
木腐真菌对松材线虫病疫木处理初探
2021-07-28 03:52蒙海勤叶建仁王旻嘉曹伊扬
南京林业大学学报(自然科学版) 2021年4期
蒙海勤,叶建仁,王旻嘉,曹伊扬
(南京林业大学林学院,南方现代林业协同创新中心,江苏 南京 210037)
松材线虫病是由松材线虫(Bursaphelenchusxylophilus)引起的一种毁灭性病害,具有寄主范围广、致病力强、染病寄主死亡速度快等特点[1-3]。该病于1905年在日本首次发现,至2018年已在我国18个省发生[4-5],目前全世界已有52个国家将其列为重要的检疫性病害,其中在中国和日本发生最严重[6-10]。几十年来各国学者一直致力于松材线虫病的相关研究,但有关松材线虫病的致病机理仍不完全清楚。在防治松材线虫病的关键环节中,疫木就地或就近处理技术有待重大突破[11-12]。防治实践中,检疫、监测、疫木处理、媒介昆虫防治等环节中的关键在于疫木处理[13],当前疫木处理主要采用粉碎或焚烧等方法,但实际应用中在一些地区存在如交通严重不便、人员很难到达等困难,导致疫木处理质量难以保证[14-15]。
在自然界中普遍存在通过改变疫木内微生境,从而使疫木内松材线虫数量减少的微生物。陈瑶等[16]利用木腐菌硫磺菌(Laetiporussulphureus)处理病死树伐桩,发现该木腐菌可有效分解伐桩,并能够有效抑制伐桩内的松材线虫数量,伐桩的质量与松材线虫含量成正比。邓习金等[17]研究发现,木腐菌硫磺菌和茯苓(Poriacocos)等可有效减少疫木内松材线虫的数量,可作为处理松材线虫病疫木的潜力菌株。本研究拟筛选出既能抑制松材线虫生长繁殖又能降解黑松木质的优良真菌菌株,经大量培养后接种于松材线虫病疫木中,以期实现松材线虫病的侵染循环在疫木环节被阻断的目标,为松材线虫病的有效防控提供科学参考。
1 材料与方法
1.1 试验材料
从南京林业大学下蜀林场和江苏省句容市摇令口工区感染松材线虫病不同腐朽阶段的病死松木上分离纯化获得B18、C11、C25、C28、D16、D17、D23等7个真菌菌株(未鉴定,不同腐朽阶段对菌株的命名不一样,第1个阶段分离获得的菌株以A1、A2命名,第2个阶段以B1、B2命名,以此类推)并置于南京林业大学森林病理实验室保存。绿色木霉(Trichodermaviride,编号Tv)购自广东省微生物菌种保藏中心,黄孢原毛平革菌(Phanerochaetechrysosporium,编号Phc)购自中国典型培养物保藏中心(武汉大学保藏中心),茯苓(Poriacocos,编号Pc)取自安徽省六安金寨县金山寨菌种厂,灰葡萄孢菌(Botrytiscinerea,编号Bc)保存于南京林业大学森林病理实验室。
松材线虫强致病力虫株AMA3,保存于南京林业大学森林病理实验室。
试验疫木来源于江苏省句容市摇令口工区感染松材线虫病14年生的黑松病死木。
1.2 试验方法
1.2.1 松材线虫在木腐真菌平板上的生长繁殖
分别将2 000条松材线虫接种至培养成熟的各种木腐真菌菌落平板上,每种真菌接种3皿,以灰葡萄孢菌培养的松材线虫作为阳性对照,置于25 ℃恒温培养箱中培养10 d。采用贝尔曼漏斗法对平板中的松材线虫进行分离,在光学显微镜下对松材线虫进行计数,最后分别计算各种真菌平板上松材线虫数量变化情况。
1.2.2 木腐真菌对黑松木质量和纤维素的影响
将木腐真菌接种至平板中,待菌丝即将长满平板时放入已灭过菌的小木块,以不接菌的PDA平板作为对照,每处理3皿。在15 ℃下培养120 d后,去掉木块上的菌丝并将木块烘干至质量恒定,并称质量,按如下公式计算木块的质量损失率:
式中:x1表示试样降解后的质量损失率,%;ω0表示试样降解前的绝干质量,g;ω1表示试样降解后的绝干质量,g。
取绝干后的木块,将其粉碎,利用化学法测定木质纤维素含量的变化,降解木材各组分降解率的计算依照以下公式:
式中:x10表示降解木材试样中某组分降解率,%;x0表示健康试样中某成分含量,g;x表示降解木材试样中某成分含量,g;x5表示试样降解后质量损失率,%[18]。
1.2.3 木腐真菌对疫木内松材线虫数量消长的影响
首先制作液体PDA培养基,高温高压灭菌后接入已活化的木腐真菌置于25 ℃,转速为150 r/min的恒温摇床中振荡培养10 d以获得液体培养菌丝。固体培养菌丝的制作配方为:松木屑2 000 g、稻壳100 g、小麦粉200 g、葡萄糖20 g、七水硫酸镁4 g、磷酸二氢钾2 g、硫酸铵4 g、水1 000 mL,这些原料混匀后分装200 g置于17 cm×33 cm的耐高压高温聚乙烯透气袋中,于105 ℃的高压灭菌锅中灭菌4 h,连续重复灭菌3次,灭菌时间达到12 h,灭好菌后待营养袋中的材料冷却至室温,将液体培养好的木腐真菌按照每袋100 mL的菌种量接种于营养菌袋中,置于恒温为25 ℃的生化培养箱中培养,培养30 d使用。
用油锯伐倒疫木,去除侧枝后将主干运至南京林业大学下蜀林场,利用电锯将疫木主干截成每段约60 cm长的木段,在木段上每隔15 cm用电钻打孔,每木段正反两面交错分布共计3个孔,每个孔深度40 mm、直径35 mm,并对圆孔进行编号,木段两端为1号和3号,中间为2号。将所有木段进行编号、分类。
在疫木木段上接种木腐真菌。疫木伐倒12 d后,采用由液体培养和固体培养两种方式培养的菌丝进行接种。接种前先在接种孔周围喷洒75% 的酒精,待酒精全部挥发后进行接种试验,试验设置5个不同菌株和2种培养菌丝接种方式,每个处理接种6个木段(基本来源于不同株),分别以接种液体PDA和固体培养基作为空白对照。将事先培养好的木腐真菌接种至疫木小孔中,液体摇培菌丝每孔接种40 mL,固体菌丝每孔接种20 g,接种后利用保鲜膜封口以保温保湿和防止其他杂菌污染。由于接种液体培养菌丝受取样时间限制,每个处理不是所有木段都有天牛蛀道,而且蛀道数量不一,导致收集到的样本数量不同,因此未做显著性分析。
接种木腐真菌后,分析不同处理的疫木中松材线虫种群的变化。定期观察接种于木段上的木腐真菌生长情况,发现天牛即将羽化(即在接种150 d)时分别钻取接种孔正面木样,然后用斧头将整个木段劈开,收集天牛蛀道周围的木样,分析蛀道周围木样内松材线虫数量(用贝尔曼漏斗法分离其中线虫数量)。同时收集漏斗上的木屑并用信封包好,放置于105 ℃烘箱中烘至质量恒定。
1.3 数据处理
利用Excel 2010和SPSS 24.0处理和分析数据,Origin 2018 软件作图。
2 结果与分析
2.1 不同木腐真菌对松材线虫生长的抑制分析
分别将2 000条松材线虫接入不同木腐真菌的平板上培养10 d后,每个平板上松材线虫数量见图1。由图1可知,松材线虫在茯苓(Pc)、黄孢原毛平革菌(Phc)、B18、C11、D16 这5种真菌的平板上几乎不能生长,从茯苓的菌落中未分离到松材线虫,在其他4种真菌的菌落上也仅分离到极少量的松材线虫。多重比较结果表明,这5种木腐真菌对松材线虫生长繁殖的影响不具有显著性差异。与接种前的数量相比,这5种菌完全不适合松材线虫的生长繁殖。当利用绿色木霉(Tv)培养松材线虫时,10 d后平板上的松材线虫数量显著高于作为对照的灰葡萄孢平板上的松材线虫数量,说明绿色木霉的菌丝极其适合于松材线虫的生长,在绿色木霉菌落上松材线虫生长快、数量多。松材线虫也能在其他D17、D23、C28和C25等菌种平板上生长,但非常缓慢,生长繁殖显著差于作为对照的灰葡萄孢上培养的松材线虫。由此初步选出茯苓、黄孢原毛平革菌、B18、C11、D16这5个明显不适宜松材线虫生长繁殖的菌株作为进一步研究的菌/菌株。
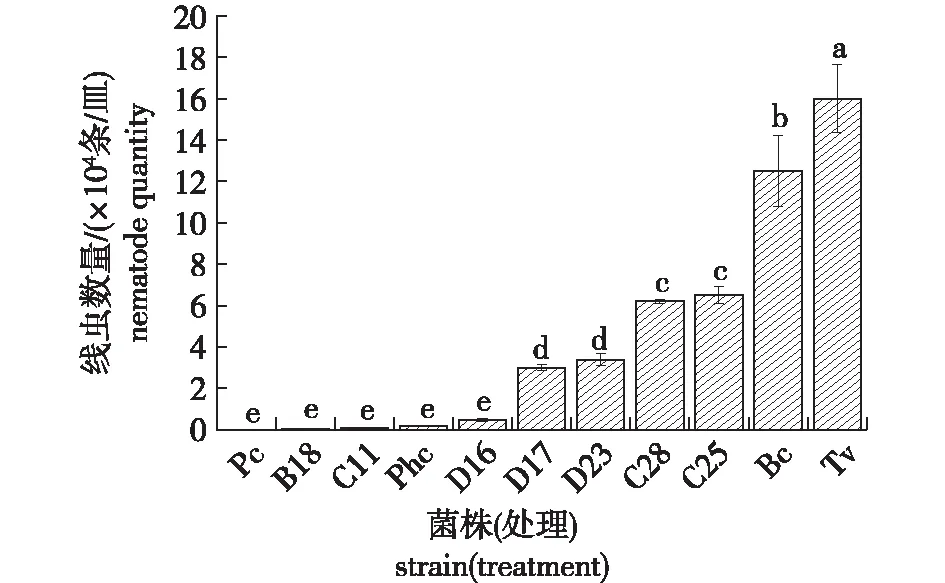
Pc.茯苓Poria cocos;Phc.黄孢原毛平革菌Phanerochaete chrysosporium;Bc.灰葡萄孢菌Botrytis cinerea;Tv.绿色木霉Trichoderma viride。不同小写字母表示处理间差异显著(P<0.05)。下同。Different lower case letters indicate significant difference among all treatments (P<0.05).The same below.
2.2 不同木腐真菌感染对黑松木块质量和木质纤维素的影响
将松木块接入能抑制松材线虫生长的优良木腐菌-茯苓、B18、黄孢原毛平革菌、C11和D16菌落上培养120 d后,去掉木块表面菌丝并把木块烘干至质量恒定后称量,比较黑松木块质量变化(图2)。由图2可知,松木块在5个木腐真菌的菌落上培养120 d后均造成了质量损失,损失率显著高于对照(df = 5,F= 288.957,P<0. 05)。其中茯苓菌对松木块质量的影响最显著,使木块质量损失率达7.5%;其次是B18菌,使木块质量损失率达6%以上。黄孢原毛平革菌和D16菌株对木块质量的影响仅次于B18菌株,C11菌株使木块质量损失率达4%。说明这5个木腐真菌/菌株都可以定殖松木块,降解木块组织使其质量减少。

图2 木腐真菌对黑松木块质量的影响
所用木腐真菌培养至120 d后,烘干木块至质量恒定,粉碎后测定木质纤维素含量的变化,结果见图3。由图3可知,不同的木腐真菌分解木质纤维素能力不一样,相同菌/菌株降解不同的木质组分能力也有差异。茯苓分解纤维素和半纤维素能力最强,分解纤维素19.21%和半纤维素29.65%,而对木质素降解能力仅为1.92%,表明茯苓分解木质素能力较弱。其他的4个木腐真菌/菌株,降解纤维素和半纤维素能力大小依次为B18 > C11 > D16 > 黄孢原毛平革菌,降解木质素能力大小依次为黄孢原毛平革菌 > C11 > D16 > B18。虽然黄孢原毛平革菌分解纤维素能力较弱,但其降解木质素最多,降解率为31.59%,降解能力显著优于其他处理(纤维素:df = 5,F= 136.704,P<0.05。半纤维素:df=5,F=78.751,P<0.05。木质素:df=5,F=63.425,P<0.05)。

图3 木腐真菌对黑松木块木质纤维素含量的影响
2.4 不同方式接种木腐真菌后疫木内松材线虫数量的变化
接种不利于松材线虫生长繁殖且具有降解松木块能力的木腐真菌液体培养菌丝和固体培养菌丝150 d后,取样分离疫木内松材线虫数量,接种前后取样的误差均在5%以内,比较接种前后疫木内线虫数量的变化(图4)。由图4可知,接种5个木腐真菌后,两种不同培养方式的菌丝均可使疫木内松材线虫数量显著减少(接种液体培养菌丝:df=5,F=1 588.329,P<0. 05。接种固体培养菌丝:df = 5,F=752.951,P<0. 05),同种木腐真菌以接种液体培养菌丝效果优于接种固体培养菌丝的。这5个木腐真菌中,不论是液体培养菌丝还是固体培养菌丝接种,茯苓效果最显著,线虫数量减少率均达65%以上,其次效果较好的为黄孢原毛平革菌和B18菌株,均可使线虫数量减少55% 以上。C11亦能使松材线虫数量显著减少,但减少率较小;而D16菌株固体培养菌丝处理后线虫数量减少率与对照无显著差异,说明此菌接种疫木后对其中线虫的数量变化影响不大。

图4 接种两种培养方式木腐菌丝150 d后松材线虫的数量
2.5 接种液体培养木腐真菌菌丝后天牛蛀道松材线虫数量的变化
将液体培养菌丝接种于疫木150 d后,用斧头将疫木劈开,收集天牛蛀道周围的木样,不同天牛蛀道周围的木样内松材线虫数量见图5。由图5可知,不同木腐真菌处理疫木后,天牛蛀道周围的木材线虫数量减少率各异。接种茯苓菌后可使天牛蛀道周围的木材中松材线虫数量减少最多,减少率达65.45%。其次接种效果较好的为B18和黄孢原毛平革菌,可分别减少疫木内松材线虫数量55% 和57%。接种C11菌株的可减少47%,接种D16菌株的减少16%(对照在同期减少12%)。由此可知,接种茯苓、黄孢原毛平革菌、B18和C11后可减少木材中天牛蛀道周围的松材线虫数量。
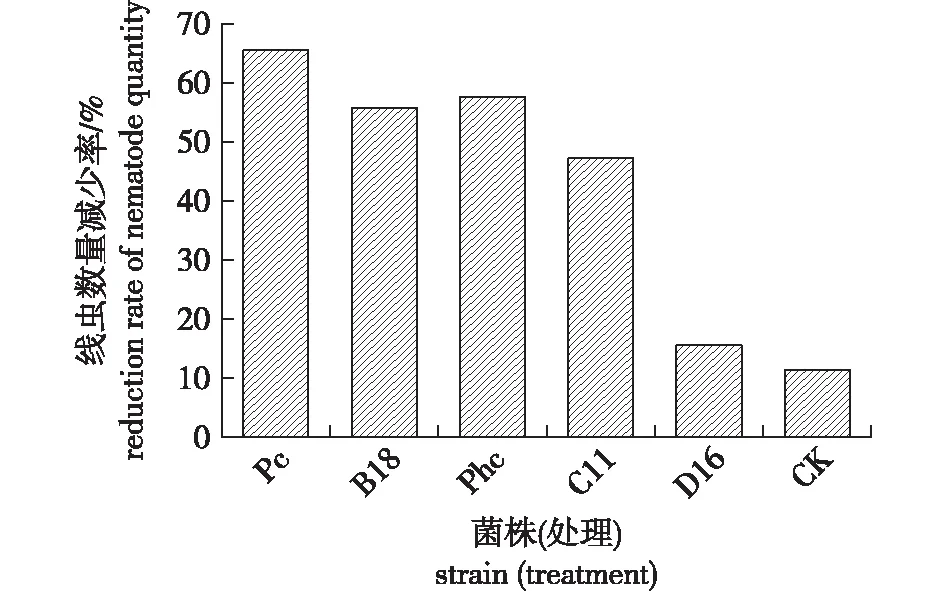
图5 接种液体培养木腐菌丝150 d后天牛蛀道周围松材线虫数量
3 讨 论
本研究通过将松材线虫接种于培养成熟的木腐菌菌落上,构成松材线虫与木腐菌的微小生态系统,使两者之间形成共生、拮抗、竞争和捕食寄生的关系[19]。研究发现包括茯苓菌在内的5种真菌可不同程度地抑制松材线虫的生长繁殖,其中茯苓菌的抑制作用最强。陈瑶等[16]研究亦发现,松材线虫也完全不能在茯苓菌落上存活。自然界中亦存在对松材线虫有不良作用的其他菌株,如总状共头霉(Syncephalastrumracemosum)[20]、日本亮耳(Lampteromycesjaponicus)[21]可较好抑制松材线虫的活性,少孢节丛孢(Arthrobotrysoligospora)[22]可捕食松材线虫,但是很多研究仅停留在室内实验,有待应用于田间验证。
松材线虫病的疫木除害处理一直是松材线虫病防控技术中的难点和要点,疫木处理的效果直接关系到松材线虫病防治的成效[14-15]。当前疫木的就地除害处理或异地除害处理方法,均存在一定的缺陷,如就地处理存在难度大、成本高等难点,异地处理存在容易传播扩散及除害效果难以管控等难题[23-25]。将木腐菌接种于疫木上,木腐真菌通过侵染疫木,改变疫木内的微生境从而使松材线虫数量减少。在两种培养接种方式中,以液体培养菌丝接种效果明显优于固体培养菌丝,可能是因为液体培养菌丝接种,可增加疫木接种部位湿度,为木腐菌的生长提供了良好环境,有利于木腐菌的定殖和扩展。接种不同木腐真菌后减除线虫效果不一,其中茯苓菌减除线虫数量效果最佳,这与胡赛蓉等[26]、泽桑梓等[27]、吴云忠[28]研究结果相似。松材线虫由南向北扩散过程中,在低温条件下具有更强的繁殖能力和对低温的适应性[29]。木腐真菌的最适生长温度为25~30 ℃,而疫木的处理主要在天牛羽化前进行,此时温度还比较低,可开展菌株的低温驯化实验,使菌株在低温下仍能快速繁殖,从而达到更好的除治效果。
松材线虫有向天牛蛹室聚集的效应[30]。本研究发现木腐真菌不仅具有消减松材线虫种群数量的作用,还可大大减少松材线虫向蛹室聚集。松墨天牛在羽化后开始传播线虫,且传播扩散时间持续16~30 d后结束[31]。但是接种木腐菌后疫木中的线虫不能向蛹室聚集,使得线虫无法通过媒介昆虫传播到其他健康松树上完成侵染循环,病害的传播扩展就可以得到有效控制。综上分析,应用不利于松材线虫生长和繁殖的木腐真菌进行松材线虫病的疫木除治具有技术可行性和应用前景,值得进一步深入开发研究。未来可通过菌株的低温驯化,构建复合菌,加大接种量或提早接种使木腐菌与疫木作用时间更长,从而达到消减疫木内松材线虫数量的效果。
参考文献(reference):
[1]BO G Z P D, KAZUYOSHI F P D, JACK R S P D, et al. Pine wilt disease[M]. Tokyo: Springer, 2008: 8. DOI: 10.1007/978-4-431-75655-2.
[2]MOTA M M, FUTAI K, VIEIRA P. Pine wilt disease and the pine wood nematode,Bursaphelenchusxylophilus[M]. Tokyo: Springer, 2009: 254. DOI: 10.1007/978-1-4020-9858-1_11.
[3]PROENA A D N, FRANCISCO R, KUBLIK S, et al. The microbiome of endophytic, wood colonizing bacteria from pine trees as affected by pine wilt disease[J]. Sci Rep, 2017, 7(1): 4205. DOI: 10.1038/s41598-017-04141-6.
[4]张锴, 梁军, 严冬辉, 等. 中国松材线虫病研究[J]. 世界林业研究, 2010,23(3): 59-63. ZHANG K, LIANG J, YAN D H, et al. Research advances of pine wood nematode disease in China[J]. World For Res, 2010, 23(3): 59-63. DOI: 10.13348/j.cnki.sjlyyj.2010.03.008.
[5]SHINYA R, MORISAKA H, TAKEUCHI Y, et al. Making headway in understanding pine wilt disease:what do we perceive in the postgenomic era?[J]. J Biosci Bioeng, 2013, 116(1):1-8.DOI: 10.1016/j.jbiosc.2013.01.003.
[6]MOTA M M, VIEIRA P. Pine wilt disease:a worldwide threat to forest ecosystems[M]. Portugal: Springer, 2008: 5. DOI: 10.1007/978-1-4020-8455-3.
[7]JONES J T, HAEGEMAN A, DANCHIN E G, et al. Top 10 plant-parasitic nematodes in molecular plant pathology.[J]. Mol Plant Pathology, 2013, 14(9): 946-961. DOI: 10.1111/mpp.12057.
[8]XUE Q, WU X Q, ZHANG W J, et al. Cathepsin L-like cysteine proteinase genes are associated with the development and pathogenicity of pine wood nematode,Bursaphelenchusxylophilus[J]. Int J Mol Sci, 2019, 20(1):215. DOI: 10.3390/ijms20010215.
[9]LI L L, TAN J J, CHEN F M.Bacilluspumilusstrain LYMC-3 shows nematicidal activity againstBursaphelenchusxylophilusvia the production of a guanidine compound[J]. Biocontrol Sci and Technol, 2018, 28(12): 1128-1139. DOI: 10.1080/09583157.2018.1514587.
[10]LI Y X, WANG Y, LIU Z Y, et al. Functional analysis of the venom allergen-like protein gene from pine wood nematodeBursaphelenchusxylophilususing a baculovirus expression system[J].Physiol Mol Plant Pathol,2016,93:58-66.DOI:10.1016/j.pmpp.2015.12.006.
[11]丁晓磊. 基于高通量测序的松材线虫致病机理与毒力差异研究[D]. 南京: 南京林业大学, 2016. DING X L. The pathogenic mechanism and virulence variations analyses ofBursaphelenchusxylophilusbased on high-throughput sequencing[D]. Nanjing: Nanjing Forestry University, 2016.
[12]LIU K C, ZENG F L, BEN A L, et al. Pathogenicity and repulsion for toxin-producing bacteria of dominant bacteria on the surface of American Pine Wood Nematodes[J]. J Phytopathol, 2017, 165(9): 580-588. DOI: 10.1111/jph.12595.
[13]王曦茁, 曹业凡, 汪来发, 等. 松材线虫病发生及防控现状[J]. 环境昆虫学报, 2018, 40(2): 256-267. WANG X Z, CAO Y F, WANG L F, et al. Current status of pine wilt disease and its control status[J]. J Environ Entomol, 2018, 40(2): 256-267. DOI: 10.3969/j.issn.1000-2006.201809039.
[14]沈彩周, 何龙喜. 江西省松材线虫病的防控思路和措施[J]. 林业科技开发, 2014, 28(6): 6-9. SHEN C Z, HE L X. Prevention and control of pine wood nematode in Jiangxi Province[J]. China For Sci Technol, 2014, 28(6): 6-9. DOI: 10.13360/j.issn.1000-8101.2014.06.002.
[15]郑礼平. 松材线虫病疫木林间就地除害技术[J]. 现代农业科技, 2017(2): 106-107. ZHENG L P. Local pest control technology between pine wood nematodes[J]. Mod Agric Sci Technol, 2017(2): 106-107. DOI: 10.3969/j.issn.1007-5739.2017.02.068.
[16]陈瑶, 汪来发, 朴春根, 等. 处理松材线虫病病死松树伐桩木腐菌的筛选[J]. 林业科学研究, 2008,21(4): 548-554. CHEN Y, WANG L F, PIAO C G, et al. Screening of wood-rotting fungi to treat stumps of dead pine trees caused by PWN (Bursaphelenchusxylophilus)[J]. For Res, 2008, 21(4): 548-554. DOI: 10.13275/j.cnki.lykxyj.2008.04.022.
[17]邓习金, 刘晖, 罗惠文, 等. 应用木腐菌处理松材线虫病疫木伐桩的研究[J]. 科技视界, 2014(26): 341-342, 343. DENG X J, LIU H, LUO H W, et al. Study on using wood-rotting fungi to treat stumps of dead wood caused by PWN (Bursaphelenchusxylophilus)[J]. Sci Technol Vision, 2014(26): 341-342, 343. DOI: 10.19694/j.cnki.issn2095-2457.2014.26.264.
[18]吴婧瑜. 木腐菌降解废弃木质材料的研究[D]. 广州: 华南农业大学, 2016. WU J Y.Research on degradation of waste wood material by three kinds of wood-rot fungus[D]. Guangzhou: South China Agricultural University, 2016.
[19]SAGÜÉS M F, PURSLOW P, FEMNDEZ S, et al. Nematophagous fungi used for the biological control of gastrointestinal nematodes in livestock and administration routes[J]. Revista Iberoamericana De Micologia, 2011, 28(4): 143. DOI: 10.1016/j.riam.2011.06.009.
[20]孙建华. 土壤真菌培养物对松材线虫生长和繁殖抑制作用的研究[J]. 南开大学学报(自然科学版),1997, 30(3): 82-88. SUN J H. Studies on inhibitory effect of soil isolated fungi cultures to growth and propagation of pine wood nematode[J]. J Nankai Univ, 1997, 30(3): 82-88. DOI: 10.3969/j.issn.2095-2457.2014.26.265.
[21]董锦艳, 莫明和, 陈江虹, 等. 亮耳菌杀线虫活性的稳定性[J]. 云南大学学报(自然科学版), 2000,22(5): 365-368. DONG J Y, MO M H, CHEN J H, et al. The morbidity stability of differentLampteromycesjaponicusstrains[J]. J Yunnan Univ(Nat Sci), 2000, 22(5): 365-368. DOI: 10.3321/j.issn: 0258-7971.2000.05.012.
[22]鄢小宁, 郑服丛, 伍素娟. 松材线虫捕食真菌: 少孢节丛孢HNQ11菌株初步研究[J]. 热带作物学报, 2006, 27(2): 90-94. YAN X N, ZHENG F C, WU S J. Preliminary study of strain HNQ11, a pinewood nematode trapping fungus[J]. Chin J Trop Crop, 2006, 27(2): 90-94. DOI: 10.3969/j.issn.1000-2561.2006.02.019.
[23]叶建仁.松材线虫病在中国的流行现状、防治技术与对策分析[J]. 林业科学, 2019, 55(9): 1-10. YE J R. Epidemic status of pine wilt disease in China and its prevention and control techniques and counter measures[J]. Sci Silvae Sin, 2019, 55(9): 1-10. DOI: 10.11707/j.1001-7488.20190901.
[24]闫闯, 宋崇康, 罗致迪, 等. 松材线虫病疫木除害技术综述[J]. 安徽农业科学, 2017, 45(19): 152-154. YAN C, SONG C K, LUO Z D, et al. Overview of techniques of extinction treatment in diseased wood caused byBursaphelenchusxylophilus[J]. J Anhui Agric Sci, 2017, 45(19): 152-154. DOI: 10.3969/j.issn.0517-6611.2017.19.052.
[25]周爱东,徐小明,王岚,等. 松材线虫病发生34 a的综合防控——以江苏省镇江市为例[J]. 江苏林业科技,2019,46(4):54-57. ZHOU A D,XU X M,WANG L,et al. Integrated control of pine wilt disease in the past thirty years in Zhenjiang City of Jiangsu Province [J].Journal of Jiangsu Forestry Science & Technology,2019,46(4):54-57. DOI: 10.3969/j.issn.1001-7380.2019.04.011.
[26]胡赛蓉, 赵宇翔, 李北屏, 等. 松材线虫病疫木安全利用新途径[J]. 中国森林病虫, 2006,25(5): 26-28. HU S R, ZHAO Y X, LI B P, et al. New method of safe application of pine wood nematode infected wood[J]. For Pest Dis, 2006, 25(5): 26-28. DOI: 10.3969/j.issn.1671-0886.2006.05.009.
[27]泽桑梓, 刘宏屏, 季梅, 等. 在思茅松松材线虫病疫木上栽培茯苓的技术研究[J]. 湖南农业科学, 2010(17): 91-94. ZE S Z, LIU H P, JI M, et al. Cultivation techniques ofporia on pine wood nematode infected Simao pine[J]. Hunan Agric Sci, 2010(17): 91-94. DOI: 10.3969/j.issn.1006-060X.2010.17.031.
[28]吴云忠. 松材线虫病疫木种植茯苓试验[J]. 福建林业科技, 2013, 40(4): 51-55. WU Y Z. A preliminary study on cultivation ofPoriacocoson nematode affected pine wood[J]. J Fujian For Sci Technol, 2013, 40(4): 51-55. DOI: 10.3969/j.issn.1002-7351.2013.04.11.
[29]盛若成, 李敏, 陈军, 等. 两株我国南北松材线虫虫株形态指标与致病力比较[J]. 南京林业大学学报(自然科学版), 2019, 43(6): 18-24. SHENG R C, LI M, CHEN J, et al. Comparison of morphological index and pathogenicity of two isolates ofBursaphelenchusxylophilusin southern and northern in China[J]. J Nanjing For Univ(Nat Sci Ed), 2019, 43(6): 18-24. DOI: 10.3969 /j.issn.1000-2006.201907023.
[30]杨振德, 赵博光, 郭建. 松材线虫行为学研究进展[J]. 南京林业大学学报(自然科学版), 2003, 27(1): 87-92. YANG Z D, ZHAO B G, GUO J. Review on behavior studies of the pine wood nematode[J]. J Nanjing For Univ(Nat Sci Ed), 2003, 27(1): 87-92. DOI: 10.3969/j.issn.1000-2006.2003.01.020.
[31]王洋, 陈军, 陈凤毛, 等. 松墨天牛取食期间传播松材线虫的特性[J]. 南京林业大学学报(自然科学版), 2019, 43(6): 1-10. WANG Y, CHEN J, CHEN F M, et al. Transmission ofBursaphelenchusxylophilus(Nematoda: Aphelenchoididae) through feeding activity ofMonochamusalternatus(Coleoptera: Cerambycidae)[J]. J Nanjing For Univ(Nat Sci Ed), 2019, 43(6): 1-10. DOI: 10.3969 /j.issn.1000-006.201903001.
猜你喜欢
科教创新与实践(2022年3期)2022-04-20
绿色天府(2021年1期)2021-12-03
绿色天府(2020年3期)2020-12-13
农民致富之友(2020年23期)2020-09-02
绿色中国(2019年13期)2019-11-26
小学生学习指导(低年级)(2019年9期)2019-09-25
现代园艺(2017年21期)2018-01-03
现代园艺(2017年21期)2018-01-03
数学小灵通(1-2年级)(2017年3期)2017-04-16
生物灾害科学(2015年2期)2015-03-11
