
丰林自然保护区阔叶红松林采伐前和采伐中地表蜘蛛群落多样性的变化 *
2021-08-20 07:57:36王露雨李格格刘启龙高梅香
哈尔滨师范大学自然科学学报 2021年4期
陈 婷,王露雨,李格格,刘 爽,刘启龙,高梅香,
(1.哈尔滨师范大学;2.西南大学;3.宁波大学/陆海国士空间利用与治理研究中心;4.宁波市高等学校协同创新中心“宁波陆海国土空间利用与治理协同创新中心”)
0 引言
蜘蛛(Araneida)在陆地生态系统中是最丰富的捕食性天敌,在农林生态系统中的作用不可忽视[1-2].蜘蛛是受植物群落结构和空间结构这类生境因素影响较大的天敌类群,复杂的生境结构维持着蜘蛛的多样性和丰富度[3].中国农林生态系统中的蜘蛛资源非常丰富[4],不同学者从不同角度出发对农林生态系统中的蜘蛛进行了大量研究,郑国等对西双版纳地区6种林型蜘蛛的多样性进行研究,认为植物群落结构、枯落物厚度是影响一个地区蜘蛛丰富度的重要因素[5].蜘蛛的数量变化和种类组成对检测环境变化具有非常重要的指示作用[6-8],蜘蛛可以作为研究人为因素对森林生态系统造成影响的指示物种[9-11],蜘蛛群落结构易受到人为干扰的影响[12],邢树文等研究不同类型茶园游猎蜘蛛特征,发现茶园外围植被多样性与稳定性为游猎蜘蛛提供了更好的生活场所,茶园的海拔、植被状况及生境复杂程度对游猎蜘蛛优势种、丰盛种和常见种具有显著影响[2].茶园内部植被群落和生境景观的复杂程度强烈影响茶园游猎型蜘蛛物种的组成与分布[3].蒋杰贤等和冉珣隆等对不同生境条件的桃园和茶园的节肢动物进行研究发现生境的变化显著影响蜘蛛的物种数与个体数[13-14].Samu等 研究发现在小麦田中地表的裂缝对蜘蛛有吸引作用,可增加蜘蛛的个体数量[15];Tscharntke等通过对蜘蛛的研究认为草本植被的盖度和枯落物是一个地区蜘蛛丰富度最重要的解释因素[16].国内外有关蜘蛛的研究主要集中于农田、茶园等农林景观生境中,把蜘蛛作为天敌在生物防治中的作用进行了大量的研究探索[2,13-14,17-21],已有的研究多集中于农田生态系统中蜘蛛群落的组成、结构功能与人为干扰活动(农药使用等)的关系[22-23],但是对林区蜘蛛多样性分布的相关研究相对较少[5, 24],对自然保护区中林区道路建设过程中采伐活动对蜘蛛多样性分布的相关研究更少.
在林区道路的建设中,采伐是不可避免的,对森林进行大面积的采伐,大量的植被破坏,对当地森林生态环境带来一定的负面影响,对区域内及周边区域生物多样性及珍稀野生动物以及其他生境也会造成很大的影响[25-26].采伐时,往往会忽视采伐对森林地表生态环境的影响,但在采伐过程中森林地表生物多样性容易遭到破坏,一旦破坏难以恢复.同时,采伐对地表的节肢动物的种类、数量等有一定的影响[27].该试验区的采伐方式为皆伐,皆伐是对森林生物多样性影响危害最严重的一种采伐方式,也是最不科学的方式,皆伐会造成生态系统的平衡性被打破,甚至完全改变[28].卢伟等研究采伐方式对森林土壤理化性质的影响发现:皆伐对采伐区域土壤物理性质有一定的影响,但并不显著;皆伐对采伐区域土壤化学性质存在较显著的影响[29].近年来,随着对蜘蛛在农林生态系统中的重要性认知的不断提高,蜘蛛对农林生态系统的影响备受关注.因此,探讨采伐对林地蜘蛛群落的影响对于保护生物多样性和森林生态系统管理具有重要的现实意义.通过比较采伐前和采伐中的森林中地表蜘蛛组成与数量的差异,可以判断采伐对蜘蛛生物多样性的影响程度.
鉴于此,以丰林国家自然保护区为研究区,在林区道路规划建设阶段设置样地,通过调查蜘蛛种群在采伐前和采伐中的多样性变化和采伐对多样性的影响程度,揭示采伐对丰林阔叶红松林内地表蜘蛛的影响.旨在通过该研究确定该区域内采伐对蜘蛛多样性的影响程度,为减少采伐活动对蜘蛛群落多样性的影响提供一定的数据支持,为该区域对蜘蛛多样性的影响及生物多样性的保护和林区合理开发利用提供一定的依据,从而指导森林管理工作促进森林道路建设与森林生态保护的协调发展.有关丰林国家自然保护区的研究主要集中于地表甲虫[30-32],对于林区蜘蛛的研究甚少.为了揭示采伐活动对丰林国家自然保护区蜘蛛多样性和群落组成的影响,并探讨该区域蜘蛛生物多样性的基本状况,于2019年6~7月,对丰林自然保护区蜘蛛的群落组成和多样性变化进行了研究.
1 研究区概况与研究方法
1.1 研究区自然概况
该实验研究区位于黑龙江省丰林国家级自然保护区(48°02′~48°12′N,128°58′~129°15′E),位于中国小兴安岭南麓的北部,地势较平缓,海拔为280~683 m,为坡状的低山丘陵地段和宽广的谷地,土壤类型主要为山地棕色森林土.该区域的气候属于北温带大陆性湿润季风气候区,特征为夏季高温多雨,冬季寒冷干燥,年平均气温为-0.5℃,年降水量680~750 mm,年平均蒸发量为930 mm[33-35].该实验于研究区内的36林班内进行,该林班内的植被类型是1931年自然火灾形成的次生白桦林逐步演替形成的阔叶混交林,主要为白桦(Betulaplatyphylla)、臭冷杉(Abiesnephrolepis)、红皮云杉(Piceakoraiensis)、春榆(Ulmuspropinqua)、色木槭(Acermono.Maxim)、香杨(Populuskoreana)等;灌木为毛榛子(Corylusmandshurica)、忍冬(Lonicerajaponica)等;草本植物为蕨类(Pteridophyta)、苔草(Carextristachya)等[33, 36-38].
1.2 研究方法
因道路建设施工需要,砍伐丰林自然保护区36号林班内的一片原始阔叶红松林,采伐方式为皆伐.该实验将样地设置在该作业林班内的皆伐林地内.森林采伐(皆伐)的时间为2018年6月28日~7月18日,该实验于6月11日~6月23日期间(12 d)捕获采伐前的地表蜘蛛,于6月28日~7月18日(20 d)捕获采伐过程中(后称采伐中)的地表蜘蛛.由于采伐之前需要多个单位相关手续的批件,故施工单位难以提前确定具体开始采伐的日期,给这次实验确定采伐前的实验日期带来了极大的困难,导致采伐前和采伐中采样的日期不一致.鉴于该实验在正式采伐之前的17 d便开始采集样品,整个采伐过程中也一直持续跟踪调查,所以仍能够反映采伐前和采伐中地表蜘蛛群落的动态变化.采伐样地位于林区道路(路面为沙石路面,宽约5m)和汤林线铁路(宽20 m)的边缘,采伐之前,该样地森林覆盖率高,林下郁闭度达100%.该次采伐使用大型的采伐工具(电锯等)进行,共计采伐树木585棵,该次砍伐较多的树种依次为春榆(Ulmuspropinqua)(174棵)、暴马丁香(Syringareticulata(Bl.)Haravar)(73棵)、紫椴(TiliaamurensisRupr.)(67棵)、冷杉(Abiesfabri(Mast.)Craib)(42棵)和红皮云杉(Piceakoraiensis)(38棵).砍伐带位于道路南侧并沿着道路延伸,长约300 m,宽约25 m,实验在林区道路南侧阔叶混交林中设置3条与道路平行的样带,这3条样带分别距道路边缘0、5和20 m,对比分析蜘蛛群落在采伐前和采伐中的变化,从而揭示采伐对蜘蛛群落的影响.
在每条样带上以40 m为间隔分别设置5个20 m×20 m的样方在每个样方内布设陷阱采集地表蜘蛛,在每个样方内,将3个陷阱(即采样点)随机布置在样线上.布设陷阱时,先用内径约7cm的土钻挖取一个深度约15 cm的柱状土坑,将塑料杯(高14 cm、内径7 cm)置于坑内作为陷阱,杯口与地面齐平,然后将杯口周围的地面凋落物置于原处,杯中倒入约占杯子容积2/3的饱和NaCl溶液,同时在杯口上方距离地面约10 cm处覆盖一个由4根筷子支撑的透明塑料餐盒作为防护,防止雨水和其他生物等进入陷阱对样品产生干扰,陷阱放置于野外,于森林采伐前采集样品7次(2018年6月11~23日,采样期共计12 d),森林采伐进行中采集样品7次(2018年6月28日~7月18日,采样期共计20 d),后将样品保存于95%的酒精中,室内进行后期鉴定分析.
1.3 样品分析与鉴定
取回陷阱后,将样品带回室内进行挑拣,将蜘蛛挑出后保存于95%的医用酒精中.待后续对所采集的蜘蛛样品进行鉴定.所有标本的鉴定均在LeicaM205C体视显微镜下进行种类鉴定,鉴定主要依据蜘蛛的外生殖器(雄蛛触肢器和雌蛛外雌器)来进行,对于雄蛛,选取左侧触肢,用镊子沿着触肢的腿节末端或转节末端取下进行观察.对于雌蛛,观察外雌器腹面基本结构[39].
1.4 数据处理与分析
分别将采伐前的7次和采伐中的7次样品混合在一起,将获得的实验数据分为采伐前和采伐中两部分进行分析.
1.4.1 多样性指数
物种多样性是反映群落物种组成结构的重要指标,是用来判断群落或生态系统的稳定性指标[40].选择以下指数来分析蜘蛛群落多样性特征:
(1)Shannon-Wiener指数:

(2)Pielou均匀度指数:
J=H′/lnS
(3)Simpson指数:
(4)Margalef丰富度指数:
D=(S-1)/lnN
式中:S为群落所有的物种数,Pi为群落中第i物种个体数占总个体数的比例,N全部物种的总个体数,ni为该群落内第i个物种的个体数.
(5)群落相似度指数:采用Jaccard相似性系数判断群落的相似程度,公式为
q=c/(a+b-c)
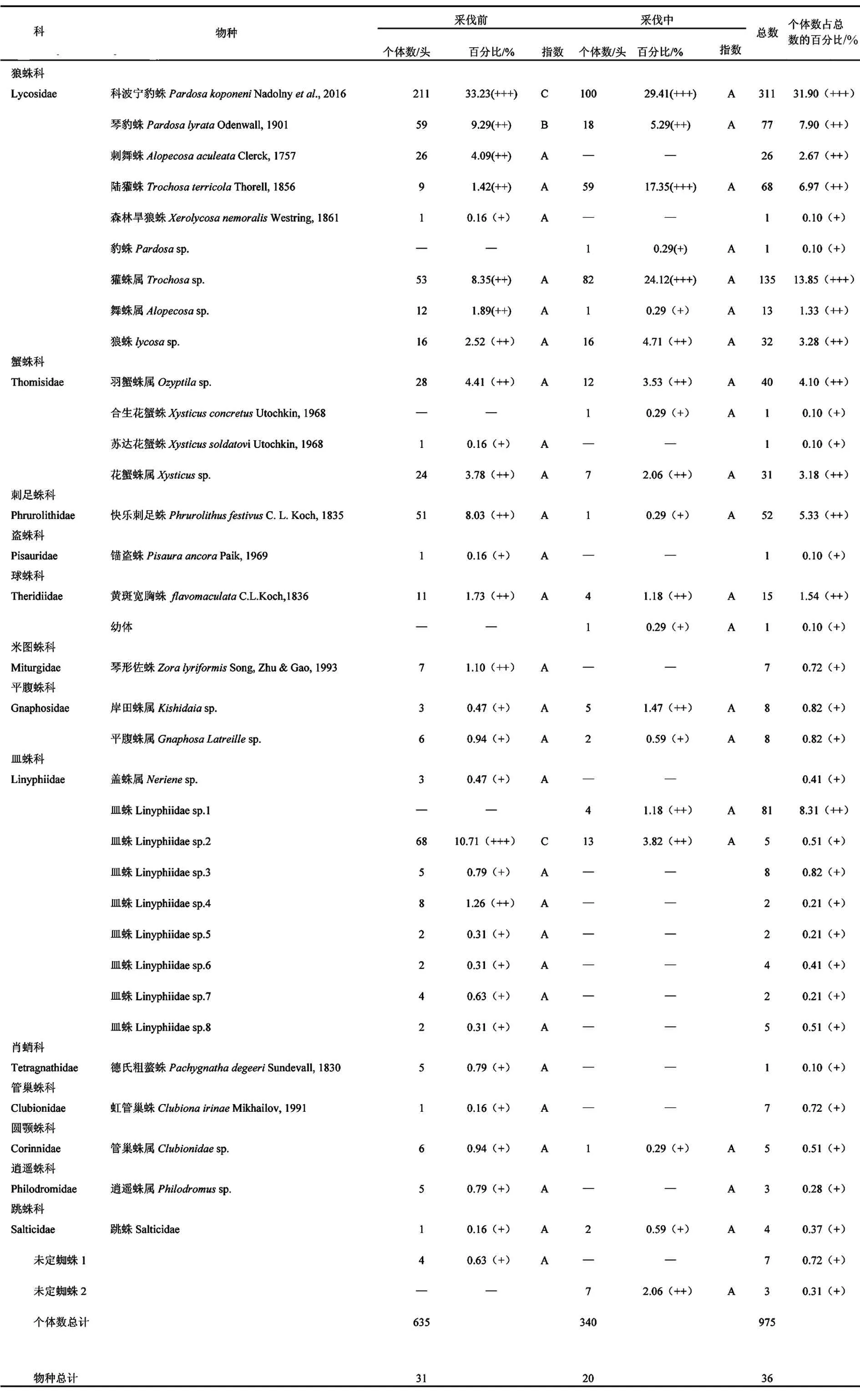
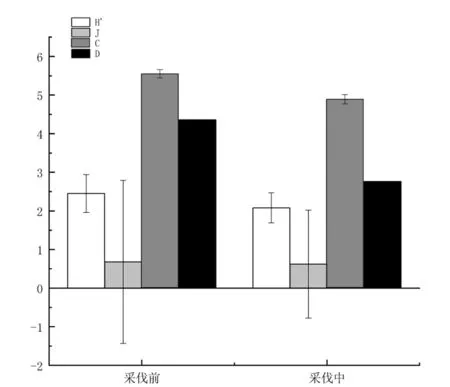
式中:c为两个群落的共有物种数,a和b分别为群落A和群落B的物种数(Jaccard)[41],若0 (6)多度等级划分 个体数占总捕获数10.0%以上者为优势物种(+++),个体数占总捕获数1.0%~10.0%者为常见物种(++),个体数占总捕获数1.0%以下者为稀有物种(+). (7)Raunkiaer频度指数 Raunkiaer频度指数为每个物种所占样点数与总样点数的比值[42],划分等级为:0~20%为A级,>20%~40%为B级,>40%~60%为C级,>60%~80%为D级,>80%~100%为E级. 1.4.2 方差分析 使用Shapiro-Wilk检验方法对数据进行正态分布检验,不符合正态分布的数据进行ln(x+1)转换使之符合或近似符合正态分布,采用单因素方差分析对比采伐前和采伐中的蜘蛛的个体数和物种数的差异性,说明采伐对蜘蛛个体数和物种数的影响.采用双因素方差分析说明采样时间和采伐活动交互作用对蜘蛛群落多样性的影响. 在软件Microsoft Excel 2016中对原始数据进行基本分析和预处理,使用R3.5.1软件中的vegan软件包中的shaprio.test函数进行正态分布检验;aov函数进行单因素方差分析和多重比较(Tukey HSD方法)来进行检验;diversity函数计算物种多样性指数. 该次实验共采集蜘蛛标本975头,分属36物种.在所有蜘蛛标本中,科波宁豹蛛(PardosakoponeniNadolny et al.)的个体数量最多,共有311头(占所有个体总数的31.90%);獾蛛属(Trochosasp.)次之,捕获135头(占蜘蛛总标本数13.85%).这2个种为优势种,合计占蜘蛛个体总数的45.75%.常见种共有10种,共有358头,占蜘蛛个体总数的44.61%.优势种和常见种的个体数占总捕获量的90.36%,是该区域地表蜘蛛的主要组成部分.其余24种为稀有物种,物种数高达总物种数的66.67%,但个体数为171头,仅占蜘蛛总捕获量的9.64%(见表1). 采伐前捕获蜘蛛635头,共31种,采伐中捕获蜘蛛340头,共20种,采伐中的蜘蛛捕获数量比采伐前减少了295头(P<0.01),采伐对蜘蛛个体数量的影响大.物种数由采伐前的31种下降至采伐中的20种,减少了11种,但差异性不显著(P>0.05)(如图1所示). 图1 采伐前和采伐中蜘蛛个体数和物种数 采伐前的优势种、常见种及稀有种的个体数量均大于采伐中的优势种、常见种及稀有种.采伐前的常见种和稀有种的物种数均大于采伐中的常见种和稀有种的物种数,采伐前的常见种有12种,稀有种有17种,采伐中的常见种有9种,稀有种有8种,采伐中的常见种比采伐前少3种,稀有种比采伐前少9种,但采伐中的优势种比采伐前的优势种多1种.采伐前多度比较为:稀有种>常见种>优势种;采伐中多度比较为:常见种>稀有种>优势种(如图2所示). 科波宁豹蛛(PardosakoponeniNadolny et al),在采伐前和采伐中均为优势物种,但个体数量减少111头;皿蛛(Linyphiidaesp.2)由采伐前的优势种变为采伐中的常见种,个体数量减少了55头.采伐中增加的优势物种为陆獾蛛(TrochosaterricolaThorell)和獾蛛属(Trochosasp.),其个体数量分别增加了50头和29头;岸田蛛属(Kishidaiasp.)和皿蛛(Linyphiidaesp.1)由采伐前的稀有物种变为采伐中的常见物种,个体数量分别增加了2头和4头;豹蛛(Pardosasp)、合生花蟹蛛(XysticusconcretusUtochkin)、皿蛛(Linyphiidaesp1)和未定蜘蛛2在采伐前未出现,在采伐中出现,但个体数量少.刺舞蛛(AlopecosaaculeateClerck,)、琴形佐蛛(ZoralyriformisSong,Zhu&Gao)和皿蛛(Linyphiidae sp.4)在采伐前为常见物种,采伐过程中未捕获;森林旱狼蛛(XerolycosanemoralisWestring)、苏达花蟹蛛(XysticussoldatoviUtochkin)和锚盗蛛(PisauraancoraPaik)采伐前为稀有物种,采伐中未捕获(见表1). 表1 采伐前和采伐中蜘蛛群落物种组成及Raunkiaer频度 就分布特征来看,优势种和常见种在采伐前和采伐中均有分布;稀有种个体数量少,多数仅分布于采伐前,在采伐中并未捕获.所有的调查中,属于Raunkiaer频度A级的物种数最多,表明绝大多数物种在样地中出现的频度较低.采伐前,优势种科波宁豹蛛(PardosakoponeniNadolny et al,)和皿蛛(Linyphiidae sp2)的频度等级均为C,琴豹蛛(PardosalyrataOdenwall)的频度等级为B,而其余物种的频度等级均为A.采伐中,所捕获的20个物种的频度等级均为A级.采伐前的Raunkiaer频度表现为A>B>C,符合Raunkiaer频度定律,即群落中低频度物种数目高于频度较高的物种数目[38](如图2所示). 从图3可以得知, Shannon-Wiener多样性指数采伐前高于采伐中,说明采伐前和采伐中蜘蛛群落的多样性存在差异,采伐活动对蜘蛛群落的多样性存在影响但不显著; Pielou均匀度指数采伐前高于采伐中,说明采伐活动对物种分布的均匀度有影响,降低了蜘蛛群落均匀度但影响不显著; Simpson优势度指数采伐前高于采伐中,说明某个物种的个体数在采伐前占总个体数的比例较高,这就是科波宁豹蛛(PardosakoponeniNadolny et al),其个体数占采伐前总个体数的33.22%;Margalef丰富度指数采伐前的高于采伐中,说明采伐前的物种丰富程度高于采伐中,采伐活动对物种的丰富度有影响. 图3 采伐前和采伐中蜘蛛群落多样性指数H′:Shannon-Wiener多样性指数 J:Pielou均匀度指数C:Simpson优势度指数 D:Margalef丰富度指数 通过计算Jaccard相似性系数,采伐前和采伐中的蜘蛛群落(q=0.41)中等不相似,群落相似程度低;对比采伐前和采伐中蜘蛛群落的优势种(q=0.25)和常见种(q=0.31),优势种中等不相似,群落优势种的相似程度和常见种的相似度都低. 采用双因素方差对采样时间和采伐对蜘蛛群落多样性指数影响对比分析,表明蜘蛛群落多样性指数在采伐前和采伐中存在极显著差异(P<0.01),均匀度指数在采伐前和采伐中存在显著差异(P<0.05),优势度指数在采伐前和采伐中的差异性不显著.多样性指数在采样时间、采样时间与采伐活动二者交互作用间均无显著差异.采伐对蜘蛛群落多样性指数和均匀度指数有显著影响. 蜘蛛一方面是重要捕食性天敌,另一方面对人为干扰以及生态环境变化敏感,已成为环境变化的指示标志[43].该研究表明,蜘蛛个体数在采伐前和采伐中存在明显差异,采伐前的蜘蛛个体数较多,有635头.蜘蛛物种数在采伐前有31种,采伐中有20种.采伐对蜘蛛的物种有影响,这与王京等研究采伐胁迫对大兴安岭针阔混交林地表节肢动物群落的长期影响得出的结论相似[44].优势种采伐前的Raunkiaer频度为A>B>C,采伐中的Raunkiaer频度均为A,说明在采伐中蜘蛛的分布不均,采伐对蜘蛛的分布产生一定影响.采伐前的常见种和稀有种无论是个体数量还是物种数均高于采伐中,采伐前的优势种虽个体数高于采伐中,但物种数却低于采伐中,说明采伐对蜘蛛群落组成有影响. 蜘蛛群落的多样性与林区自然环境和外界的人为干扰互相适应,在采伐前和采伐中产生与周围环境相适应的周期性节律动态,并且与之协同进化,该结果与马滟滟等在云南苍山火烧迹地研究不同恢复期地表植株群落多样性的发现规律基本一致[45].采伐前和采伐中的Shannon-Wiener多样性指数的变化主要与物种数和个体数的变化相关,一般地,物种数和个体数增加,多样性指数变大[46],反之则变小,该实验中多样性指数在采伐前和采伐中的变化符合这一规律.蜘蛛群落多样性指数的变化特征表明采伐活动使蜘蛛群落的多样性降低了,这与龙涛的研究结果相一致,说明采伐活动对蜘蛛群落多样性有影响[47]. 采伐对蜘蛛群落多样性指数和优势度指数的影响显著,对均匀度无显著影响,这可能与蜘蛛栖息环境和郁闭度突然下降[17],原有的生态环境遭到破坏,形成单一的环境,蜘蛛由于缺乏庇护场所,四处逃散.同时,采伐活动对蜘蛛的捕食和繁殖造成一定的影响,导致在不同样带上的蜘蛛在采伐前和采伐中的多样性发生一定变化,蜘蛛的个体数和物种数也产生变化.采伐前和采伐中蜘蛛物种数差异性不显著,但个体数和物种数的数量在采伐前和采伐中有明显的差别,可能是在采伐过程中人为的踩踏及树木倒地以及采伐工具产生的噪声等人为因素造成的蜘蛛物种数量的差异.Jaccard相似性系数表明采伐前和采伐中的蜘蛛物种相似程度低,采伐前和采伐中的优势种和常见种相似程度低,这可能与试验区捕获的蜘蛛多为游猎型蜘蛛有关[2]. 该次实验捕获的蜘蛛多为游猎型蜘蛛,游猎型蜘蛛在农林生态系统中的分布因种而异,地表的游猎型蜘蛛主要包括狼蛛科、平腹蛛科、圆颚蛛科和皿蛛科等,林中包括管巢蛛科和跳蛛科等,这次试验中这些科的蜘蛛均有捕获,其中捕获的狼蛛科蜘蛛的物种数和个体数均为捕获的所有蜘蛛中最多的,说明在实验期间,样地中狼蛛科的蜘蛛较多,这可能与地表蜘蛛地理分布的差异有关.游猎型蜘蛛的活动范围大,在地表和树干上都有游猎蜘蛛的活动,说明游猎蜘蛛在林中的分布范围较广[49],采伐前和采伐中蜘蛛个体数和物种数的变化与游猎蜘蛛的分布特点有密不可分的关系.狼蛛科的科波宁豹蛛(PardosakoponeniNadolny et al)和皿蛛科的皿蛛(Linyphiidae sp2)在采伐前为优势科,科波宁豹蛛(PardosakoponeniNadolny et al)的个体数量在采伐前捕获量和采伐中的捕获量均为总捕获个体数量中最高,采伐前捕获该物种211头,采伐中捕获该物种100头,该物种个体数量在采伐前和采伐中的捕获量虽差距较大,但仍然是所捕获蜘蛛物种中的优势种,说明该物种在该样地中的个体数量较多,符合该物种为游猎性蜘蛛的习性特点.常见种和稀有种的个体数和物种数在采伐前和采伐中的变化,及多度比较在采伐前和采伐中的变化都说明了采伐对蜘蛛群落的个体数和物种组成均有影响.游猎型蜘蛛对维持林地生态平衡起着非常重要的作用[50],在森林管理中人为干扰和管理措施的不同,比如筑路、采伐和火烧等对地表和林中的游猎型蜘蛛有很大的影响,该实验说明采伐活动对蜘蛛群落的个体数和物种数存在影响. 越来越多的研究表明,复杂多样的植被环境中蜘蛛群落多样性也增加[51-52].随着采伐的进行,森林郁闭度降低,枯落物被人为踩踏,蜘蛛的栖息环境发生变化,蜘蛛的个体数和物种多样性受到影响.采伐前与采伐中的个体数之比为0.13∶0.07,物种数量之比为0.43∶0.28.主要有两个因素,其一是采伐前与采伐中的生境存在很大的差异,采伐的森林为自然林地,自然林地具有较复杂的植被群落更厚的地表枯落物覆盖,丰富的植被形成复杂的生态环境,可以为蜘蛛提供适宜的栖息环境和遮蔽场所[1],采伐中,蜘蛛的栖息环境发生改变,生境结构不利于蜘蛛的生活与繁殖,导致采伐中的蜘蛛个体数与物种数低于采伐前的.其二是局地小气候的变化,气温和湿度、降水量等气候条件也是导致林区蜘蛛群落多样性差异的重要因素,短时的降水会对蜘蛛的物种多样性产生一定的影响.在该实验期间,有降水的天数为23 d,其中降水量在10 mm以上为11 d.采伐前适宜的温度和湿度有利于蜘蛛的生存与繁殖提高林区蜘蛛的数量和多样性,在采伐中,局地生境发生改变,湿度和温度发生变化,导致蜘蛛个体数和种类数都会减少.采伐前与采伐中的蜘蛛林地内的草本植被对蜘蛛多样性有较强的影响,合理的降低人为活动,有助于增加蜘蛛生活的生境,提高蜘蛛种群的多度,对林中生物多样性的保护有很大帮助[53]. 2018年6~7月在丰林国家自然保护区蜘蛛975头,分属36物种,个体数和物种组成在采伐前和采伐中虽有明显变化,个体数在采伐前和采伐中的差异性显著,但物种的差异性不显著.可见,采伐对林区的蜘蛛个体数和物种有一定影响.蜘蛛群落物种组成在采伐前和采伐中的相似性为中等不相似,采伐对样地蜘蛛物种组成的相似性存在影响.对比采伐和采样时间对蜘蛛群落多样性的影响表明:采伐与蜘蛛群落多样性指数和优势度指数存在显著差异.因此,采伐对林区蜘蛛群落个体数和多样性有影响,未来需要进一步探究蜘蛛对采伐等外界生物指示物作用,为监测和科学评估人类干扰活动对森林生态系统功能的影响提供依据.2 结果与分析
2.1 蜘蛛群落物种组成及数量特征

2.2 采伐前和采伐中蜘蛛群落多样性指数特征
2.3 采伐对蜘蛛群落多样性的影响
3 讨论
3.1 采伐前和采伐中蜘蛛群落组成和多样性的变化
3.2 采伐对蜘蛛群落个体数和物种组成的影响
4 结论
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
食品安全导刊(2021年20期)2021-08-30 06:40:50
小朋友·快乐手工(2018年3期)2018-04-22 11:48:52
小学阅读指南·低年级版(2017年6期)2017-06-12 08:22:47
妇女之友(2017年3期)2017-04-20 09:20:00
中国药物应用与监测(2015年5期)2015-12-11 03:15:55
小朋友·快乐手工(2015年1期)2015-03-13 00:05:24
水生生物学报(2015年1期)2015-02-28 16:01:05
河南科技(2014年18期)2014-02-27 14:14:54
中学英语之友·上(2008年11期)2008-12-08 09:29:46
