
三阴性乳腺癌组织中FOXJ1、SATB1的表达变化及临床意义
2021-07-29 05:50付飞国王忆丽雷秋模
中国医药导报 2021年16期
付飞国 王忆丽 雷秋模
南昌市第三医院乳腺外科,江西南昌 330009
乳腺癌的死亡率与发病率位居女性肿瘤疾病首 位,近几年乳腺癌的发病率和死亡率呈逐年上升,严重影响女性的身心健康[1]。而三阴性乳腺癌在所有乳腺癌中死亡率最高,5 年预后生存率不足30%[2]。目前主要依赖TNM 分期对三阴性乳腺癌患者预后进行判定[3]。叉头框蛋白J1(FOXJ1)是FOX 蛋白家族成员,是一种转录因子,在胚胎发育、免疫调节以及肿瘤发生发展中起重要作用[4-5]。核基质结合区结合蛋白质1(SATB1)在胰腺癌和前列腺癌等肿瘤中表达较高,参与基因转录及转录后修饰过程,在肿瘤转移及侵袭调控中起重要作用[6-7]。有研究发现,FOXJ1 和SATB1 在胃癌中均出现表达,且共同参与胃癌的发生和发展过程[8],然而目前临床上关于三阴性乳腺癌FOXJ1 和SATB1 的研究报道较少,因此笔者进行此研究。现报道如下:
1 资料与方法
1.1 一般资料
选择2014 年2 月—2015 年3 月南昌市第三医院行三阴性乳腺癌根治术治疗的98 例患者为研究对象。纳入标准:①临床资料完整;②病理学诊断确诊为三阴性乳腺癌,符合三阴性乳腺癌的临床诊断标准[9]。排除标准:①全身免疫性缺陷及肝肾功能障碍;②患者并发其他恶性肿瘤;③未行肿瘤根治术。本研究经医院医学伦理委员会批准。
1.2 试剂与方法
手术切除的肿瘤组织及相应的癌旁组织(距离肿瘤边缘>2 cm)先进行常规的中性甲醛(诸暨市和谐科技有限公司,产品编号:59400402551)固定,然后切取部分进行酒精梯度脱水,脱水结束后放于二甲苯中透明,然后用生物组织包埋机(北京伊莱尔医疗设备有限公司,型号:MB2271)对组织进行常规的石蜡包埋,制备组织切片。组织切片制备成功后先放于烘箱加热脱蜡,然后放于二甲苯和梯度酒精进行脱蜡水化,将水化后的组织切片置于37℃湿盒中使用3%过氧化氢(广东恒健制药有限公司,批号:0299001)孵育20 min,将孵育完成的组织切片放于高压锅中进行抗原修复后在37℃封闭液中孵育30 min。孵育结束后在切片组织周围用组化笔画圈并在组织上滴加Anti-FOXJ1抗体或Anti-SATB1(德国ACAM 生物科技有限公司,货 号:ab235445、ab109122)抗体,抗体加完后置于4℃冰箱过夜。次日清洗后滴加二抗,然后置于37℃恒温箱孵育30 min,最后用二氨基联苯胺法(四川海诺威科技有限公司,货号:91-95-2)进行显色,并用苏木精(南京春秋生物工程有限公司,货号:517-28-2)进行复染,最后对组织切片进行脱水、透明和固定。染色完成后将组织切片置于倒置生物显微镜(上海市德意曼医疗器械有限公司,型号:DYM-5100)下观察染色结果。根据细胞阳性数可分为5 个等级,每个等级对应记0~4 分,阳性细胞比例≤5%记0 分,5%<阳性细胞比例<25%记1 分,25%≤阳性细胞比例<50%记2 分,50%≤阳性细胞比例<75%记3 分,阳性细胞比例≥75%记4 分。根据染色强度将切片分为4 个等级,无色记0 分,淡黄色记1 分,棕黄色记2 分,棕褐色记3 分,最后将阳性细胞数评分和染色强度评分相乘得到最终评分并按等级分为阴性和阳性,其中0~4 分为阴性、5~12 分为阳性[10]。
通过复查和电话对患者进行随访,随访截止时间为2020 年3 月,根据生存情况统计所有患者生存时间。
1.3 统计学方法
采用SPSS 20.0 统计学软件进行数据分析,计数资料用例数或百分率表示,组间比较采用χ2检验;Kaplan-Meier 法进行生存分析,生存率比较采用Logrank 检验。FOXJ1 和SATB1 表达相关性用Pearson 法分析。以P <0.05 为差异有统计学意义。
2 结果
2.1 三阴性乳腺癌组织及癌旁组织中的SATB1 和FOXJ1 的表达
FOXJ1 主要表达在细胞核,SATB1 主要表达在细胞膜细胞外。免疫组化结果显示,三阴性乳腺癌组织中FOXJ1 阳性率为31.63%(31/98)低于癌旁组织的60.20%(59/98),差异有高度统计学意义(χ2=16.107,P <0.001),见图1A~B。三阴性乳腺癌组织中SATB1阳性率为72.45%(71/98),高于癌旁组织的23.47%(23/98),差异高度有统计学意义(χ2=47.099,P <0.001),图1C~D。
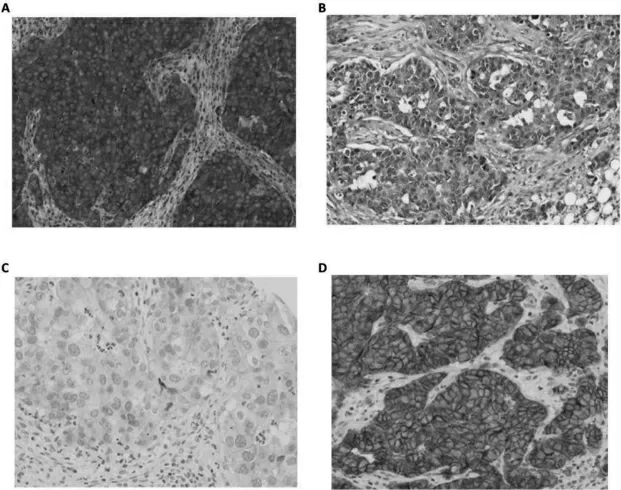
图1 FOXJ1 和SATB1 在不同组织中的表达情况(免疫组化,400×)
2.2 三阴性乳腺癌组织中FOXJ1 和SATB1 表达和临床参数的关系
三阴性乳腺癌组织中,不同FOXJ1 表达与TNM分期及肿瘤分化程度有关(P<0.05 或P<0.01);不同SATB1 表达与肿瘤浸润深度、TNM 分期以及淋巴结是否转移有关(P <0.05 或P <0.01)。见表1。

表1 三阴性乳腺癌组织中FOXJ1 和SATB1 表达与临床参数的关系(例)
2.3 三阴性乳腺癌组织中FOXJ1 和SATB1 的表达相关性
三阴性乳腺癌中,FOXJ1 表达与SATB1 表达呈负相关(r=-0.625,P <0.05)。
2.4 三阴性乳腺癌组织中FOXJ1 和SATB1 的表达和患者预后之间的关系
对数据进行Kaplan-Meier 生存曲线分析,结果显示,SATB1 阳性患者5 年总生存率为35.2%(25/71)低于阴性患者的59.3%(16/27),差异有高度统计学意义(χ2=14.170,P <0.001),图2A。FOXJ1 阳性患者5 年总生存率为64.5%(20/31),高于阴性患者的31.3%(21/67),差异有高度统计学意义(χ2=22.386,P <0.001),图2B。

图2 三阴性乳腺癌组织FOXJ1 和SATB1 表达与患者预后的关系
3 讨论
乳腺癌是女性常见的恶性肿瘤,并且其发病率有不断升高的趋势[11]。三阴性乳腺癌的发生发展不依赖于体内激素的表达,对靶向治疗药物的治疗效果较差,只能通过放化疗延长患者生存期,预后不佳[12-13]。乳腺癌的发生涉及多种癌基因的异常激活及抑癌基因的失活,引起肿瘤细胞的恶性增殖及转移等[14-15]。寻找导致三阴性乳腺癌发生发展的关键驱动基因,有助于寻找新的治疗药物及治疗方案。
FOXJ1 基因位于人类染色体17q25.1,该基因编码蛋白属于转录因子叉头框家族的成员,具有调节控制纤毛产生及运动的生物学功能,参与调控肿瘤转移、侵袭、增殖等恶性生物学过程[16-18]。本研究结果显示,三阴性乳腺癌组织中FOXJ1 表达下降,提示FOXJ1在三阴性乳腺癌中可能作为抑癌基因,参与三阴性乳腺癌的发生发展过程。有学者发现在MDA-MB-231、HCC1937 和HCC1806 等三阴性乳腺癌细胞系中FOXJ1 有表达下调的现象[19],与本研究的结果一致。目前对于FOXJ1 表达下调的具体分子机制尚不清楚,可能与其他转录因子的转录调控和蛋白酶体降解有关。在胃癌的研究中发现,miR-6852 可以结合FOXJ1 的信使RNA,进而抑制FOXJ1 表达及功能,促进胃癌细胞的转移和增殖[20]。同时,TNM 分期越高、肿瘤分化程度越差的三阴性乳腺癌组织中FOXJ1 表达越低。其机制可能是肿瘤中FOXJ1 能够下调细胞外信号调节激酶通路蛋白如丝裂原活化的蛋白激酶1等的表达,抑制细胞外信号调节激酶通路的活化,发挥抑癌基因的功能,FOXJ1 的表达下调激活细胞外信号调节激酶通路下游靶基因,如c-met、血管内皮生长因子的表达,促进肿瘤细胞的增殖浸润及转移,导致肿瘤进展[16]。FOXJ1 低水平能够反映患者的不良预后,能够成为新的三阴性乳腺癌预后相关标志物。其原因可能与FOXJ1 的低表达多发生在恶性程度较高的肿瘤当中,FOXJ1 表达降低导致对丝氨酸-苏氨酸激酶信号通路活化抑制作用减弱,进而引发肿瘤细胞增殖及侵袭转移,最终加快癌症进展,缩短预后生存[21]。
SATB1 基因位于3p24.3,该基因编码蛋白作为一种基质蛋白,参与募集染色质重塑因子以调节染色质结构。研究显示,SATB1 在促进肿瘤上皮-间质转化中起到重要的调节作用,并调控细胞氧化应激等病理生理过程[22]。本研究结果显示,三阴性乳腺癌组织中SATB1 阳性占比高于癌旁组织(P <0.01),其机制可能与结合到SATB1 基因启动子上的组蛋白乙酰化水平升高有关,如在结直肠癌当中,乙酰化修饰的组蛋白结合到SATB1 基因启动子区,促进其转录,进而起到促进肿瘤进展的作用[23]。此外,不同SATB1 表达与肿瘤浸润深度、TNM 分期以及淋巴结是否转移有关联(P <0.05 或P <0.01),提示SATB1 能够作为一种促癌基因,促进肿瘤的恶性进展。研究报道显示,在乳腺癌中SATB1 能够促进E 型钙黏蛋白的表达,使得乳腺癌细胞易发生局部浸润及淋巴结转移,进而促进肿瘤发生发展[24]。此外,SATB1 高表达患者的预后较差,癌组织中SATB1 的表达程度可反映三阴性乳腺癌患者的预后生存情况。
本研究结果显示,三阴性乳腺癌组织中FOXJ1 和SATB1 表达呈负相关,提示在三阴性乳腺癌发生发展过程中FOXJ1 和SATB1 均起到一定调节作用。研究表明,喉鳞状细胞癌中,FOXJ1 作为转录抑制因子,能够与组蛋白结合并抑制其乙酰化,组蛋白乙酰化水平下调而抑制SATB1 启动子区转录起始复合物的形成,抑制SATB1 表达,进而抑制肿瘤进展[25]。但三阴性乳腺癌中SATB1 是否是FOXJ1 的靶基因以及二者在三阴性乳腺癌发生发展中是否发挥关键作用仍需进一步深入探究。
综上所述,三阴性乳腺癌组织中FOXJ1 低表达,SATB1 高表达,共同参与三阴性乳腺癌的发生发展过程,并与三阴性乳腺癌患者的预后相关,有可能成为三阴性乳腺癌患者预后预测和治疗的潜在标志物。
猜你喜欢
中国医学影像学杂志(2018年9期)2018-10-17
中国医药指南(2017年3期)2017-11-13
电信科学(2016年11期)2016-11-23
中国组织化学与细胞化学杂志(2016年3期)2016-02-27
中国组织化学与细胞化学杂志(2016年3期)2016-02-27
医学研究杂志(2015年12期)2015-06-10
中国当代医药(2015年17期)2015-03-01
西安交通大学学报(医学版)(2014年5期)2014-06-23
中国药理学通报(2014年2期)2014-05-09
检验医学(2012年6期)2012-09-11
