
新疆草地优势种植物相对生物量沿海拔梯度变化特征
2021-08-21 02:32:04马婧婧刘耘华盛建东李宁武红旗贾宏涛孙宗玖程军回
草业学报 2021年8期
马婧婧,刘耘华,盛建东,李宁,武红旗,贾宏涛,孙宗玖,程军回*
(1.新疆农业大学草业与环境科学学院,新疆 乌鲁木齐830052;2.新疆土壤与植物生态过程重点实验室,新疆 乌鲁木齐830052)
草地作为世界上面积最大的陆地生态系统,其地上生物量在防风固沙、涵养水源、畜牧业发展和碳循环中均扮演着重要作用[1-2]。现有研究表明,草地生态系统中植物的地上生物量既受到海拔和降水等环境因素的影响[3-4],也受到群落结构的影响[5]。质量比假说(mass-ratio hypothesis)认为,优势种由于在群落中拥有较多的个体且在资源竞争中占据优势地位,是草地生态系统中生物量的主要贡献者[6]。我国内蒙古草地和北美草地的研究也证实了优势种地上生物量可占据总生物量的46.3%~87.6%和70%~80%[7-8]。鉴于优势种在群落地上生物量中的重要作用,研究优势种地上生物量沿环境梯度的变化规律,对于深入理解草地植物地上生物量变化特征和草地管理,均有重要意义。
山地生态系统中,海拔被认为是影响地上生物量变化的主要因素[9]。现有研究表明,草地生态系统中优势种地上生物量随海拔的变化,因研究对象的不同和海拔梯度差异,呈多种变化特征。例如:对我国四川岷江干旱河谷中的优势种地上生物量研究发现,莸(Caryopteris)灌丛的地上生物量随海拔上升呈降低的趋势,而小花滇紫草(Onosma farreri)和川甘亚菊(Ajania potaninii)灌丛的地上生物量则随海拔梯度的升高呈增加的变化趋势[10]。此外,对九寨沟马脑壳金矿山暖温带高山草甸草地的研究则显示,一些主要优势种植物,如糙野青茅(Deyeuxila scabrescens)、白顶早熟禾(Poa acroleuca)和四川嵩草(Kobresia setchw anensis),其地上生物量随海拔升高均呈先升高后降低的单峰型变化趋势[11]。而对新疆塔尔巴哈台山优势种鸭茅(Dactylis glomerata)的研究则发现,其地上生物量在各海拔地段无显著差异[12]。上述这些研究极大地促进了人们对优势种地上生物量随海拔梯度变化规律的理解,但也存在一些亟待解决的问题。首先,这些研究侧重于分析优势种地上生物量随海拔变化特征,而忽略了其相对生物量的变化特征。考虑到优势种地上生物量因物种和所处的环境梯度不同存在显著差异[8],用地上生物量研究时存在标准不统一的问题,而相对生物量则反映了优势种地上生物量占群落总生物量的百分比,具有统一的比较标准。因此,从相对生物量的角度来分析优势种地上生物量随海拔梯度的变化特征,可扩大和丰富人们对优势种生物量与海拔之间关系的理解。其次,考虑到群落中同时存在着多个优势种[7],人们对于这些共存优势种之间,其相对生物量之间关系的理解,仍所知甚少。
新疆作为我国主要的草地分布区之一,由于“三山夹两盆”的特殊地形地貌,使得新疆草地表现为明显的垂直地带性分布特征[13]。新疆草地拥有较大的海拔梯度,其海拔落差从最低处的-155 m(吐鲁番艾丁湖)到最高处的8611 m(喀喇昆仑山的乔戈里峰),海拔落差为8766 m[13]。这种较大的海拔梯度,不仅形成了丰富的草地类型,也孕育了大量的特有植物,如千叶蓍(Achillea millefolium)和伊犁绢蒿(Seriphidium transiliense)等[13]。这些特征为研究草地中优势种相对生物量与海拔梯度的关系,提供了理想的研究对象。基于此,本研究以新疆草地为研究对象,通过对新疆9种草地类型中397个样地优势种地上生物量的调查,重点解决以下3个科学问题:1)不同草地类型中优势种组成有何异同?2)优势种植物相对生物量沿海拔梯度,呈何种变化特征?3)共存优势种之间,其相对生物量呈何种内在联系?
1 材料与方法
1.1 研究区概况
本研究区位于我国新疆维吾尔自治区,地理位置为73°40′-96°18′E,34°25′-58°10′N。研究区总面积为1.66×106km2,约占中国陆地面积的1/6。新疆由于地处我国西北,远离海洋,属典型的温带大陆性气候,光热资源充足,日照时数达2550~3500 h,无霜期长达180~220 d,降水少,气候干燥。海拔变化范围为-155(吐鲁番艾丁湖)~8611 m(喀喇昆仑山的乔戈里峰),年均降水量变化范围为10~550 mm,年平均温度在9~12oC。草地是新疆的主要植被类型之一,总面积达5725.88×104hm2,草地面积占新疆总面积的34%[13]。据统计,新疆共有11种草地类型,分别为低平地草甸、温性荒漠、温性草原化荒漠、温性荒漠草原、温性草原、温性草甸草原、山地草甸、高寒荒漠、高寒草原、高寒草甸和沼泽。其中,山地草地为新疆所特有的草地类型。该区域主要的土壤类型为荒漠土、棕钙土、栗钙土、黑钙土、高山(亚高山)草原土、高山(亚高山)草甸土、盐碱土、风沙土等[13]。
1.2 样地布设和地上生物量调查
本研究以中国1∶100万的植被图为基础,结合草地类型及其空间分布位置,于2011-2013年7月中旬-8月下旬,植物生长达到最大生物量时进行调查[14]。在全疆共调查了588个样地,各样地具体调查方法如下:首先,在每个样地,随机设置一个100 m×100 m的调查区域,记录其草地类型、经纬度和海拔后(GPS,UniStrong)。在调查区域的对角线上设置一条100 m的样线,沿此样线,随机设置3~5个1 m×1 m的样方。其后,对每个样方根据《中国高等植物图鉴》[15]和《新疆高等植物检索表》[16]进行物种鉴定和记录,其后,按物种分别收获其地上生物量,并经野外阴干和实验室烘干(75℃,48 h)后,测定其干重。
1.3 相对生物量计算和优势种筛选
在完成上述调查的基础上,本研究仅对397个按照1 m×1 m标准面积调查的样地,根据文献中介绍的方法,首先对每个样方,计算各物种地上生物量占群落总生物量的百分比作为相对生物量[6]。其次,将各样地的物种按相对生物量排序后,将相对生物量>5%的物种确定为优势种[6]。筛选后统计发现,这397个样地共涉及9个草地类型(图1)。其中,低平地草甸(low plain meadow,LM)、高寒草甸(alpine meadow,AM)、高寒草原(alpine steppe,AS)、山地草甸(mountain meadow,MM)、温性草甸草原(temperate meadow steppe,TMS)、温性草原化荒漠(temperate steppe desert,TSD)、温性草原(temperate steppe,TS)、温性荒漠草原(temperate desert steppe,TDS)和温性荒漠(temperate desert,TD)各有样地38、2、20、6、10、36、67、85和133个(图1)。
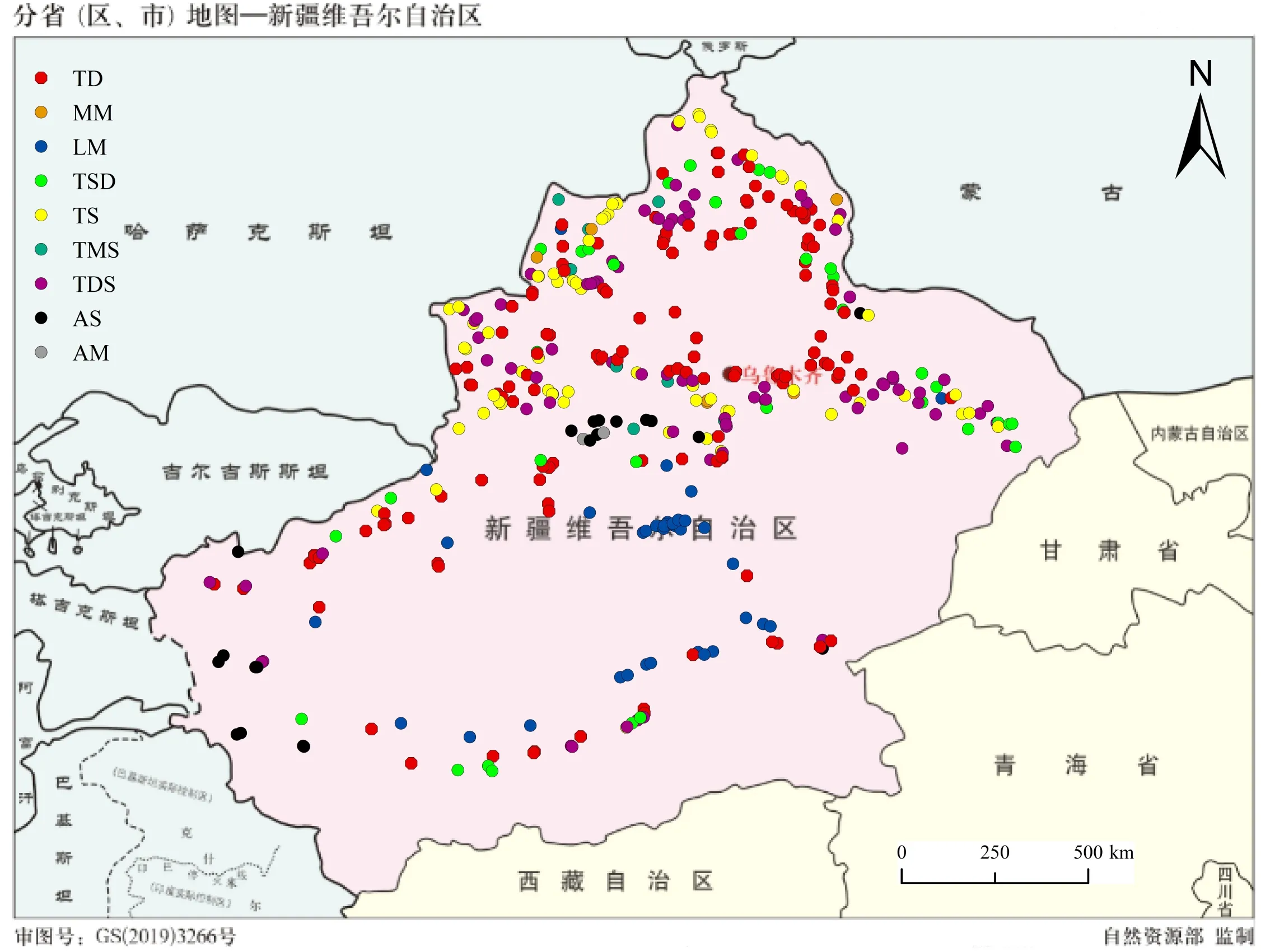
图1 研究区域和样地分布Fig.1 Study area and sites distr ibution
1.4 数据统计与分析
首先,在统计各草地类型优势种基础上,按样方统计各优势种出现的频次,来阐述各草地类型的主要优势种组成。其次,根据统计学要求,对样本量大于5的优势种(出现样方数大于5),采用一般线性模型对二者关系分别进行了线性和非线性的拟合[17]。如果某物种相对生物量与海拔之间同时存在显著的线性和非线性关系,则进一步通过赤池信息标准(akaike information criterion,AIC)来选取最优的拟合模型(AIC值越低,模型拟合效果越好)[18]。如果某物种相对生物量与海拔之间,仅存在一种显著关系(如线性或非线性关系),则直接采用该关系作为最优拟合模型[19]。如果某物种相对生物量与海拔之间线性和非线性关系均不显著,则表明二者之间无显著关系[19]。最后,对群落中共存的优势种且符合统计要求(出现样方数大于5)的物种,采用Pearson相关系数,两两量化二者之间关系。本研究所有分析均在R软件中显著性水平0.05的情况下完成。并根据分析结果,利用SigmaPlot 10.0进行作图。
2 结果与分析
2.1 不同草地类型中优势种组成
本研究所涉及的9种草地类型中,共有优势种植物169个,隶属31科120属。其中,优势种植物出现最多的为禾本科,共计33个物种,占总优势种植物的19.53%。其次分别为菊科、藜科和豆科,分别占总优势种植物的17.75%、14.20%和8.88%。优势种植物出现最少为白刺科、报春花科、车前科、大戟科、大麻科、夹竹桃科、龙胆科、牻牛儿苗科、毛茛科、旋花科、鸢尾科、紫草科,各自仅占总优势种的0.59%(表1)。

表1 各科优势种名录、数量和占总优势种的百分比Table 1 Names,number and percent of dominant species to total dominant species at family level
从草地类型来看,各草地类型中出现的优势种,因草地类型不同而异(表2)。其中,高寒草甸和山地草甸中优势种数量均少于5。高寒草甸优势种为珠芽蓼和紫花针茅,而山地草甸则为千叶蓍、早熟禾和苔草。其他7种草地类型中,优势种数均大于5。其中,芦苇是低平地草甸的最主要优势种,其出现的样方数为104个。在高寒草原、温性草甸草原、温性草原化荒漠、温性草原和温性荒漠草原中,最主要的优势种为针茅,其出现的样方数分别为30、13、26、91和81个。而在温性荒漠,其最主要的优势种则为角果藜,共出现在74个样方。
2.2 优势种相对生物量与海拔之间的关系
对满足统计学要求,即出现样方数大于5的77个优势种,进一步通过一般线性模型分析发现,在新疆草地中,优势种相对生物量与海拔之间关系,可以归为以下5类:1)优势种相对生物量与海拔之间无显著关系,共计有50个优势种(P>0.05),占总分析优势种的64.9%。2)优势种相对生物量随海拔升高,呈先降低后增加的U型变化趋势(图2)。该类共有9个优势种,分别为白羊草、假木贼、赖草、蛇麻黄、苔草、星毛委陵菜、珠芽蓼、猪毛菜、补血草,占总分析优势种的11.7%。3)优势种相对生物量与海拔之间呈显著正相关关系(图3)。该类优势种共有8个,分别为白茎绢蒿、旱蒿、黄芪、藜、蒲公英、沙生针茅、野韭菜、针茅,占总分析优势种的10.4%。4)优势种相对生物量与海拔之间呈显著负相关关系(图4),该类优势种共有6个,分别为博洛塔绢蒿、高山绢蒿、角果藜、芦苇、短花针茅和苦豆子,占总分析优势种的7.8%,其共同特征为普遍存在于温性荒漠中(表2)。5)优势种相对生物量与海拔之间呈先升高后降低的单峰型关系(图5),该类优势种共有4个,分别为叉毛蓬、昆仑针茅、镰芒针茅、羊茅,占总分析优势种的5.2%。
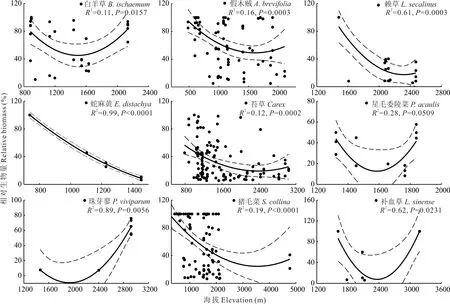
图2 9种相对生物量与海拔之间表现显著U型关系的优势种Fig.2 Nine dominant species for which their relative biomass exhibited a U-shaped r elationship with elevation
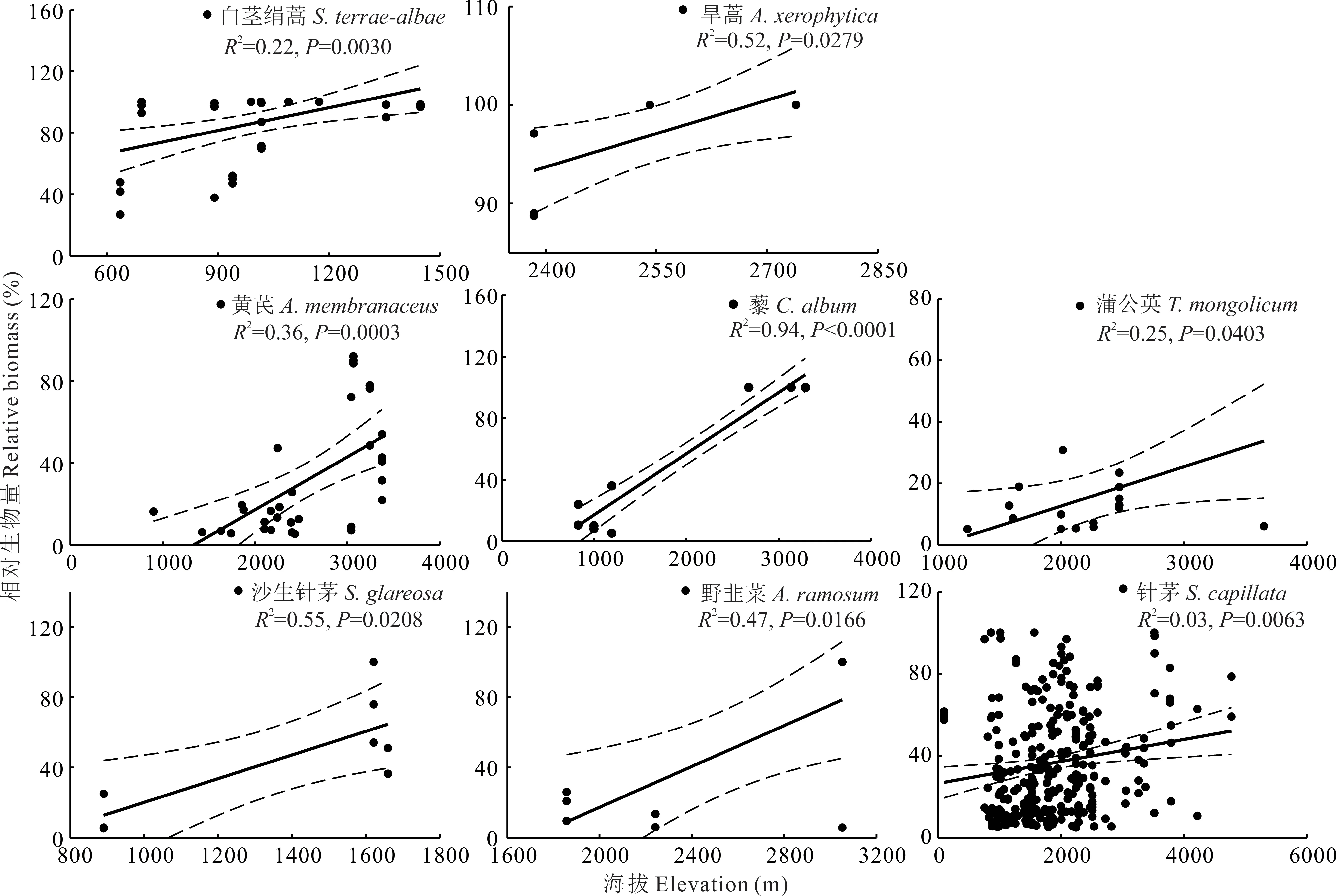
图3 8种相对生物量与海拔呈显著正相关关系的优势种Fig.3 Eight dominant species for which their relative biomass were positively correlated with elevation
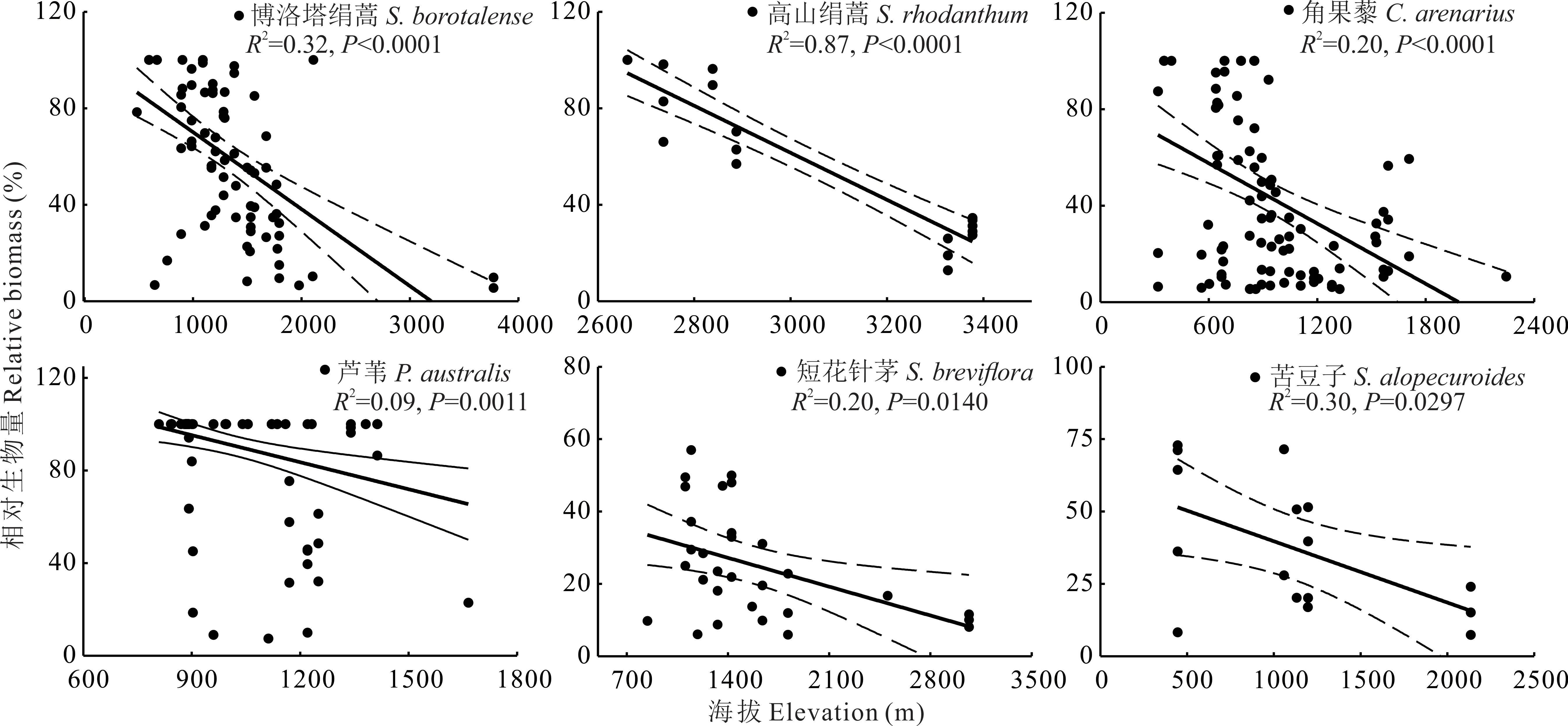
图4 6种相对生物量与海拔呈显著负相关关系的优势种Fig.4 Six dominant species for which their relative biomass were negatively associated with elevation

图5 4种相对生物量与海拔呈单峰型关系的优势种Fig.5 Four dominant species for which their relative biomass exhibited a unimodal pattern with elevation

表2 各草地类型中优势种名录及其出现的频次Table 2 Names and frequency of dominant species for each grassland type

续表1 Continued Table1
2.3 共存优势种相对生物量之间的关系
对样本量符合统计要求的24个共存优势种相对生物量分析结果表明,有19对共存优势种表现为显著相关关系(表3)。其中,仅羊茅和星毛委陵菜相对生物量之间表现为显著正相关关系,其他18对优势种的相对生物量,均表现为显著负相关关系。具体而言,博洛塔绢蒿与4种优势种植物呈显著负相关关系,分别为针茅、叉毛蓬、角果藜和苔草,角果藜和木地肤与3种优势种植物呈显著负相关关系。戈壁针茅和针茅与2种优势种植物呈显著负相关关系。此外,白羊草、羊茅、花花柴和唐松草仅与1个优势种呈显著负相关关系。

表3 共存优势种相对生物量之间相关系数Table 3 Correlation coefficients of relative biomass among coexisted dominant species
3 讨论
3.1 新疆草地不同草地类型中优势种组成
本研究基于2011-2013年对新疆草地优势种的调查发现,9种草地类型共涉及169个优势种,隶属于31科120属。通过对比20世纪80年代对新疆草地的调查发现,虽然经过20多年的变化,但新疆草地中优势种并未发现明显变化。例如,20世纪80年代调查结果显示,温性草原的主要优势种为羊茅、针茅和昆仑针茅,温性草原化荒漠中的主要优势种为纤细绢蒿、博洛塔绢蒿、沙生针茅、镰芒针茅,低平地草甸中的主要优势种为芦苇、芨芨草、甘草、花花柴等[13],本研究结果与其一致。这些结果表明,新疆各草地类型的优势种组成,并未随时间发生变化。
3.2 优势种相对生物量与海拔之间的关系
前期研究发现随海拔升高,草地生态系统中优势种生物量呈增加、降低,先增加后降低和无显著关系等多种形式[10-12]。本研究通过分析新疆草地中优势种相对生物量与海拔的关系也发现,二者关系可分为以下5类:无显著关系、先降低后增加的U型变化特征、显著正相关关系、显著负相关关系和先增加后降低的单峰型变化特征。第一类为出现比例最多的,二者之间无显著关系,这与前期在塔尔巴哈台山地优势种鸭茅的研究结果相同[12]。导致这种现象的可能原因有:第一,海拔梯度的不同所致,由于植物分布受到海拔的影响[9,20-21],这一类植物普遍的特征是其分布的海拔范围较窄。第二,由于这些植物普遍表现为一年生植物,存在明显的年际效应,其在多雨的季节和年份对群落地上生物量贡献较大[22],可能也是导致没有显著关系的原因之一。第二类关系随海拔升高呈U型变化趋势,即随着海拔的升高优势种相对生物量呈先降低后升高的变化趋势,这类变化趋势的原因目前并不清楚,有待未来进一步的研究。第三类为正相关关系,这些植物主要分布在温性草原及温性荒漠类草原中,其分布的海拔较低且水分条件较差。由于植物的生长和生物量普遍受到水分的限制,随着海拔的升高,使得土壤水分增多,有利于植被的生长,其地上生物量也呈增加的趋势[10,23]。第四类为显著的负相关关系,导致这种现象的可
能原因是,这些植物主要分布于海拔较低的荒漠类草原,普遍表现为耐旱和生长速度较低的特征,随着海拔升高和水分可利用性增加,其他生长速度快的物种,由于在水分竞争中占据优势,可通过竞争排斥降低该类植物的生物量[24]。第五类为单峰型,即随着海拔的升高优势种地上生物量呈先升高后降低的变化趋势,这与在九寨沟马脑壳金矿山暖温带高山草甸草地优势种生物量的研究结果一致[11]。这可能是因为随着海拔的升高,中海拔地区适宜大多数草本植物的增长,而高海拔地区的温度及湿度降低、空气稀薄,不适于植物的生长,则地上生物量表现为先升高后降低的变化趋势。如本研究中的羊茅,在海拔3000 m时生物量达到峰值,其后呈下降的变化特征。
3.3 共存优势种间相对生物量之间的关系
大量研究证实,群落中地上生物量主要由几个共存的优势种所贡献[7-8]。本研究表明,新疆草地中共存优势种间相对生物量以负相关关系为主,仅有1对物种在相对生物量之间,表现为正相关关系。对于正相关关系而言,其主要的原因可能是种间的互惠关系。有研究表明,在环境严酷的条件下,如高海拔和荒漠地区,个体较高的植物可通过遮挡太阳辐射,进而改变微环境,为个体较矮的植物提供一个相对适宜的环境,从而有利于其生长[25]。本研究中,由于羊茅和星毛委陵菜同时共存于气候温暖、半干旱的温性草原,羊茅作为牛羊喜食的牧草,会吸引牛羊的啃食和践踏,而放牧践踏有益于星毛委陵菜不定根的形成,促进其繁殖与生长[26]。共存优势种相对生物量之间的负相关关系,可能存在以下原因:在资源有限的生境中,一些对资源竞争占优势的植物,势必会限制其他植物的生长[27-28]。例如,博洛塔绢蒿与针茅都为生长于温性荒漠中的多年生草本植物,二者对资源的利用有同样的偏好,其对资源的竞争可能是二者存在负相关关系的主要原因[29-30]。
4 结论
通过对新疆9种草地类型中397个样地优势种系统性的调查发现,新疆草地共有优势种169种,且主要集中在禾本科。随海拔梯度,优势种相对生物量呈无显著关系、U型(先降低后增加)、显著正相关、显著负相关和单峰型(先增加后降低)5种变化形式。这些结果表明,前期研究中所报道的这些变化趋势并不矛盾,不同优势种相对生物量与海拔之间的关系,因优势种所在的草地类型和分布的海拔范围而异。此外,群落中共存优势种相对生物量之间,多表现为显著的负相关关系,表明优势种之间存在较强的竞争作用。这些结果表明,未来新疆草地在地上生物量管理和维持中,应针对各草地类型的优势种,尤其是群落中共存的优势种,予以重点关注。
猜你喜欢
科学24小时(2022年10期)2022-10-11 01:50:06
散文诗(2021年22期)2022-01-12 06:14:04
草食家畜(2021年3期)2021-06-08 03:18:16
安徽师范大学学报(2020年6期)2020-12-18 04:15:09
今日农业(2020年19期)2020-12-14 14:16:52
环球时报(2019-05-23)2019-05-23 06:17:36
少儿美术(快乐历史地理)(2018年7期)2018-11-16 05:31:12
中国科技信息(2015年2期)2015-11-16 08:18:32
河北遥感(2014年4期)2014-07-10 13:54:58
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
