
PPARγ激动因子与胰岛素互作影响延边黄牛前脂肪细胞分化
2021-08-10 07:27:10郭盼盼张军芳李香子严昌国尹云厚
中国兽医学报 2021年6期
郭盼盼,闫 妍,张军芳,金 鑫,李香子,严昌国*,尹云厚
(1.延边大学 农学院,吉林 延吉 133000;2.贵州民族大学,贵州 贵阳 550025)
延边黄牛是我国五大地方优良品种之一,是我国畜禽品种基因库中一份极其珍贵的“财富”,经过长期的自然选择和人工选择,已形成了适应性强、屠宰率高、肉质优良等特点,具有生产高档牛肉的优秀潜力,但其优良肉质性状形成的具体调控机制仍不是很明确。目前,畜牧学研究者比较重视肌内脂肪的研究,因为动物肉品质的好坏与肌内脂肪含量的高低显著相关[1],而较低的肌内脂肪(大理石花纹)仍是提高延边黄牛牛肉品质的挑战。而且,在外来商业肉牛的竞争下,延边黄牛的经济价值也因其胴体中较低的肌内脂肪(大理石纹)含量而严重降低。因此,提高肌内脂肪含量便成为改善肉类品质的关键,而脂肪生成分子机制研究可为改善肉质提供宝贵的信息。
肌内脂肪的沉积涉及一个连续的过程链,包括前脂肪细胞的增殖、分化和成熟[2]。胰岛素是调控机体能量代谢的主要激素之一,可以显著提高脂肪细胞分化标志基因FAS和ACC1的表达,有效促进前体脂肪细胞的分化[3-4],且存在明显的剂量-效应关系。过氧化物酶体增殖物激活受体-PPARγ(peroxisome proliferator-activated receptor gamma,PPARγ)是调控脂肪细胞形成、脂肪组织发育和维持成熟脂肪细胞功能的关键调控因子,而环格列酮(ciglitazone)是PPARγ主要的激活剂之一,可通过激活PPARγ基因从而有效促进脂肪生成相关基因的表达[5-6]。单不饱和脂肪酸是前脂肪细胞增殖与分化过程重要的调控因子。其中油酸就是比其他长链脂肪酸更能充分诱导脂肪生成的一种重要的单不饱和脂肪酸(MUFA),不但可以从牛的胃肠道吸收,还可以通过内源性生成[7-8]。而且油酸作为PPARγ的天然配体,可促进PPARγ向细胞核内的转位,调节下游基因的表达[9]。
此前,我们已经确定了胰岛素、油酸和环格列酮三者对延边黄牛前脂肪细胞分化中主要基因表达的影响,为了继续研究三者在调控前脂肪细胞分化机制中的作用,本试验以延边黄牛为研究对象,利用胶原酶消化法,在无菌条件下分离获得延边黄牛前脂肪细胞并建立体外培养体系,通过油红O染色和实时荧光定量PCR技术比较分析胰岛素和PPARγ激动因子对脂肪细胞分化过程中相关基因表达及脂滴生成情况的影响。试验材料虽然是采集延边黄牛皮下脂肪组织的前脂肪细胞,但目标是为了更好地了解调节脂肪生成的因素,以便将所发现应用于肌内脂肪细胞的发育研究。
1 材料与方法
1.1 主要试剂FBS、PBS、DMEM、青链霉素混合液(双抗)、胰蛋白酶、TRIzol Reagent、环格列酮、油酸、油红O染色试剂盒、甘油三酯试剂盒均购自长春朝阳区迈希试剂耗材经销处;GAPDH、PPARγ、SCD、SREBP1、LPL、C/EBPβ引物购自英潍捷基(上海)贸易有限公司;其他试剂均为国产分析纯。
1.2 培养液的配置基础培养液:DMEM+10% FBS+1% PS(2.5 mg/L两性霉素+50 mg/L庆大霉素);分化培养液:DMEM+5% FBS+1% PS(2.5 mg/L 两性霉素+50 mg/L庆大霉素);冻存液:DMEM+10% DMSO+10% FBS;清洗液:1% PS+PBS+二酶(2.5 mg/L两性霉素+50 mg/L庆大霉素);消化液:DMEM+1% PS+2 g/L胶原酶(collagenase)+1% BSA。
1.3 原代脂肪细胞的分离培养从延边黄牛皮下脂肪组织中提取原代脂肪细胞。取样部位先用75%乙醇清洗,然后用无菌的手术刀小心取下100 g脂肪组织,用清洗液清洗2~3次后立即转移到细胞培养室。
将取回的脂肪组织放置于灭菌的培养皿中,用清洗液清洗4~5次,用无菌钳和剪刀去除血管和筋膜等,并将其剪碎,再用胶原酶进行酶消化,并在37℃水浴中持续1 h。每隔5 min振荡1次,以保证充分消化,用等体积的培养液中和该消化混合物后,使用100 μm的细胞筛过滤。将过滤后的滤液1 500 r/min 离心10 min,去上清液,加入10 mL PBS清洗细胞颗粒,用70 μm细胞筛再次过滤,1 500 r/min 离心10 min,去除上清。最后将细胞悬浮于培养基中,置于37℃、5% CO2的恒温培养箱中培养,每隔48 h用PBS冲洗3次,以去除碎屑和游离漂浮细胞后更换培养液。
1.4 传代培养及分化处理待细胞增殖到培养皿底面的70%~80% 时,吸出培养液,用PBS清洗2~3次,加入2 mL 0.25%的胰蛋白酶,放入37℃ 恒温培养箱中消化2~3 min,加2 mL的基础培养液终止消化。将细胞悬液转移到离心管中, 1 500 r/min, 离心5 min,弃上清,再次用基础培养液悬浮细胞后传代至新的培养皿中,继续进行培养。
待细胞增殖到70%~80%后进行不同的分化处理。CON组:10 mg/L胰岛素;IC组:10 mg/L胰岛素+10 mg/L环格列酮;IO组:10 mg/L胰岛素+100 μmol/L油酸;ICO组:10 mg/L胰岛素+10 mg/L 环格列酮+100 μmol/L 油酸。每个处理重复3次,置于 37℃、5% CO2恒温培养箱中培养,每隔24 h换1次分化培养液,分化处理96 h后,进行后续试验。以上操作均在无菌条件下进行。
1.5 油红O染色参照油红O染色试剂盒(G1262)进行,用PBS缓冲液冲洗细胞2~3次后,用细胞色素固定剂在室温条件下将其固定30 min,再使用PBS缓冲液冲洗1次,60%异丙醇清洗 20~30 s后,涂上油红O染色(工作液),在室温下密闭染色 10~15 min,再加入60%异丙醇洗去染液,用PBS缓冲液冲洗2~3次,加入Mayer苏木素染色液复染核 1~2 min,加 ORO Buffer处理 1 min,镜检采集细胞图像,观察脂质滴。
1.6 甘油三酯含量的测定使用普利来甘油三酯测定试剂盒进行甘油三酯含量的测定。用PBS清洗细胞2次,2 mL胰蛋白酶消化2~3 min后,加入同等体积的培养液终止消化,移液枪反复吹打制成细胞悬液,1 500 r/min,离心5 min,弃上清,反复2次,加入200 μL裂解液充分混匀,室温条件下静止10 min,使细胞中甘油三酯完全释放于裂解液中,70℃金属浴中10 min,2 000 r/min离心5 min并吸取上清。将10 μL待测样品(每个样品3个重复)和190 μL工作液加入到96孔板中,放置于37℃培养箱中孵育10 min,之后使用酶标仪在570 nm波长下测定吸光值,再根据标准曲线计算甘油三酯含量。
标准液浓度:0,200,400,600,800,1 000,1 200 mmol/L,酶标仪于570 nm波长下测定吸光值并绘制标准曲线。
1.7 相关基因mRNA表达量检测用TRIzolTM试剂从细胞中提取总RNA,通过D260和D260/D280检查提取总RNA的完整性。用TRIzol法提取各试验组脂肪细胞总RNA,cDNA的构建参照TaKaRa的反转录试剂盒进行。GAPDH作为内参基因,引物序列参照表1,由生工生物工程(上海)股份有限公司合成。
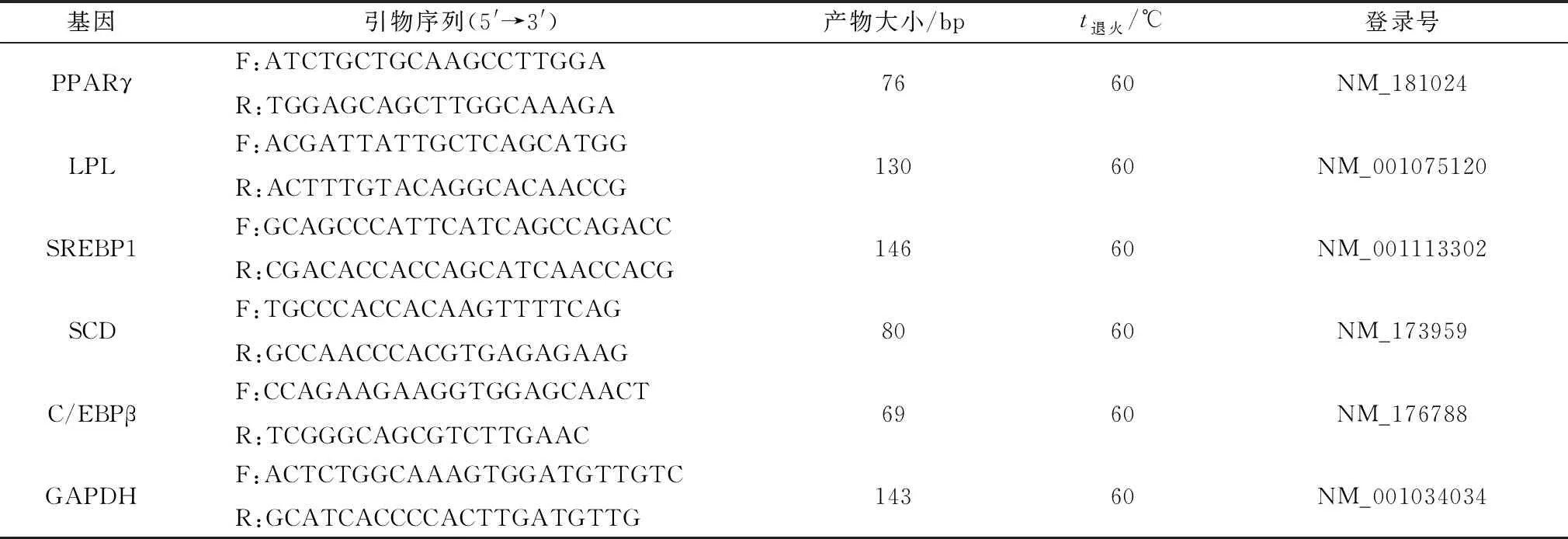
表1 实时荧光定量PCR引物信息
PCR反应条件为:95℃条件下预变性 15 s,95℃变性 5 s,55℃退火30 s,72℃延伸30 s,共40个循环。
1.8 统计分析使用 2-△△Ct计算相对mRNA表达量,利用Graph Pad Prism 6.0软件进行数据分析与作图,差异在P<0.05处有统计学意义,作为差异显著性判断标准。
2 结果
2.1 延边黄牛前脂肪细胞生长趋势应用MTS比色法每隔24 h测定1次490 nm的吸光值,每次取3个孔分别进行测定并绘制延边黄牛前脂肪细胞生长曲线图。由图1可见,延边黄牛前脂肪细胞具备稳定的生长特性,培养2~6 d,细胞活力显著增强,进入指数生长期,6~10 d细胞的增殖速度明显减慢,进入平台期,之后逐渐退出细胞增殖周期,因此可用于后续试验。
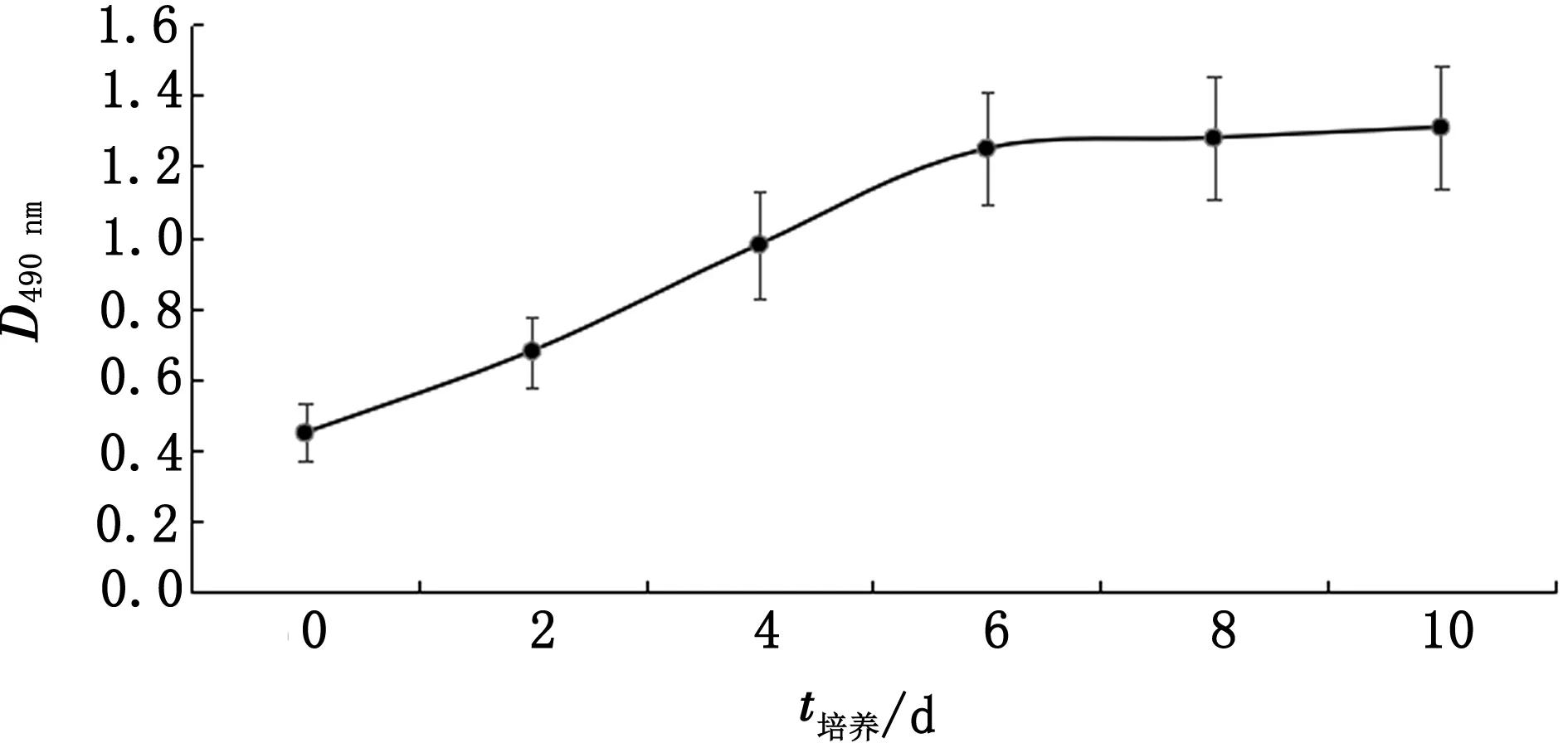
图1 延边黄牛前脂肪细胞生长曲线
细胞进行体外培养,将消化酶处理后的细胞悬液接种到新的培养皿,细胞缩成小圆球形(图2A),培养1 d后,有部分呈不规则三角形的细胞已经贴壁(图2B),之后的培养过程中,贴壁细胞逐步开始伸展变形,最后形成了均一、完整的单室脂肪细胞,大多数呈狭长梭形的成纤维样细胞形态(图2C)。继续培养延边黄牛前脂肪细胞,细胞增殖速度相对减缓,且细胞增殖到80%以上,准备开始进入分化阶段(图2D)。

A.复苏0 h的细胞悬浊液,含有杂质细胞及肌肉组织碎片;B.培养1 d后细胞开始贴壁生长,呈长梭型;C.细胞进入快速生长期,大量增殖;D.细胞增殖到80%
2.2 不同处理对延边黄牛前脂肪细胞脂滴形成及甘油三酯含量的影响油红O染色结果(图3)显示,在分化处理96 h后,与对照组相比,各试验组均有脂滴形成,IC组只有少量脂滴形成,而IO和ICO组形成的脂滴数量较多,且脂滴形态较大,说明胰岛素和油酸共同处理可有效促进延边黄牛前脂肪细胞脂滴的形成。
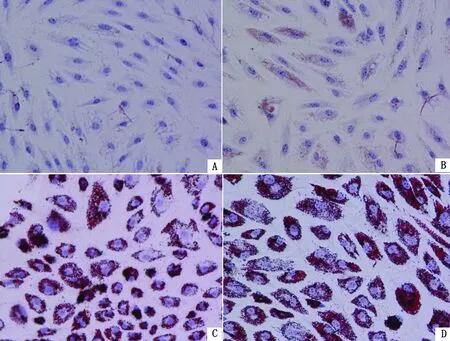
A.对照组(CON)组;B.IC组;C.IO组;D.ICO组
由图4可知,与对照组相比,各试验组均可提高甘油三酯含量,其中IO和ICO组显著高于对照组和IC组(P<0.05),且ICO组含量最高。
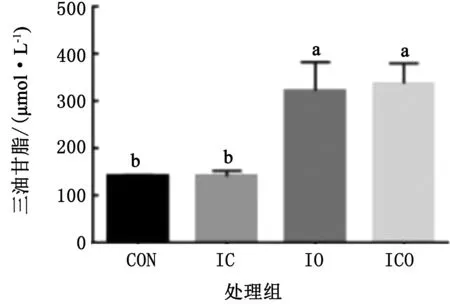
注:组间标注不同小写字母示差异显著,P<0.05。下同
2.3 不同处理对延边黄牛前脂肪细胞分化相关基因表达的影响由图5可知,各试验组间PPARγ基因的mRNA表达量差异显著(P<0.05),与对照组相比,IC和ICO试验组显著高于对照组(P<0.05),IO组与对照组间差异不显著(P>0.05)。IC和ICO组SCD基因mRNA表达量高于对照组,其中IC组与对照组间差异显著(P<0.05),而IO组SCD基因mRNA的表达量最低。各试验组间LPL基因mRNA表达量具有显著性差异(P<0.05),与对照组相比,IC和ICO组显著上调LPL基因的mRNA表达量(P<0.05),而IO组却显著下调了LPL基因的mRNA表达量(P<0.05)。相比于对照组,各试验组均显著提高了C/EBPβ基因mRNA表达量(P<0.05),其中IO组表达量最高。胰岛素分别与环格列酮和油酸处理,可明显增加SREBP1基因的mRNA表达量,且显著高于对照组和ICO组(P<0.05),而相比于对照组,ICO组却降低了该基因的表达量,但差异不显著(P>0.05)。
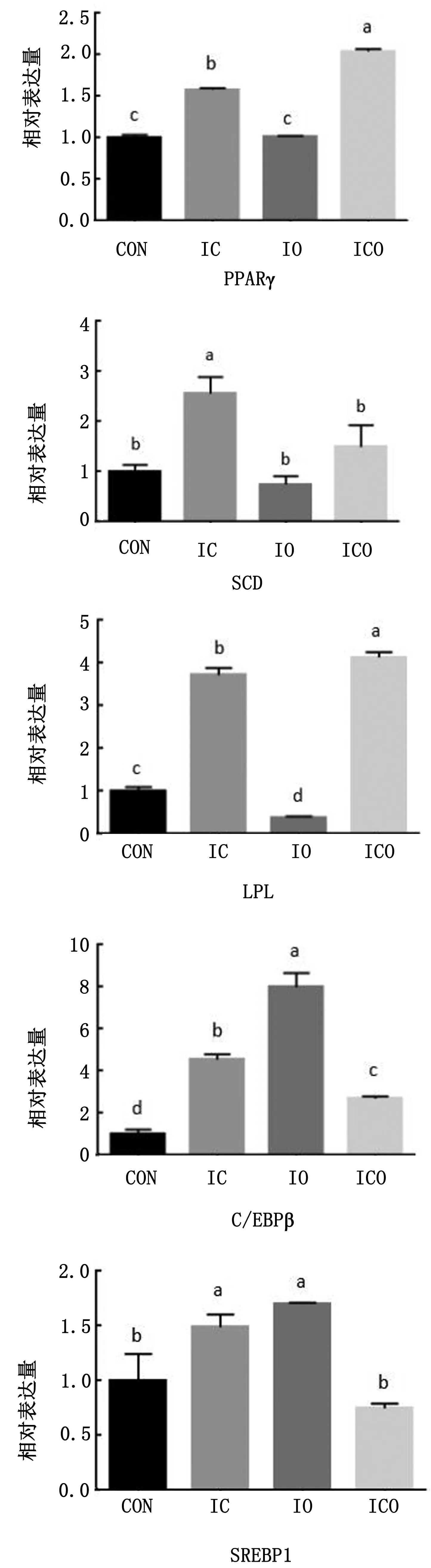
图5 PPARγ、SCD、 LPL、CEBP/β和SREBP1基因mRNA相对表达量
3 讨 论
目前细胞增殖的研究方法比较多,MTS比色法是改良的MTT比色法,与 MTT 或 INT 法相比,对细胞毒性更小,步骤更为简便[10-11]。所以本试验延边黄牛前脂肪细胞生长曲线的检测采用MTS比色法,以期反映细胞生长的基本规律。研究表明,前脂肪细胞生长曲线划分为潜伏期、指数生长期、平台期以及退化衰亡期[12]。本试验检测结果显示,培养2~6 d,细胞活力显著增强,进入指数生长期;6~10 d 脂肪细胞进入平台期,增殖速度明显减慢。本试验没有出现潜伏期和衰亡期,这可能是由于测定时间点较少而导致的。培养8 d后细胞生长速度并未出现明显减缓,这可能是由于培养基中营养成分依然满足高密度细胞的生长,细胞竞争性生长还未出现,所以细胞没有出现凋亡的现象。
细胞主要是以甘油三酯和胆固醇酯的形式来储存多余的能量[13]。油红O为脂溶性染料,可高度溶解于脂肪内,使组织内甘油三酯等中性脂肪着色。前期试验研究结果显示,环格列酮和油酸单独处理均可促进脂滴的形成,而环格列酮和油酸共同处理,脂滴数量肉眼判断虽不如环格列酮组和油酸组多,但脂滴大小却明显大于环格列酮组和油酸组,而胰岛素单独处理却不能促进脂滴的形成[5,20,22],这与本试验研究结果一致。本试验中,在与前期试验条件相一致的条件下,胰岛素、油酸和环格列酮三者共处理组无论是脂滴的数量还是大小均优于其他试验组,这说明胰岛素的添加,可提高油酸和环格列酮共处理本身对脂滴形成的影响效果。本试验油红O染色结果与甘油三酯结果相一致。
胰岛β细胞分泌的内分泌激素——胰岛素,在机体的血糖调节、能量代谢及细胞增殖、细胞分化等多个方面扮演重要角色[14-16]。在脂肪细胞中,胰岛素是调节脂肪形成的主要激素,在抑制脂肪分解的同时,可诱导血液中脂肪的生成和对脂肪酸的摄取[3,17]。GUO等[18]研究发现胰岛素可通过与前脂肪细胞膜表面的 IR 结合,激活PPARγ2 促进脂肪细胞的分化。FARMER等[19]发现胰岛素信号通路在胰岛素及cAMP激活剂的诱导刺激下,可促进多个成脂转录相关因子SREBP-1c、C/EBPβ、C/EBPα和PPARγ2 的表达。金鑫等[20]研究结果表明,胰岛素处理牛的前脂肪细胞,虽不能促进脂滴的形成,但可显著提高PPARγ基因的表达量。前期试验表明,与油酸和环格列酮单独处理相比较,两者共同处理并不会显著促进脂肪生成基因的表达。而本试验中,胰岛素添加后,胰岛素与油酸、环格列酮共处理对PPARγ基因表达的促进作用却明显高于胰岛素分别与油酸和环格列酮共处理对其的影响,这可能是由于胰岛素促进了油酸和环格列酮的叠加使用导致的PPARγ信号通路上某些基因的表达。
游离脂肪酸是甘油三酯的重要组成成分,尽管其在血液中浓度很低,但却是脂类代谢中最活跃的部分。大量研究结果表明游离脂肪酸能够促进脂肪前体细胞的分化。GARCIA等[21]研究发现来源于牛饲料的不饱和脂肪酸和类胡萝卜素能够提高PPARγ的表达,是PPARγ的天然活化剂。金鑫等[22]研究发现油酸不仅对脂滴的形成有着明显的促进作用,同时还显著提高了PPARγ和SREBP1基因的表达量。张军芳等[23]研究结果表明,添加油酸和棕榈油酸等不饱和脂肪酸可增加脂肪合成相关基因PPARγ、SREBP1、C/EBPα表达,抑制SCD基因表达。张靖伟等[24]在细胞分化的过程中,用不同浓度(20,40,60 μmol/L)的油酸分别作用细胞3 d,应用油红O染色和胞内甘油三酯测定法分析发现油酸可以在体外显著地促进细胞胞浆中脂质的积累,促进脂肪干细胞成脂分化。宋沙等[25]研究发现油酸能促进肉牛前脂肪细胞的分化或提高脂肪细胞内的脂肪含量,提高牛肉的大理石纹等级和风味。以上研究结果均表明,油酸可促进相关脂肪生成基因的表达量,从而促进前脂肪细胞的分化。而本试验中,在油酸处理中添加胰岛素以后,在抑制SCD基因表达同时,也抑制了LPL基因的表达,但是却提高了PPARγ、SREBP1和C/EBPβ基因mRNA的表达量,这与前人研究结果相似,说明胰岛素和油酸共处理不会影响油酸本身对前脂肪细胞分化的影响,有可能还会提高促进分化的效果。
PPARγ的特异性配体主要是噻唑烷二酮类化合物,而环格列酮就属于噻唑烷二酮类药物(TZDs),是PPARγ最主要的激活剂之一,其能够将PPARγ活化,并提高相关脂肪因子的表达从而提高肌内脂肪沉积水平[26]。饲粮中添加PPARγ激活剂会使猪肌内脂肪含量和大理石纹评分等级显著提高[27]。本试验前期研究结果表明环格列酮单独处理可提高PPARγ、C/EBPβ和SCD基因的相对表达量。而在本试验中,环格列酮和胰岛素共处理,均可显著提高PPARγ、SCD、C/EBPβ、LPL以及SREBP1的表达量,这说明胰岛素的添加并没有抑制环格列酮对PPARγ的激活效果。
综上所述,我们推测胰岛素和PPARγ激动剂油酸和环格列酮共处理对延边黄牛前脂肪细胞分化的促进作用要高于各因素单独处理或两两共同处理的效果,这一研究结果为延边黄牛脂肪代谢的研究提供一个新视角。但胰岛素是如何与环格列酮和油酸相互作用来调控脂肪细胞成脂分化过程的具体的调控机制有待进一步研究。
猜你喜欢
家畜生态学报(2022年2期)2022-02-28 01:52:28
延边大学学报(社会科学版)(2020年6期)2021-01-14 00:45:24
新农业(2020年18期)2021-01-07 02:17:28
复旦学报(医学版)(2019年4期)2019-08-07 04:55:38
外国问题研究(2018年4期)2018-03-06 07:53:44
韩国语教学与研究(2017年2期)2017-10-20 05:20:48
中国民族医药杂志(2016年4期)2016-05-09 07:41:11
听力学及言语疾病杂志(2015年5期)2015-12-24 01:47:05
精细石油化工(2015年3期)2015-12-14 09:07:40
当代音乐(下旬刊)(2015年5期)2015-05-30 05:21:05
