
遮光处理对草珊瑚光合特性及叶片解剖结构的影响
2021-07-28 02:13聂小英朱春晓郑东升陈阳峰李自强肖深根
西北植物学报 2021年6期
聂小英,朱春晓,郑东升,陈阳峰,李自强,肖深根*
(1 湖南农业大学 园艺学院, 长沙 410128;2 湖南天地恒一现代中药产业有限公司, 湖南衡阳 421256)
草珊瑚(Sarcandraglabra(Thunb.)Nakai)属金粟兰科草珊瑚属多年生常绿亚灌木,喜阴、忌强光,是林药间套作优选中药材植物之一,林下郁闭度是影响植物生长发育的重要环境因素。研究表明,郁闭度为0.5~0.7的林分可种植喜阴性中药材[1]。郁闭度的大小直接影响草珊瑚的生长及药材的品质。草珊瑚适宜在郁闭度为0.5~0.7的环境生长,在郁闭度低于0.4、高于0.9时则生长不良[2-4]。魏家鸿等[5]报道荫蔽桐林下间套作草珊瑚综合效益可观,且土壤水分涵养能力在6种模式中最强;杨城等[6]发现60%遮光下的草珊瑚生物量积累最佳;程龙军等的研究表明草珊瑚具有良好的耐阴性,适当遮光处理可以减缓午休现象,提高其光合效率[7]。
草珊瑚喜阴,是林下套作重要的中药材植物之一。目前对草珊瑚耐阴特性和林下套作研究大多以光合生理指标而确定其对光照的需求,存在一定的局限性;且前人对草珊瑚的研究多基于单一指标而进行试验,试验结果与实际生产存在一定的差距。基于此,该研究从光照环境出发,通过人工遮光模拟林下不同郁闭度,测定遮光处理下草珊瑚的光合作用参数,考察草珊瑚的光合作用对光照强度的响应特征,并对各处理叶片解剖结构及叶绿体超微结构进行观测,从分子水平进一步解释其对光照的需求和适应性,更加全面、系统地了解真实生长环境下草珊瑚对光照变化的响应规律,及其叶片解剖结构及植株形态的变化特征,探究草珊瑚耐阴性的强弱以及在湖南地区生长的最适光照条件,进而为草珊瑚人工栽培和林下间套作提供理论参考。
1 材料和方法
1.1 试验材料
试验于2020年2-9月份在湖南省湖南天地恒一现代中药产业有限公司育苗圃进行。选取2年生草珊瑚实生苗按行株距40 cm × 30 cm移栽,缓苗3个月后选取无病虫害、长势基本一致的植株进行相关试验。
1.2 试验设计
于2020年5月在苗圃内采用不同遮阳率遮阳网搭建高约1.5 m的遮阳小棚,设置全光照(L0,对照)以及50%(L50)、70%(L70)、90%(L90)的遮光环境,每个遮光处理间隔1 m。每处理50株,3次重复。期间进行相同田间管理,连续处理120 d后,进行各项指标的测定。
1.3 指标观测
1.3.1 光合作用指标测定取草珊瑚植株顶部往下数第3对叶片,每株取4片,采用Li-6400XT便携式光合作用测定仪测定光合有效辐射(PAR)和光合气体交换参数净光合速率(Pn)、蒸腾系数(Tr)、气孔导度(Gs)和胞间二氧化碳浓度(Ci)[8],并运用Photosynthesis Assistant3.0软件按非直角双曲线修正模型[9]计算光合作用特征参数最大净光合速(Pnmax)、光补偿点(LCP)、光饱和点(LSP)、暗呼吸速率(RD)及表观量子效率(AQE),3次重复。
1.3.2 叶片解剖结构观察摘取顶部往下第3对健康无病虫害叶片,每处理取4片。切取1 cm×1 cm的小块,用70%FAA固定液固定24 h后,采用常规石蜡切片法[10]制片,用Nikon Eclipse E100光学显微镜观察和拍照,每处理取6张切片,每切片取3个视野;同时,采用Image-Pro Plus 6.0软件测定叶片解剖结构指标:叶片厚度、角质层厚度、上表皮厚度、下表皮厚度、栅栏组织厚度、海绵组织厚度,并计算栅海比和组织密实度、组织疏松度。其中,(1)栅海比=栅栏组织厚度/海绵组织厚度;(2)组织密实度=栅栏组织厚度/叶片厚度;(3)组织疏松度=海绵组织厚度/叶片厚度。
1.3.3 叶绿体超微结构观察每处理取顶部往下第3对健康无病虫害叶片,每处理取4片,在叶脉两侧各取1 mm × 1 mm的方形小块。用浓度2.5%戊二醛和1%锇酸进行双固定,固定后用pH 7.4磷酸缓冲液进行漂洗,再用不同浓度乙醇和丙酮溶液依次脱水,然后用不同比例丙酮和包埋剂渗透环氧树脂包埋[11],最后用LeicaUC7超薄切片机切片,于HT7800型透射电子显微镜下观察拍照。
1.4 数据整理与分析
采用Excel进行数据整理与计算,采用SPSS 22.0进行数据方差分析,用Duncan法进行多重比较。
2 结果与分析
2.1 遮光对草珊瑚叶片表面光照强度和光合气体交换参数的影响
2.1.1 叶片表面光合有效辐射日变化特征各遮光处理的草珊瑚叶片表面光合有效辐射(PAR)日变化曲线均呈现出先增后减的单峰型(图1),并均在12:00达到最大峰值;同期各遮光处理的PAR始终表现为L0>L50> L70> L90,且其日均值在各处理组间均差异显著(表1);同时,PAR的日变化幅度在全光照下最大,并随着遮光程度的增加而逐渐减小。可见,各遮光处理达到了较好的调整光照强度的效果。
2.1.2 光合气体交换参数日变化特征草珊瑚光合气体交换参数的日变化曲线在各遮光处理间均相似(图2)。其中,草珊瑚净光合速率(Pn)日变化在各遮光处理下均呈双峰型变化趋势(图2,A),分别在10:00和14:00出现峰值,且在‘午休’时(12:00)出现谷值,在14:00达到最大值,14:00后急剧下降;与L0相比,遮光处理下‘午休’现象得到一定程度的减缓;草珊瑚Pn日均值表现为L70>L50> L90> L0(表1),且各处理组间均差异显著(P<0.05)。草珊瑚叶片的胞间二氧化碳浓度(Ci)日变化在各遮光处理下均呈先降后升再降再升的变化趋势,8:00至10:00呈缓慢下降,10:00后略微上升,中午12:00后逐渐下降, 14:00后快速上升至最大值(图2,B);各遮光处理的Ci表现为L0>L90> L50> L70,且前两者与后两者间差异显著;叶片在14:00前维持较高的Pn,CO2利用率较高,导致其相应的Ci下降(表1)。草珊瑚气孔导度(Gs)和蒸腾速率(Tr)在各遮光处理下均呈单峰型日变化曲线(图2,C、D),并均在全光照下变化幅度最大,且均于12:00达到最大值;Gs和Tr日均值都表现为L0>L50> L70> L90,与相应Pn的表现一致,且L0显著高于各遮光处理(P<0.05),L50和L70处理又显著高于L90处理,但L50和L70处理间差异不显著(表1)。以上结果说明遮光处理显著降低了草珊瑚叶面光合有效辐射和气孔导度、蒸腾速率,减缓了光合‘午休’程度,显著提高了净光合速率,并以遮光70%处理效果较佳。

表1 不同遮光处理下草珊瑚叶片光合气体交换参数日均值
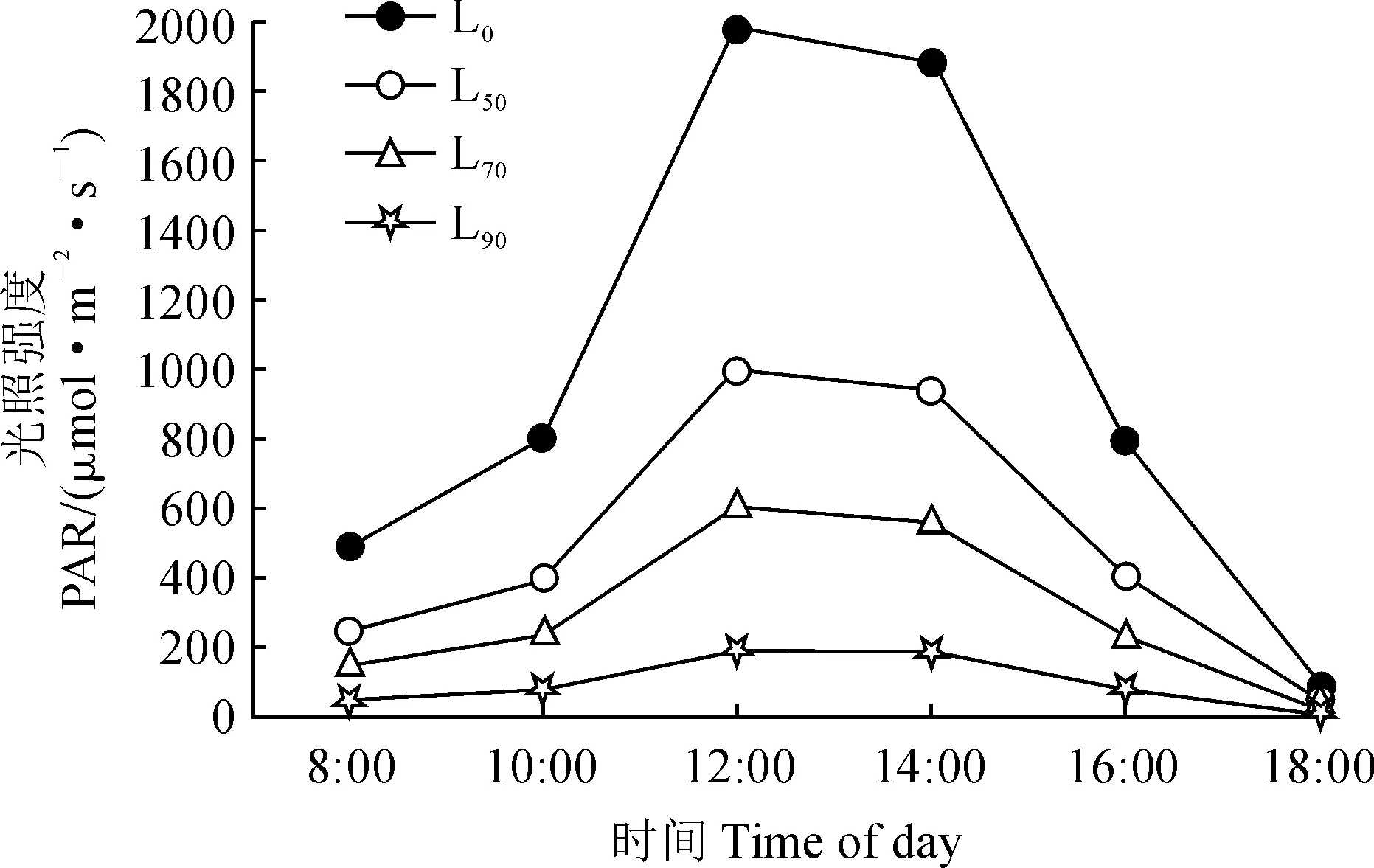
L0、L50、L70、L90分别表示全光照及50%、70%和90%的遮光处理,下同图1 不同遮光处理叶面光合有效辐射日变化L0,L50,L70,L90 stand for 0, 50%, 70% and 90% shading treatments, respectively; the same as belowFig.1 Diurnal variation of photosynthetically active radiation under different shading treatments

图2 不同遮光处理草珊瑚叶片光合气体交换参数日变化Fig.2 Diurnal variation curves of photosynthetic gas exchange parameters in leaves of S. glabra under different shading treatments
2.1.3 光响应曲线各遮光处理草珊瑚光响应曲线(图3)的变化趋势基本一致,随着环境光照强度的增加,净光合速率先迅速增加,达到最大值后稍有下降并基本保持稳定。其中,在各光照强度下,各遮光处理草珊瑚的净光合速率均比全光照处理不同程度提高,并始终表现为L70>L50> L90> L0,且L70和L50处理明显高于L0,而L90处理与L0较接近。

图3 不同遮光处理下草珊瑚光响应曲线Fig.3 Light response curves of S. glabra under different shading treatments
同时,依据光响应曲线计算的光合特征参数(表2)显示,草珊瑚最大净光合速率(Pnmax)在遮光处理下均比L0处理不同程度提高,且L70和L50处理的增幅均达到显著水平,并以L70处理最高且显著高于其他处理(P<0.05);草珊瑚光补偿点(LCP)在各遮光处理下均显著降低,且有随着遮光率的增加而下降的趋势,并以L70和L50处理较大,且显著高于L90处理;光补偿点(LSP)在L50和L70处理下比L0处理显著增加,而在L90处理下比L0处理显著降低,但L50和L70处理间无显著差异;草珊瑚暗呼吸速率(Rd)与LSP的表现相似,在L50和L70处理下显著高于L0处理,在L90处理下稍低于L0处理,而L50与L70处理之间无显著差异;草珊瑚表观量子效率(AQE)在遮光处理下均显著升高,并有随遮光率的增加而逐渐上升的趋势,并以L90处理最大且与其他处理组差异显著(P<0.05)。以上结果说明适当遮光处理降低了草珊瑚叶片的光补偿点,提高了叶片光饱和点和表观量子效率,显著提高了叶片最大净光合速率,并以70%遮光处理效果最佳。

表2 不同遮光处理下草珊瑚光合特征参数
2.2 遮光对草珊瑚叶片解剖结构的影响
2.2.1 叶表皮特征各遮光处理草珊瑚叶片解剖结构和观测结果(图4,表3)显示,随着遮光率的增加,叶片厚度先增后减,且L50和L70处理增幅达到显著水平,L70处理稍有降低;角质层厚度逐渐变薄,且各遮光处理降幅均达到显著水平,但遮光处理间无显著差异;上下表皮均由1层细胞构成,上表皮细胞呈椭圆形或近长方形,排列紧密,下表皮细胞呈不规则形状紧密排列,上下表皮厚度在各遮光处理下均不同程度增加,但仅L70处理增幅达到显著水平。草珊瑚叶片厚度、上下表皮厚度均在L70处理下达到最大值。
2.2.2 叶肉结构特征遮光处理使草珊瑚叶肉组织结构发生明显变化(图4)。其中,在L0处理下,栅栏组织由1层细胞组成,细胞呈椭圆形或长椭圆形,排列整齐,有一定细胞间隙,细胞较短;海绵组织分布疏散、细胞间隙大、呈不规则排列;叶肉细胞内叶绿体数量较少。在L50和L70处理下,栅栏组织均由2层细胞组成,细胞呈椭圆形或近方形,第1层排列整齐,具一定细胞间隙,第2层部分细胞消失;海绵组织呈长椭圆形或椭圆形,不规整排列,在整体叶肉结构中占比最大;两者叶肉细胞中的叶绿体数量明显增加。在L90处理下,栅栏组织由2层细胞组成,但下层部分细胞消失,细胞变短,呈椭圆形,具一定细胞间隙;海绵组织呈不规则疏松排列,细胞间隙大。
同时,表3显示,随着遮光率的增加,各处理栅栏组织和海绵组织的厚度先增加后降低,且均不同程度地高于L0处理;栅栏组织和海绵组织厚度在L50与L70处理间均差异不显著(P>0.05),但两者均显著大于L0和L90处理,L90处理栅栏组织厚度也与L0处理差异显著(P<0.05)。栅栏组织厚度/海绵组织厚度(栅海比)、组织密实度均随着遮光强度增加呈先增后减的变化趋势,并均以L70处理最大,且各遮光处理均显著高于全光照(L0)处理,而各遮光处理间均差异不显著(P>0.05);组织疏松度全光照与各遮光处理之间均差异不显著(P>0.05)。

表3 不同遮光处理下草珊瑚叶片解剖结构指标
2.2.3 叶片叶绿体超微结构在全光照(L0)条件下,草珊瑚叶肉细胞结构完好,未出现质壁分离现象(图5,A)。同时,叶绿体细长,部分叶绿体膜破裂,出现解体现象,叶绿体内含有1~2个淀粉粒,部分淀粉粒出现空洞现象;基粒类囊体变形,片层减少、空隙大,基质类囊体变形严重,排列散乱;在叶绿体内分布少量嗜锇颗粒,线粒体结构完整,呈圆球形,单个或数个聚集分布在叶绿体细胞上(图5,B)。在L50处理下,叶肉细胞和叶绿体结构完整,叶绿体呈椭圆形或纺锤型紧密分布在细胞内壁;少数叶绿体含有淀粉粒,淀粉粒体积小;基粒类囊体片层排列规整,基质类囊体排列紧密,叶绿体上分布有较多的嗜锇颗粒。在L70处理下,叶肉细胞和叶绿体结构完整;叶绿体多呈纺锤型,紧密分布在细胞内壁,叶绿体内出现1~3个淀粉粒,淀粉粒体积较小;基粒类囊体和基质类囊体排列紧密,垛叠规整;在叶绿体内分布少量嗜锇颗粒,线粒体单个或数个聚集分布在叶绿体内。在L4处理下,叶肉细胞结构完整,叶绿体呈长圆形紧密分布在细胞内壁,结构受到轻微破坏;叶绿体内淀粉粒较少,且体积较小;基粒类囊体片层排列规整,部分基质类囊体片层排列松散;在叶绿体上分布少量的嗜锇颗粒(图5,A、B)。

UE.上表皮;LE.下表皮;PT.栅栏组织;ST.海绵组织;Ch.叶绿体图4 不同遮光处理下草珊瑚叶片解剖结构UE. Upper epidermis; LE. Lower epidermis; PT. Palisade tissue; ST. Spongy tissue; Ch. ChloroplastFig.4 Anatomical structure of Sarcandra glabra leaves under different shading treatments
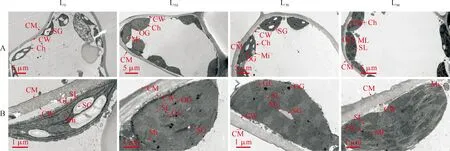
Ch.叶绿体;CM.细胞膜;CW.细胞壁;Mi.线粒体;GL.基粒片层,SL.基质片层;OG.嗜锇颗粒图5 不同遮光处理下草珊瑚叶肉细胞结构(A)和叶绿体超微结构(B)Ch. Chloroplast; CM. Cell membrane; CW. Cell wall; Mi. Mitochondrion; GL. Grana lamellae; SL. Stromal lamellae; OG. Osmiophilic granulesFig.5 Mesophyll cell structure (A) and chloroplast ultrastructure (B) of S. glabra leaves under different shading treatments
2.3 遮光对草珊瑚植株生长的影响
草珊瑚喜阴,忌强光,遮光减小了太阳辐射强度,提高了叶片光合效率,导致其植株生长形态发生显著变化(表4)。其中,草珊瑚株高、地径、叶面积及生物量均随着遮光率的增加而先增后减,且各遮光处理均不同程度地高于全光照处理(L0);各生长指标在L50和L70处理下均显著高于L0和L90处理,并以L70处理最高,其除地径以外的指标也显著高于L50处理。以上说明全光照和重度遮光处理对草珊瑚的生长产生严重的胁迫,适度遮光能显著促进草珊瑚植株的生长。

表4 不同遮光处理草珊瑚生长指标
3 讨 论
光照是影响植物生长发育和进行光合作用最直接的环境因素之一[12-13]。本研究发现,遮光处理对草珊瑚的光合效率有显著影响。全光照和遮光处理的草珊瑚叶片Pn日变化均呈双峰变化,分别于10:00和14:00出现峰值;遮光处理下,草珊瑚叶片具有较低的Ci、Gs、Tr,维持较高的Pn;70%遮光并未对草珊瑚的生长产生抑制作用,但全光照和90%重度遮光对草珊瑚的生长产生明显的抑制作用。从而说明草珊瑚具有一定的光可塑性。同时,不同处理草珊瑚的光响应曲线差异显著。50%和70%遮光处理草珊瑚叶片LCP显著低于全光照处理,而其LSP和AQE则高于全光照处理,同时遮光处理下叶片能维持较高的Pn。可见,全光照下过剩的强光对草珊瑚产生光抑制,而遮光处理显著提高了叶片光合能力,说明草珊瑚具有一定的耐阴性和喜阴性,这与前人的研究结果基本一致[6-7]。
同时,叶片作为进行光合作用的植物器官,外界光照的变化势必会影响叶片形态、表皮、栅栏组织和海绵组织厚度及细胞形态,进而影响其光合特征。研究表明,表皮厚度减小、栅栏组织和海绵组织发达是叶片适应弱光的一种生理特性[14-15];而近方形的栅栏细胞可提高轴面和侧面的叶绿体分布密度,海绵组织厚度的相对增加,排列疏松,有利于提高叶片对光能的利用率[13]。本研究结果表明,随着遮光率的增加,草珊瑚叶片表皮细胞厚度在整个叶肉结构占比减小,栅栏组织和海绵组织发达,栅栏组织细胞长度逐渐变短,排列变得疏松;而海绵组织厚度显著增加,排列疏松;此外,遮光处理下叶肉细胞中叶绿体数量显著高于全光照处理,说明草珊瑚叶片的解剖结构受光照强度的影响较大。
另外,已有研究发现叶片超微结构的变化能极大地影响其光合能力,而叶绿体基粒和基粒片层数目及基粒片层的密集排列有利于增加光合效率[16]。本研究结果表明,在全光照处理下,草珊瑚叶绿体结构出现较严重变形,出现解体现象,叶绿体基粒和基质片层显著减少,垛叠散乱,表明在全光照强度下草珊瑚表现出严重的光抑制,严重影响了叶肉细胞的正常功能;而在50%和70%遮光处理下,草珊瑚叶肉细胞和叶绿体结构完整,基粒类囊体和基质类囊体片层结构发达,叶片捕获光能力增强;但在重度遮光下(90%),叶绿体变细长,基粒类囊体和基质类囊体片层出现松散,叶片光能捕获能力下降,这可能与极端弱光条件下,光合有效辐射较低,叶绿体合成受限有关。
综上所述,草珊瑚在适度(50%和70%)遮光条件下,草珊瑚叶片栅栏组织和海绵组织发达,栅栏组织细胞长度变短,海绵组织厚度显著增加,细胞排列疏松,叶绿体数量显著增加;叶肉细胞和叶绿体结构完整,基粒类囊体和基质类囊体片层结构发达,叶片捕获光能力增强;叶片具有较低的蒸腾速率、气孔导度和胞间二氧化碳浓度,光合‘午休’现象得到有效缓解,具有更高的净光合速率;株高、地径、叶面积及生物量显著增加。草珊瑚具有较强的耐阴性,适度遮光有利于植株生长,全光照或者过度郁闭则会严重抑制生长。本研究从叶片解剖结构、叶绿体超微结构、叶片光合特性和植株生长等方面探究了不同遮光处理对草珊瑚生长的影响,证明50%~70%遮光环境最有利于草珊瑚进行光合作用及光合产物积累和生物量增加,从而显著促进植株生长。因此,在人工栽培时可适当进行遮光处理,在林下间套作时可优选中郁闭度的林分。
猜你喜欢
蔬菜(2022年12期)2022-01-01
星星·散文诗(2021年34期)2021-12-21
作物研究(2020年6期)2020-01-10
农村百事通(2019年1期)2019-01-30
中国林副特产(2018年3期)2018-06-22
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年4期)2016-03-16
绿色科技(2015年8期)2015-04-20
农家顾问(2014年11期)2014-12-01
食品科学(2013年6期)2013-03-11
