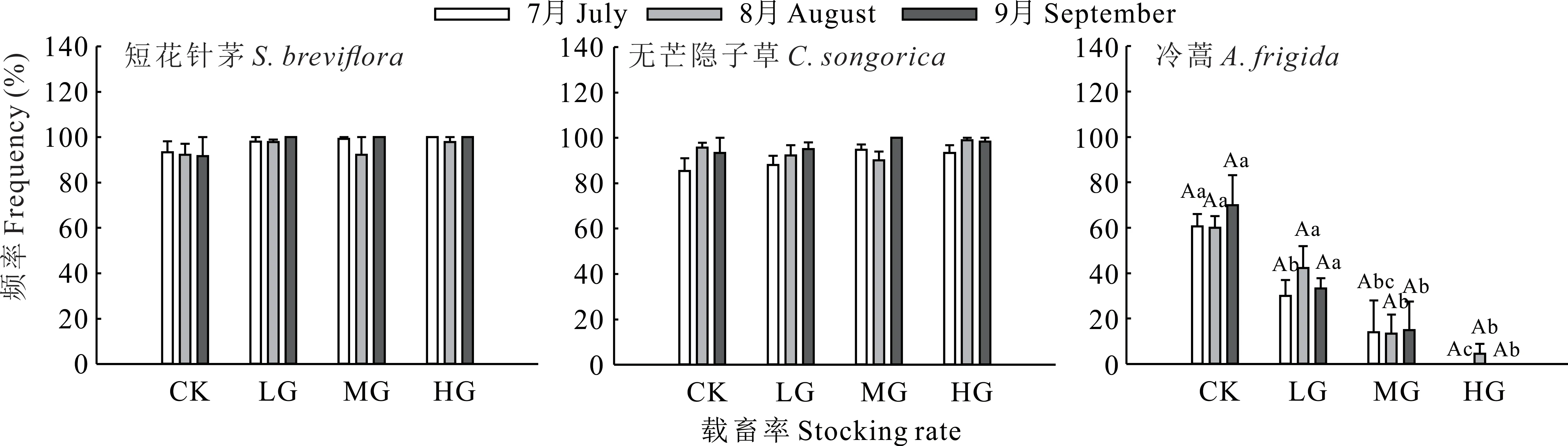
图2 不同载畜率下优势物种出现频率Fig.2 Frequency of dominant species under different stocking rates
2.3 种群多度
对短花针茅而言,与CK相比,种群多度在7-9月均显著增加(P<0.05),LG、MG和HG分别在7-9月平均增加约37.34%、75.72%和99.32%;但与7月相比,8和9月短花针茅种群多度均显著降低(P<0.05)。对无芒隐子草而言,与CK相比,LG在7和8月时显著增加(P<0.05),MG和HG在7-9月时均显著增加(P<0.05);与7月相比,LG和MG在8月时种群多度显著增加(P<0.05)。对于冷蒿而言,与CK相比,各处理种群多度在7-9月均显著降低(P<0.05),LG、MG和HG分别在7-9月平均降低约61.26%、66.32%和99.31%;与7月相比,CK、MG和HG在8月时种群多度均显著增加(P<0.05)(图3)。

图3 不同载畜率下优势物种多度Fig.3 Abundance of dominant species under different stocking rates
2.4 建群种与优势种间的回归分析
7月时,短花针茅与无芒隐子草间在CK中存在显著正相关关系(P<0.05);8月时,在MG和HG中存在显著正相关关系(P<0.05);9月时,在CK中存在显著正相关关系(P<0.05),在HG中存在显著负相关关系(P<0.05)。然而,7月时,短花针茅与冷蒿间在CK和MG中存在显著负相关关系(P<0.05);8月时,在CK、LG和MG中存在显著负相关关系(P<0.05);9月时,在CK和LG中存在显著负相关关系(P<0.05)(表2)。
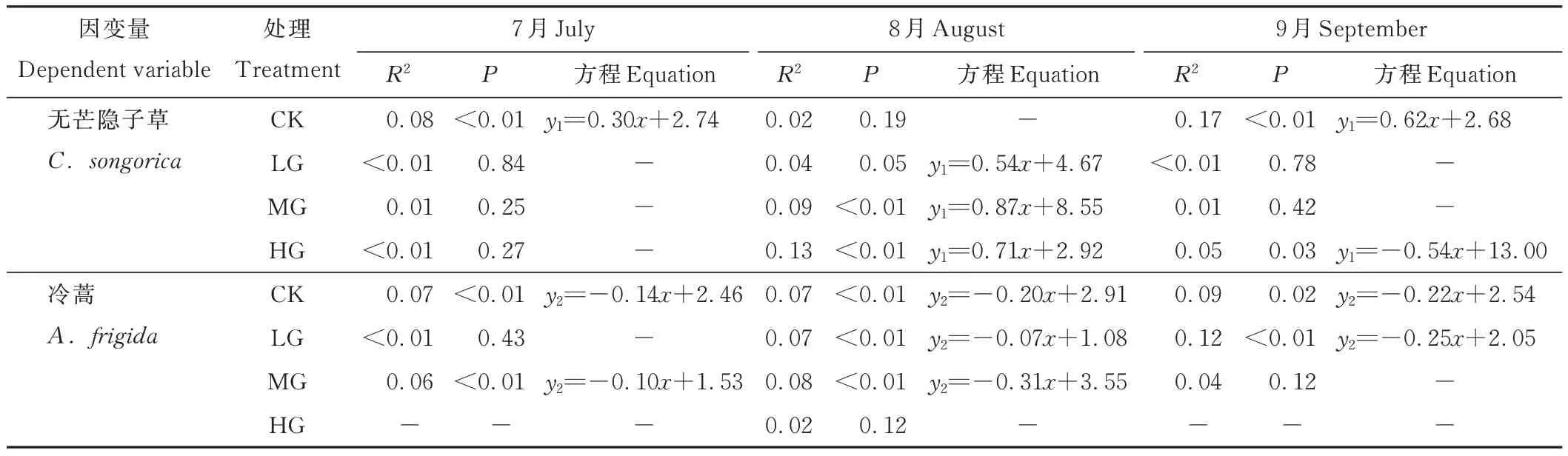
表2 建群种与优势种间的回归分析Table 2 Regression analysis between the constructive species and the dominant species
2.5 物种间的同分布指数
对短花针茅、无芒隐子草和冷蒿进行两两物种对同分布指数分析表明,短花针茅与无芒隐子草对冷蒿的作用强度较大,而冷蒿对短花针茅和无芒隐子草的作用强度较小;此外,短花针茅、无芒隐子草和冷蒿之间的相互作用主要集中在7和8月。与CK相比,7和8月时,LG下短花针茅对无芒隐子草和冷蒿作用强度增大。与CK相比,8月时,LG和MG下无芒隐子草对短花针茅和冷蒿作用强度降低,HG下无芒隐子草对短花针茅作用强度增大。与CK相比,冷蒿对短花针茅和无芒隐子草的作用强度变化较小(表3)。

表3 物种间的同分布指数Table 3 Co-occurrence index among species
2.6 建群种与优势种间的空间分布
9月时,HG下无芒隐子草与短花针茅种群多度存在负相关关系(表2),而其他月份或处理中,短花针茅种群多度均与无芒隐子草种群多度为正相关关系,与冷蒿种群多度为负相关关系,故主要对9月的空间分布进行分析。结果表明,CK中,短花针茅种群多度随无芒隐子草种群多度增加而呈增加的趋势,随冷蒿种群多度增加而呈降低的趋势。LG中,当冷蒿种群多度较低时,随着无芒隐子草种群多度的增加,短花针茅呈先增加后降低,而后增加再降低的趋势;当冷蒿种群多度较高时,随着无芒隐子草种群多度增加,短花针茅种群多度呈先降低后增加的趋势。MG中,当冷蒿种群多度较低时,随着无芒隐子草种群多度增加,短花针茅种群多度呈先增加后降低的趋势;当冷蒿种群多度较高时,随着无芒隐子草种群多度增加,短花针茅种群多度呈逐渐降低的趋势。HG中,由于冷蒿出现较少、密度较低,导致其种群分布较少,故其种群密度在图中形成窄条,其多度不随短花针茅与无芒隐子草种群多度的变化而变化,随着无芒隐子草种群多度增加,短花针茅种群多度呈先增加后降低,而再增加后再降低的趋势,整体上呈波动下降的趋势(图4)。
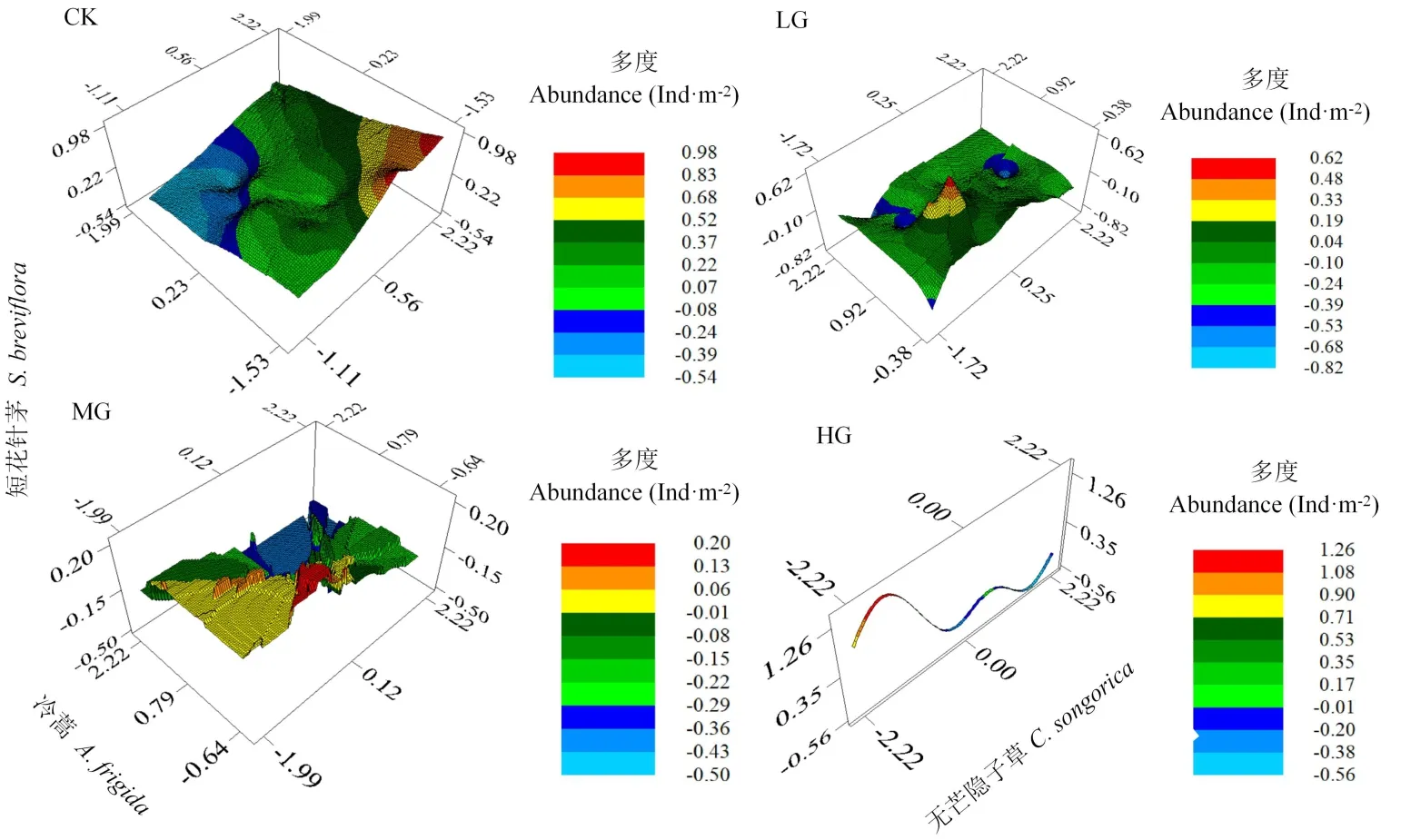
图4 不同载畜率下建群种与优势种间空间分布特征Fig.4 Spatial distr ibution characteristics of constr uctive species and dominant species under different stocking r ates
3 讨论
在长期放牧过程中,群落结构及种群分布会发生改变[18]。本研究中,放牧导致短花针茅和无芒隐子草种群多度增加,冷蒿种群多度降低。一方面由于放牧会移除一部分植株而降低顶端优势,因而促进丛生禾草分蘖,造成株丛增大或植株密度增加[19];另一方面由于在长期放牧过程中,随放牧强度增大,短花针茅大株丛逐渐形成若干个小株丛,脱离母株后进行营养繁殖使种群密度增大[20-21]。古琛等[22]在对荒漠草原建群种短花针茅生长和繁殖策略的研究也表明,随放牧强度的增大,短花针茅种群受家畜践踏趋于破碎化;张峰等[14]在对大针茅典型草原建群种大针茅和优势种羊草(Leymus chinensis)及糙隐子草(Cleistogenes squarrosa)的研究中发现,其三者在放牧下种群密度显著增加。本研究中,短花针茅和无芒隐子草均为多年生丛生禾草,受家畜采食和践踏作用,使得株丛破碎、株幅变小,使其种群密度增大;一方面由于冷蒿适口性高于短花针茅和无芒隐子草,家畜喜食冷蒿造成其光合面积减少及再生能力下降,进而造成其种群多度及出现频率降低;另一方面,冷蒿生殖方式与短花针茅与无芒隐子草存在差异,短花针茅与无芒隐子草受放牧干扰时出现矮型化、破碎化,而冷蒿主要以匍匐地面的方式降低株高以躲避家畜采食。此外,放牧家畜的过度践踏使土壤紧实度和容重增大[23],间接阻碍了根系的伸长与扩展[24],也会对冷蒿生长与繁殖造成一定的影响。
希吉日塔娜等[25]对不同放牧制度下短花针茅荒漠草原种间关系的分析表明,种间关系在两物种间是不对等的,种间亲和关系会因放牧制度的改变而改变;本研究也发现,短花针茅与无芒隐子草、冷蒿种群多度之间的相关性存在动态变化;此外,短花针茅与无芒隐子草对冷蒿的作用强度较大,而冷蒿对短花针茅和无芒隐子草的作用强度较小。此外,本研究发现,短花针茅主要与无芒隐子草表现为亲和关系,与冷蒿表现为竞争关系;但9月时短花针茅与无芒隐子草间关系从对照下的亲和关系转化为重度放牧下的竞争关系。吴艳玲等[26]对不同放牧强度下物种间关系研究表明,重度放牧造成物种对正相关对数减少,种间竞争与亲和作用可以相互转化。方楷等[27]对不同放牧制度下荒漠草原种间关系研究也表明,轻度放牧不会改变植物的种间关系,但重度放牧加剧了资源竞争,还会引起群落逆向演替。但张爽[28]在短花针茅荒漠草原种间关系对放牧响应的研究中表明,放牧使得主要物种间关系向正关联方向发展;此外,不同放牧处理下,物种间亲和或竞争关系均存在不对称性;放牧造成种内竞争强度增加,但种间竞争强度减弱。刘菊红等[29]对重度放牧下荒漠草原主要植物种间关系的研究表明,重牧使群落物种间关系由竞争转变为亲和。由于种间关系存在多度效应,因干扰条件或物种不同,植物种间关系存在多种表现形式[30]。
由于各物种在放牧干扰下存在不同的生存繁殖策略,而其繁殖策略是植物在其生活史中对外界干扰的响应,在不同生境中的繁殖对策反映了其对环境的适应能力和生殖潜力[31]。短花针茅和无芒隐子草均为多年生丛生禾草,在长期放牧过程中,短花针茅和无芒隐子草株丛倾向于破碎化,种群高度倾向于矮型化[20-22];随放牧强度增大,冷蒿通过茎的斜倚程度甚至匍匐于地面使得种群矮化来躲避家畜采食[32]。此外,草地群落中大多植物同时采取有性和无性繁殖方式来实现种群生长和繁殖,植物在扰动大、竞争较弱的环境中倾向于有性繁殖,而在相对稳定的环境中倾向于无性繁殖[33-34]。随着载畜率增大,冷蒿有性繁殖分配减少、生殖枝数和种子产量降低,重度放牧下有性生殖可能消失[35];而短花针茅也倾向于分蘖减少、种子减少和降低幼苗存活率。故在本研究中,重度放牧下短花针茅与无芒隐子草关系从亲和变为竞争,表明重度放牧导致种间关系发生变化。
4 结论
放牧降低了冷蒿的出现频率,但未改变短花针茅和无芒隐子草的出现频率;放牧使得短花针茅和无芒隐子草种群多度增加,造成冷蒿种群多度降低。其次,短花针茅主要与无芒隐子草表现为亲和关系,与冷蒿表现为竞争关系;但9月时短花针茅与无芒隐子草间关系从对照下的亲和关系转化为重度放牧下的竞争关系,短花针茅与冷蒿间关系无变化。此外,通过对9月物种间空间分布进行模拟也发现短花针茅与无芒隐子草之间关系由对照中的亲和关系转变为重度放牧下的竞争关系,表明利用地统计学对其物种间空间分布分析较合理。
猜你喜欢
今日农业(2022年15期)2022-09-20
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
晚晴(2020年1期)2020-06-19
老年教育(老年大学)(2020年6期)2020-01-02
大学教育(2019年9期)2019-09-20
红土地(2018年7期)2018-09-26
蔬菜(2018年5期)2018-05-17
杂草学报(2015年2期)2016-01-04
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
当代畜禽养殖业(2014年10期)2014-02-27
