
花鲈aqp3a基因的克隆、表达分析及渗透调节功能研究*
2021-07-27 14:29:38李金库温海深孙冬磊王灵钰
中国海洋大学学报(自然科学版) 2021年9期
李金库, 于 朋, 温海深, 齐 鑫, 孙冬磊, 王灵钰, 李 昀
(海水养殖教育部重点实验室(中国海洋大学), 山东 青岛 266003)
硬骨鱼类与陆生脊椎动物一样都需要维持体液的离子平衡及渗透稳态,鱼类在水中面临更加多变的离子环境,因而维持体液稳态对硬骨鱼类来说更具有挑战性[1]。为了应对水环境中的盐度变化,鱼类进化出复杂的渗透调节机制[2]。长久以来,人们普遍认为水分的跨膜运输是以自由扩散的方式通过磷脂双分子层。然而,在之后的研究中人们发现水分子可以快速的通过选择性通道进入红细胞而其它的溶质分子和离子则不能通过。直至1991 年,Agre 等完成了对水通道蛋白1(AQP1)的克隆和鉴定[3],从而确定了细胞膜上存在转运水的特定性通道蛋白,由此揭开了研究水通道蛋白的序幕。随后人们在脊椎动物中共鉴定出17个水通道蛋白同源基因(aqp0~aqp16),将其称为水通道蛋白家族,其功能多样,能显著增加细胞膜水的通透性,参与水的分泌、吸收及细胞内外平衡的调节,广泛参与细胞渗透压以及生理与病理的调节。
水通道蛋白3(AQP3)参与水的分泌、吸收及细胞内外平衡的调节,在鱼类的渗透调节过程中发挥重要作用。AQP3作为水通道蛋白家族的一员,除了对水有通透性以外,还允许甘油和尿素等小分子物质的通过。近年来有关硬骨鱼中aqp3基因的研究取得了一些进展。 Cutler等在欧洲鳗鲡(Anguillaanguilla)中克隆出aqp3基因并对其组织表达谱进行了分析,发现其主要表达于眼、食道、肠和鳃,在胃和肾脏中没有表达或表达量很低。对其鳃中aqp3的表达量进行分析,结果显示其表达水平在海水适应过程中显著降低[4]。进一步研究表明aqp3与水或尿素的运输有关,AQP3可能在基底外侧膜中充当一个管道,将水释放到浆膜液中,并防止细胞肿胀[4-6]。随后,Hirata 等在瓦氏雅罗鱼(Tribolodonhakonensis)中也鉴定出aqp3基因。研究发现酸性环境中瓦氏雅罗鱼鳃组织中aqp3的表达量显著增加,并利用免疫组化技术将AQP3定位在了瓦氏雅罗鱼鳃组织的氯细胞中,与Na+/K+-ATP酶的定位结果相似。这表明酸诱导的aqp3表达促进了基底外侧膜的水转运,在一定程度参与了渗透调节[7]。Watanabe等在莫桑比克罗非鱼(Oreochromismossambicus)中克隆出了aqp3基因,利用RT-PCR对海水和淡水适应过程中的aqp3基因进行组织表达分析,结果显示在淡水适应过程中,AQP3在主要的渗透调节器官如鳃中的表达量明显高于海水,而在肾、肠中的表达情况相反。此外利用免疫组化技术将aqp3定位在了鳃中氯细胞的基底外侧[8]。随着多种硬骨鱼类全基因组数据的公布,越来越多的aqp3在硬骨鱼中被鉴定出来,例如日本鳗鲡(Anguillajaponica)[9]、日本青鳉(Oryziaslatipes)[10]、欧洲鲈(Dicentrarchuslabrax)[11]、萨罗罗非鱼(Sarotherodonmelanothern)[12]等,使得硬骨鱼aqp3的结构、功能方面的研究愈发深入。
花鲈(Lateolabraxmaculatus)又称中国花鲈,俗称海鲈、七星鲈、寨花等,广泛分布于中国沿海以及朝鲜沿海,是中国重要的海产经济鱼类[13]。花鲈具有广盐性的特征(能适应0~45的盐度范围)[14],是研究渗透调节机制的理想模型,然而关于花鲈水通道蛋白基因的研究未见报道。本研究获得了花鲈aqp3a基因的cDNA全长序列,并对其序列特征进行了系统分析;利用枝位点模型进行了选择压力分析,预测出了aqp3a基因进化过程中受到正选择的位点;利用qRT-PCR技术检测了花鲈aqp3a基因的组织表达谱,及其在淡水和海水转换后的各个时间点(0h、12h、1d、3d、7d)鳃中的表达情况。此外,利用显微注射技术将花鲈aqp3a在非洲爪蟾的卵母细胞中进行异源表达,以进一步检测AQP3a蛋白的透水活性。本研究结果为深入研究花鲈aqp3a基因的渗透调节功能提供了基础,同时也为其它鱼类水通道蛋白的相关研究提供了参考。
1 材料与方法
1.1 盐度转换实验
实验所用花鲈及实验场地均由东营市利津县双瀛水产苗种有限责任公司提供。随机选取90尾室内海水(盐度30)人工养殖的健康的1龄花鲈((120.66 ± 1.87) g),随机分配到6个体积为100 L的白色塑化桶中(3个淡水桶,3个海水桶),每桶放置15尾。于实验开始前3周分别在淡水(FW,盐度0)和海水(SW,盐度30)环境下驯化,驯化期间于每日8:30和16:30投喂配合饲料,并在投喂1 h后吸除残饵和粪便。每2天换一次水,换水量为50%。
驯化3周后正式进行盐度转换实验:将淡水中的个体迅速转入海水,标记为FW-SW组;同时将海水中的个体迅速转移至淡水中,并标记为SW-FW组,每个实验组设置3个重复。其它实验条件包括温度((15.0±0.5 ) ℃)、溶解氧((7.1±0.4) mg/L)、pH(8.0±0.3)等在整个实验过程中保持恒定。分别于盐度转换前(0 h)和转换后的4个时间点(12 h、1 d、3 d 和 7 d)采集样品,于各实验组的3个平行桶中各取3尾鱼,即每个实验组在各个时间点共取9尾鱼,经MS222(200 mg/L)麻醉后立即取样。采集鳃组织样本并迅速放入液氮中,随后置于-80 ℃保存。另外,0 h对照组每条鱼除鳃组织外,采集肾、性腺、胃、肠、肌肉、心、脾、肝和脑9个组织,并立即放入液氮中,随后置于-80 ℃保存,用于总RNA的提取和组织表达谱的分析。
1.2 总RNA的提取与cDNA的合成
用TRIzol试剂(Invitrogen, Carlsbad, CA, USA)从所取组织样品中提取总RNA。采用Biodropsis BD-1000核酸分析仪(OSTC, Beijing)和琼脂糖凝胶电泳检测提取的RNA的浓度和完整性。使用PrimeScriptTM RT Reagent Kit with gDNA Eraser (Perfect Real Time) (RR047A Takara)试剂盒并按照说明书进行cDNA的合成。合成的cDNA置于-20 ℃保存备用。
1.3 花鲈aqp3a基因的鉴定及克隆
从NCBI (http://www.ncbi.nlm.nih.gov/)数据库中检索并下载6种脊椎动物的aqp3核苷酸序列,包括人(Homosapiens)、小鼠(Musmusculus)、鸡(Gallusgallus)、非洲爪蟾(Xenopuslaevis)、尼罗罗非鱼(Oreochromisniloticus)和斑马鱼(Daniorerio),利用TBLASTN (1×e-5)对花鲈参考基因组(PRJNA408177)及全长转录组数据库(PRJNA515783)进行检索,初步确定花鲈候选aqp3a基因序列。
按照SMARTer RACE 5′/3′ Kit (TaKaRa)说明书,以花鲈鳃组织的总RNA为模板合成第一链cDNA。根据获得的候选aqp3a基因序列设计用于CDS克隆的正、反向引物和用于5′、3′非编码区克隆的特异性引物,以花鲈鳃组织cDNA为模板进行PCR扩增。PCR 扩增产物经 1%的琼脂糖凝胶电泳后,用FastPure® Gel DNA Extraction Mini Kit胶回收试剂盒(Vazyme)纯化,纯化产物与TA/Blunt-Zero载体(Vazyme)连接,转化进大肠杆菌DH5α感受态细胞,并筛选阳性克隆测序。实验所用引物信息详见表1, 引物合成及测序均由华大基因完成。将获得的序列利用open rea-ding frame (ORF) finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html)在线软件来预测氨基酸序列,并通过BLASTP比对NCBI非冗余(NR)蛋白序列数据库进行进一步验证。利用CDD数据库(https://www.ncbi.nlm.nih.gov/cdd/)对鉴定得到的花鲈aqp3a基因的特征结构域进行预测。

表1 引物名称及序列Table 1 Primer names and sequences
1.4 氨基酸序列分析
通过ExPASy Prot-Param在线工具(https://web.expasy.org/protparam/)计算得到预测的花鲈AQP3a蛋白的分子量(MW, kD)和等电点(pI)。利用TMHMM Server v2.0 (http://www.cbs.dtu.dk/servi-ces/TMHMM/)预测编码氨基酸序列中的α跨膜螺旋。通过Swiss-Model程序(https://swissmodel.expasy.org/)构建花鲈AQP3a蛋白的三维结构模型,并用PyMOL 1.3软件作图。通过DNAMAN软件对人、小鼠、鸡、非洲爪蟾、欧洲鲈、尼罗罗非鱼、遮目鱼(Chanoschanos)、黄颡鱼(Tachysurusfulvidraco)、银大马哈鱼(Oncorhynchuskisutch)、斑马鱼以及花鲈的预测AQP3a蛋白进行氨基酸多重序列比对,并对序列同源性及亲水性进行分析。
1.5 系统进化分析
利用花鲈和其它几种脊椎动物的AQP3氨基酸序列进行系统进化分析。利用MEGA 7.0软件的Neighbor-Joining(NJ)法构建系统进化树,bootstrap值为1 000。
1.6 选择压力分析
基于花鲈与其它物种构建的系统进化树,采用枝位点模型对两个主要的进化枝(硬骨鱼类与高等脊椎动物)进行选择压力分析。以硬骨鱼类为前景枝,高等脊椎动物为后景枝,利用EasyCodeML程序对枝位点模型的各项参数进行计算,采用似然比检验(LRT)比较模型对的拟合程度,采用贝叶斯法(BEB)对正选择位点进行识别[15],利用Protter (http://wlab.ethz.ch/protter/start/)在线软件绘制AQP3氨基酸的二级结构并标注正选择位点的位置分布。
1.7 qRT-PCR分析
使用StepOne Plus Real-Time PCR system (Applied Biosystems) PCR仪,对花鲈aqp3a进行qRT-PCR相对表达定量分析。20 μL反应总体系包含2 μL cDNA模板,正向和反向引物各0.4 μL,10 μL SYBR®FAST qPCR Master Mix、0.4 μL ROX Dye和6.8 μL ddH2O。反应程序如下:95 ℃预变性5 s;95 ℃变性5 s,58 ℃退火30 s,72 ℃延伸30 s, 共计40个循环。以18s RNA为内参[16]。使用primer 5软件设计基因特异性引物,引物具体信息见表1。采用2-ΔΔCT法进行基因的相对表达量的计算, 采用SPSS 21.0统计软件进行单因素方差分析(one way-ANOVA),当P<0.05时认为差异显著。
1.8 花鲈aqp3a基因的异源表达
实验所用非洲爪蟾由山东大学生命学院动物细胞与发育生物学重点实验室提供。非洲爪蟾的年龄为3~4龄,体长(10.22 ± 3.00) cm。实验开始前将非洲爪蟾暂养在循环水系统中(自来水充分曝气;水温:15 ℃;光照:12 h光照/12 h黑暗),每周投喂3次。实验开始后挑选发育良好的雌性个体,注射50单位的人绒毛膜促性腺激素促进其产卵,以人工挤卵的方式获得Ⅴ~Ⅵ期的卵母细胞,并在解剖镜下剥除卵膜。将花鲈aqp3acDNA经体外转录合成cRNA用于非洲爪蟾卵母细胞的显微注射。显微注射系统(ZGENEBIO, PCO-1500, Taiwan)的注射压力为43 psi,注射时间为10 ms。用于显微注射的卵母细胞分为2组,其中实验组每颗卵母细胞注射含10 ngaqp3acRNA的水溶液50 nL,对照组每颗卵母细胞注射50 nL蒸馏水,之后在改良的Barth溶液(MBS,88 mmol/L; NaCl,1 mmol/L; KCl, 0.7 mmol/L; CaCl2,2.5 mmol/L; NaHCO3,1 mmol/L;MgSO4,5 mmol/L;HEPES,1 000 IU/mL;青霉素,链霉素100 μg/mL;制真菌素125 IU/mL)中孵育48 h(18 ℃,渗透压:200 mOsmol)。随后,将卵母细胞转移至10倍稀释的MBS (20 mOsmol)中,通过显微镜5 min内每30 s获取一张卵母细胞轮廓图像。保存的图像随后使用tpsDig232软件进行测量(http://life.bio.sunysb.edu/morph/),以此来测定卵母细胞的透水率(Pf)。Pf值计算公式为:Pf=V0·[d(V/V0)/dt]/ [S·Vw·(Oin-Oout)],其中V0代表卵母细胞的初始体积;[d(V/V0)dt]表示相对卵母细胞体积随时间的变化;Vw为水的摩尔体积(Vw=18 cm3/mol);S为卵母细胞初始表面积;Oin和Oout为卵母胞内、外环境的渗透压。
2 实验结果
2.1 花鲈aqp3a基因的鉴定及序列分析
克隆获得了花鲈aqp3a基因的cDNA全长序列,其序列的特征信息列于表2:aqp3a基因的mRNA全长2 058 bp,其中5′非编码区长84 bp,3′非编码区长1 065 bp,CDS长909 bp,编码302个氨基酸,编码区含有2个水通道蛋白家族的特征性天冬酰胺-脯氨酸-丙氨酸(NPA)序列(见图1)。预测结果表明其存在6个α跨膜螺旋,跨膜位点为:21~43、58~80、101~123、162~181、194~216和249~271(见图1)。预测的花鲈AQP3a蛋白的三维结构如图2所示。AQP3a的C端和N端氨基酸的亲水系数较高,其亲水性氨基酸位点主要分布在跨膜区域,而其它位点表现出较强的疏水性(见图3)。

(加粗部分为起始密码子(ATG)和终止密码子(TAG);黄色标注序列为6个α跨膜螺旋;红色圆圈内序列为2个水通道蛋白家族的特征性天冬酰胺-脯氨酸-丙氨酸(NPA)序列。 The bold part is the start codon (ATG) and the stop codon (TAG),The yellow labeled sequences were 6 transmembrane helices. The sequence in the red circle is the characteristic Asparagine-Proline -Alanine (NPA) sequence of the Aquaporin family.)

表2 花鲈aqp3a基因序列信息统计Table 2 Summary of characteristics of aqp3a genes in spotted sea bass

(图中TM1~TM6为α跨膜螺旋,NPA为天冬酰胺-脯氨酸-丙氨酸序列。In the figure, TM1~TM6 is the transmembrane helix, and NPA is the sequence of Asparagine-Proline-Alanine.)

(纵轴数值大小代表氨基酸序列的亲水性,值越大亲水性越强。 The value of the vertical axis represents the hydrophilicity of amino acid sequence, the higher the value, the stronger the hydrophilicity.)
2.2 AQP3氨基酸序列的多重比对及同源性分析
利用DANMAN软件对花鲈与其它10种脊椎动物的AQP3的氨基酸序列进行同源比对,结果如图4所示,各个物种间的蛋白序列相对保守,都有2个高度保守的天冬酰胺-脯氨酸-丙氨酸(NPA)模体以及6个α跨膜螺旋。花鲈AQP3a与欧洲鲈AQP3的同源性最高,为91.1%,其次是尼罗罗非鱼AQP3ab,同源性为89.6%,与其它高等脊椎动物的同源性则较低,为68.1%~69.4%。

(图中黄色矩形内的序列代表6个共同的α跨膜螺旋;橙色矩形内的序列代表水通道蛋白家族的特征性天冬酰胺-脯氨酸-丙氨酸(NPA)序列。The sequence in the yellow rectangle represents 6 common transmembrane helices. The orange rectangle represents the characteristic Asparagine-Proline-Alanine (NPA) sequence of the Aquaporin family.)
2.3 AQP3a的系统进化分析
利用MAGA 7.0 软件构建了aqp3基因的系统进化树。如图5所示,高等脊椎动物的aqp3基因聚成一枝,硬骨鱼类的aqp3基因则聚为另外一枝。其中花鲈aqp3a与欧洲鲈aqp3、尼罗罗非鱼aqp3ab的同源关系相对较近。

图5 AQP3的系统进化树Fig. 5 Phylogenetic trees of AQP3
2.4 选择压力分析
我们采用枝位点模型,选取硬骨鱼为前景枝、高等脊椎动物为后景枝,来检验aqp3在硬骨鱼中表现出的正选择位点,并通过似然率检验(LRT)获得P值确定零假设模型和备选模型之间是否存在差异。如表3所示,根据贝叶斯法(BEB),硬骨鱼和高等脊椎动物之间存在22个正选择位点:15 F,16 Q,42 A,46 L,55 M,61 F,67 A,73 V,74 C,92 L,94 G ,99 R*,105 F**,107 F**,119 I,122 M,127 L,164 I,174 I,203 L**,219 L,236 T,其中99 R,105 F,107 F和203 L为显著受正选择的位点(P>0.95),利用Protter (http://wlab.ethz.ch/protter/start/)在线软件根据花鲈AQP3a氨基酸序列绘制其二级结构并标注正选择位点(见图6),其中的3个位点分别在2个跨膜螺旋中,剩下的1个位点位于构成水孔通道的重要位置。相比于高等脊椎动物,在硬骨鱼中,99位的精氨酸替换了异亮氨酸,105位和107位的苯丙氨酸替换了丙氨酸,203位的亮氨酸替换了苏氨酸。

表3 AQP3的选择压力分析Table 3 Selective pressure analysis of AQP3
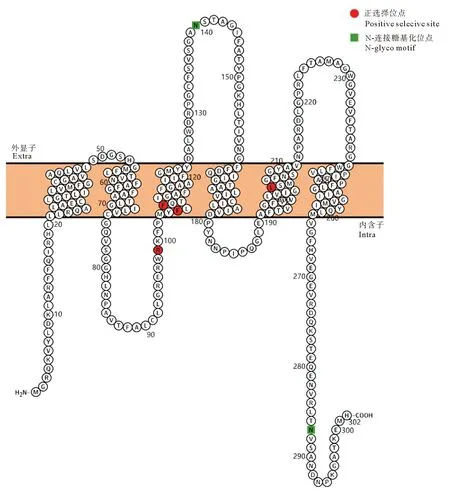
图6 枝位点模型预测的AQP3的正选择位点Fig. 6 Distribution of positive selection sites of AQP3 predicted by the branch-site models
2.5 花鲈aqp3a基因mRNA的组织表达谱分析
花鲈aqp3a基因的组织表达谱分析结果如图7所示,aqp3a基因在各个组织中的表达情况存在显著差异。以表达量最低的心脏组织为参照,计算aqp3a基因在其它各个组织中的相对表达量,结果显示,aqp3a在鳃中的表达量显著高于其它各个组织。aqp3a在鳃组织中的表达量最高,是其在心脏中表达量的416.36倍,而在其它组织中表达量相对较低。

(柱形图上方字母不同表示差异显著(P<0.05)。Different letters on the column indicate significant difference (P<0.05).)
2.6 在淡水及海水适应过程中花鲈aqp3a基因在鳃中的表达响应
在淡水适应(SW-FW组)过程中,花鲈aqp3a基因在鳃中的表达情况如图8A所示,转入淡水后12 h,aqp3a基因的表达量与对照组无明显差异,1 d时表达量显著上调并达到最大值,为0 h的32.6倍,3 d的表达量上调为0 h的18.5倍,7 d时的表达量降低至与0 h无显著差异。在海水适应(FW-SW)过程中,aqp3a基因在鳃中的表达情况如图8B所示,在转入海水后其表达量逐渐下调然后维持稳定,在1 d时显著下调为0 h的0.32倍,3和7 d时aqp3a的表达量维持在较低水平,与1 d时无明显差异。
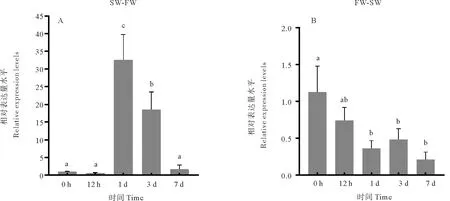
(柱形图上方字母不同表示差异显著(P<0.05)。图A为淡水适应过程中,花鲈aqp3a基因在鳃组织中的相对表达量水平。图B为海水适应过程中花鲈aqp3a基因在鳃组织中的相对表达量水平。Different letters on the column indicate significant difference (P<0.05). Fig. A shows the relative expression level of aqp3a gene in the gill tissue of spotted sea bass during freshwater adaptation. Fig. B shows the relative expression level of aqp3a gene in the gill tissue of spotted sea bass during seawater adaptation.)
2.7 花鲈AQP3a在非洲爪蟾卵母细胞中的功能验证
为了检测花鲈AQP3a的水转运活性,本研究将花鲈AQP3a在非洲爪蟾的卵母细胞中进行了异源表达。结果如图9所示,显微注射了aqp3acRNA的实验组卵母细胞体积在5 min内膨胀至初始体积的1.3~1.4倍,而注射蒸馏水的对照组体积膨胀较小或无显著膨胀(见图9A);表达AQP3a蛋白的卵母细胞的透水率(Pf)约为对照组的5倍(见图9B),由此表明花鲈的AQP3a对水的转运及细胞体积的调节发挥重要作用。

(图A为注射了蒸馏水的对照组卵母细胞和注射了花鲈AQP3a cRNA的实验组卵母细胞在转移至低渗培养液后的相对体积变化。矩形代表对照组,圆形代表实验组。图B为对照组与实验组卵母细胞的透水率对比图。 Fig. A shows the relative volume changes of oocytes in the control group injected with distilled water and those in the experimental group injected with AQP3a cRNA after being transferred to low-osmotic medium. The rectangle represents the control group and the circle represents the experimental group. Fig. B shows the comparison of oocyte permeability between the control group and the experimental group.)
3 讨论
本研究成功从花鲈中鉴定出了aqp3a基因,并通过系统进化树分析确定了命名的正确性。将花鲈AQP3a序列与其它物种进行多重比对,结果显示其具有高度保守性。系统进化分析显示,硬骨鱼类的aqp3基因和高等脊椎动物的aqp3基因各聚为一枝,其中花鲈的AQP3a先后与欧洲鲈的AQP3、尼罗罗非鱼的AQP3ab聚为一枝,这表明它们的同源关系较近。此外,采用枝位点模型对aqp3进行选择压力分析并在硬骨鱼分支上筛选出了4个显著的正选择位点,其中有3个位点在2个跨膜螺旋中,剩下的位点位于构成水孔通道的重要位置。这些受正选择作用的非同义氨基酸替换可能导致硬骨鱼与高等脊椎动物的AQP3蛋白透水性的显著差异,影响水的通透性并影响蛋白质结构的稳定性。这种氨基酸的非同义替换对蛋白功能的影响需要通过氨基酸残基诱变实验进一步验证。
在硬骨鱼中,aqp3基因的表达具有组织特异性。Cutler等的研究证实了aqp3基因广泛分布在鱼类的鳃、肾、肠等多个组织中且分布和表达量会随着鱼的种类及环境的变化而变化[4-6]。甘远迪等对萨罗罗非鱼(Sarotherodonmelanothern)的研究表明,aqp3基因在鳃、肌肉、皮肤等组织中表达量较高,且表达量随盐度的变化而变化[12]。本研究利用qRT-PCR技术对花鲈aqp3a基因的组织表达谱进行检测,得到了相似的结果,结果显示aqp3a基因在各组织中广泛表达(心脏组织除外),且在鳃中的表达量显著高于其它组织。这表明花鲈aqp3a基因在鳃组织中发挥至关重要的作用。
硬骨鱼类复杂的渗透调节机制使其在面对海、淡水环境时,可通过渗透调节器官调节水盐平衡,在整个渗透调节过程中鳃发挥着至关重要的作用[17-19]。在盐度适应过程aqp3基因在鳃中的表达量具有显著差异,如欧洲鲈、欧洲鳗鲡、日本鳗鲡、莫桑比克罗非鱼等相关研究结果表明aqp3mRNA在淡水条件下的表达量均高于海水条件下的表达量。部分学者认为这是由于AQP3的作用可能是通过水分的转运来调节细胞体积。本实验对花鲈aqp3a基因在淡水适应过程中的鳃组织中的表达变化情况进行研究,结果显示转入淡水后其表达量出现显著上调,与之前的研究结果一致,但在3 d后表达出现下调,7 d后表达趋于稳定并与对照组无显著差异,该结果与Giffard-Mena等人对欧洲鲈aqp3在淡水适应过程中的表达变化的研究略有不同,其鳃组织中aqp3在淡水适应一周后的表达量仍比对照组上调8倍。因此,我们提出另一种假设,在花鲈鳃组织中,aqp3a的作用并非通过建立细胞膜通路直接控制水分运输,而是间接参与细胞体积的调节。以往对欧洲鳗鲡、莫桑比克罗非鱼的免疫组化研究结果显示AQP3主要分布在氯细胞中,与Na+/K+-ATP酶共定位于基底侧的微管网中[8],在低渗环境下,水分子通过水通道蛋白由外界水环境进入氯细胞,导致细胞肿胀,进而激活Na+/K+-ATP酶的活性,调节离子浓度,进而维持细胞体积。而短时间内花鲈aqp3a的显著上调可能与氯细胞的凋亡、转化和细胞分化有关。对罗非鱼胚胎表皮细胞的研究表明,氯细胞存在4种类型且在海、淡水环境中以不同的类型存在[20]。当花鲈由海水转入淡水时,海水型氯细胞大量凋亡而淡水型氯细胞的建立和分化促使aqp3amRNA的显著上调,至淡水适应7 d时表达趋于稳定。同样有很多学者对aqp3在海水适应过程中的表达情况进行了研究,Cutler等对欧洲鳗鲡进行海水驯化,3周后aqp3的表达量显著下降[5]。Tse等将日本鳗鲡由FW转入SW中,鳃中aqp3mRNA水平也出现类似的下降[9]。我们同样对花鲈aqp3a基因海水适应过程中在鳃组织中的表达模式进行了研究,结果显示在转入海水后其表达量逐渐下调,并在1 d后维持稳定。Isaia等认为鳗鲡aqp3mRNA表达的下降与向SW转移后鳃的水渗透性下降有关[21]。Lignot等利用免疫组化在盐度适应的欧洲鳗鲡中,将AQP3定位在了鳃的氯细胞、基底上皮细胞及覆盖鳃弓的上皮细胞中,且在SW驯化的欧洲鳗鲡中,这两个非氯细胞位置的染色均显著减少,并由此推测氯细胞中AQP3蛋白水平的降低可能是导致SW中AQP3 mRNA和蛋白表达下调的原因[22-24]。同样在SW驯化后的罗非鱼鳃中AQP3免疫组化染色程度也有所降低[24]。针对花鲈aqp3a在海水驯化过程中的表达情况,根据我们的假设,其在海水适应过程中表达量逐渐降低然后维持稳定一方面原因是由于海、淡水不同类型的氯细胞相互转化,另一方面可能是由于海水环境中除了氯细胞外,上皮细胞等其它类型细胞中aqp3a的表达量降低。由此表明,aqp3a在花鲈的渗透调节过程中发挥着相当重要的作用。
非洲爪蟾的卵母细胞作为功能基因组学中重要的研究工具可在短时间内正确的表达外源基因,翻译合成蛋白质[25]。为进一步验证花鲈aqp3a基因的功能,利用非洲爪蟾Ⅴ~Ⅵ期的卵母细胞对其进行异源表达。通过显微注射技术将花鲈aqp3a的cRNA注射入卵母细胞中,以此检测其透水活性。实验结果显示花鲈aqp3a的cRNA能够在非洲爪蟾卵母细胞的细胞膜中高效表达,并显著提高细胞膜的透水能力,这表明花鲈aqp3a基因对水的转运和细胞体积调节有重要作用。
4 结语
本研究克隆、分析了花鲈的aqp3a基因,并检测了该基因的组织表达情况。结果表明aqp3a基因存在组织表达特异性,且在鳃中的表达量显著高于其它组织。通过短期盐度转换实验检测该基因在海、淡水适应过程中在鳃组织中的表达变化情况,结果显示aqp3a基因在海、淡水适应过程中的变化情况存在显著差异。同时利用非洲爪蟾的卵母细胞对花鲈aqp3a基因进行异源表达以检测其透水活性,结果显示该基因能显著提高细胞膜的透水活性。本实验对花鲈aqp3a基因在渗透调节中发挥的作用进行了初步研究,其具体的分子机制还需要进一步研究。
猜你喜欢
当代水产(2019年4期)2019-05-16 03:05:14
当代水产(2018年8期)2018-11-02 05:30:16
农家之友(2018年12期)2018-03-12 00:50:36
河南畜牧兽医(2017年8期)2017-11-24 03:21:44
上海农业学报(2017年3期)2017-04-10 12:39:26
渔业信息与战略(2016年2期)2016-12-02 09:54:55
河北渔业(2016年4期)2016-07-30 10:31:22
动物营养学报(2015年9期)2016-01-07 11:29:34
中国海洋大学学报(自然科学版)(2014年9期)2014-02-28 12:21:38
当代畜禽养殖业(2014年1期)2014-02-27 07:58:29
