
日本鲺(Argulus japonicus)寄生大口黑鲈的首次报道及分子系统发育研究
2021-07-26 13:48杨浩顾泽茂
华中农业大学学报(自然科学版) 2021年4期
杨浩,顾泽茂
华中农业大学水产学院/湖北省水生动物病害防控工程技术研究中心,武汉 430070
鲺(Argulus)是一类世界分布的甲壳动物寄生虫,可感染多种淡水鱼类,引发鲺病,造成宿主的大量死亡[1-2]。目前,鲺属内已报道的物种有129个[3]。其中,日本鲺(ArgulusjaponicusThiele 1900)是最常见的种类之一,最早由Thiele在日本发现并命名,之后随着渔业贸易的全球化,在世界各流域均有发现和报道[4-5]。日本鲺在我国分布的首次记载是在1939年[6],遗憾的是该研究未对宿主进行说明。之后,王耕南[7]、黄琪琰等[8]分别在显微和亚显微水平上详细描述了日本鲺的形态学特征。2014年,Alsarakibi等[9]调查了广东地区青鱼、鲢、虹鳟和鳜等日本鲺的寄生情况,发现在8种淡水鱼上均存在寄生和感染,且池塘养殖条件下日本鲺的感染率和感染强度显著高于野生环境,提出在水产养殖过程中应尤其重视日本鲺病。
大口黑鲈(Micropterussalmoides),俗称加州鲈,隶属于鲈形目(Perciformes)、太阳鱼科(Centrachidae)、黑鲈属(Micropterus),原产于北美洲的密西西比河流域,20世纪80年代引入我国[10]。近年来,大口黑鲈已成为我国重要的特色淡水养殖品种之一。然而,随着养殖规模的扩大、集约化程度的提高,大口黑鲈病害频发,严重限制了其养殖产业的发展。2020年5月,湖北省武汉市某养殖场的大口黑鲈发生了一种鲺病,并造成大口黑鲈大量死亡。本文采用形态学和分子生物学的方法,研究了鲺的种类和系统发育关系,将其鉴定为日本鲺,这是日本鲺寄生大口黑鲈的首次报道。
1 材料与方法
1.1 病鱼采集与寄生虫分离
2020年5月,湖北省武汉市某养殖场的大口黑鲈发病,取4尾病鱼(全长30~40 cm,体质量750~850 g),肉眼观察并记录病鱼体表症状,吸管吸取少量蒸馏水浸湿病鱼体表,用镊子轻轻夹取体表寄生虫转移至装有蒸馏水的培养皿中,用于后续形态学和分子生物学鉴定。
1.2 寄生虫形态学观察
挑选形态完整的虫体于1.5 mL的离心管中,加入4%多聚甲醛于-4 ℃固定24 h。用镊子将固定好的虫体转移到干净的载玻片上,使用Olympus BX53显微镜和Olympus DP72数码成像系统从低倍镜到高倍镜依次观察并记录虫体的形态特征。
1.3 基因组DNA提取,SSU rDNA扩增、测序及系统发育分析
1)基因组总DNA提取。将本文“1.1”中的虫体收集到1.5 mL的离心管中,滴加95%的乙醇于-4 ℃下固定24 h。取出虫体置于新的1.5 mL的EP中,待乙醇挥发后,用基因组提取试剂盒(康为世纪,北京)提取虫体基因组总DNA,保存于-20 ℃备用。
2)PCR扩增SSU rDNA基因序列。选用SSU rDNA通用引物,上游引物:5′-AACCTGGTTGATCCTGCCAGT-3′,下游引物:5′-TGATCCTTCTGCAGGTTCACCTAC-3′[11],以材料与方法“1.3 1)”中提取的总DNA为模板,进行PCR扩增。50 μL的PCR反应体系包括:2 × EsTaqMasterMix(25 μL)、游引物(2 μL)、下游引物(2 μL)、模板DNA(2 μL)、ddH2O(19 μL)。PCR扩增条件:94 ℃预变性5 min;94 ℃变性45 s,56 ℃退火30 s,72 ℃延伸1 min,35个循环;72 ℃终延伸10 min。
3)基因克隆与测序。对本文“1.3 2)”中PCR产物进行1.5%琼脂糖凝胶电泳,采用琼脂糖凝胶DNA回收试剂盒(Omega,美国)纯化回收PCR产物,将纯化后的PCR产物连接至pMDTM-19T载体中,转化至感受态细胞DH5α,接于含有Amp的LB固体培养基中,37 ℃培养12 h后挑取单克隆于LB液体培养基中震荡培养5 h,经PCR验证后,将阳性菌株送北京奥科鼎盛生物科技有限公司进行测序。
4)序列比对与分析。将测序结果进行Blast比对,下载BLASTn相似序列以及鲺属中已收录的小亚基核糖体序列。经MAFFT(v7.273)[12]软件比对后,使用G-blocks(v0.91b)[13]删除模糊比对片段,利用IQ-TREE (v1.6.12)[14]软件构建ML系统进化树。
2 结果与分析
2.1 患病鱼症状
病鱼沿池塘四周漫游,呈缺氧状态,活力不强,易捕捉,死亡率达30%。肉眼可见鳍条处有出血症状,体表有淡黄色的圆盘状寄生虫吸附或移行(图1)。镜检鳃部无寄生虫,剖检内脏器官无明显病变。

A.患病大口黑鲈的整体图,箭头指示寄生在体表的鲺; B.大口黑鲈体表病灶图,箭头指示病灶周围的鲺; C.寄生大口黑鲈的鲺(箭头指示)。A. Holistic view of the largemouth bass,arrowheads indicated the parasites; B.The focus of the parasitic infection,arrowheads indicated the parasites; C.Holistic view of the fish louse.
2.2 寄生虫形态特征
镜检观察发现虫体背部有马蹄形背甲,长度略大于宽度,两侧叶上有明显的树状色素条,侧叶末端钝圆,伸达第三游泳足的后缘(图2A);腹面观虫体背甲前缘和两侧覆盖有尖端向后的小刺(图2B),由前向后依次为第一触角、第二触角、复眼、吸盘、口器、颚足、游泳足。腹部呈对称的两椭圆状结构,根据腹部中是否含卵或者对称的纺锤状精巢可判断虫体性别(图2A~D)。形态学数据显示,雌性虫体体长1.93±0.46(1.42~2.54)(n=13) mm,背甲大小为(1.17~2.09) mm× (1.00~1.78) mm;雄性虫体体长2.18±0.49(1.57~2.87)(n=11) mm,背甲大小为(1.18~2.08) mm× (1.00~1.96) mm。
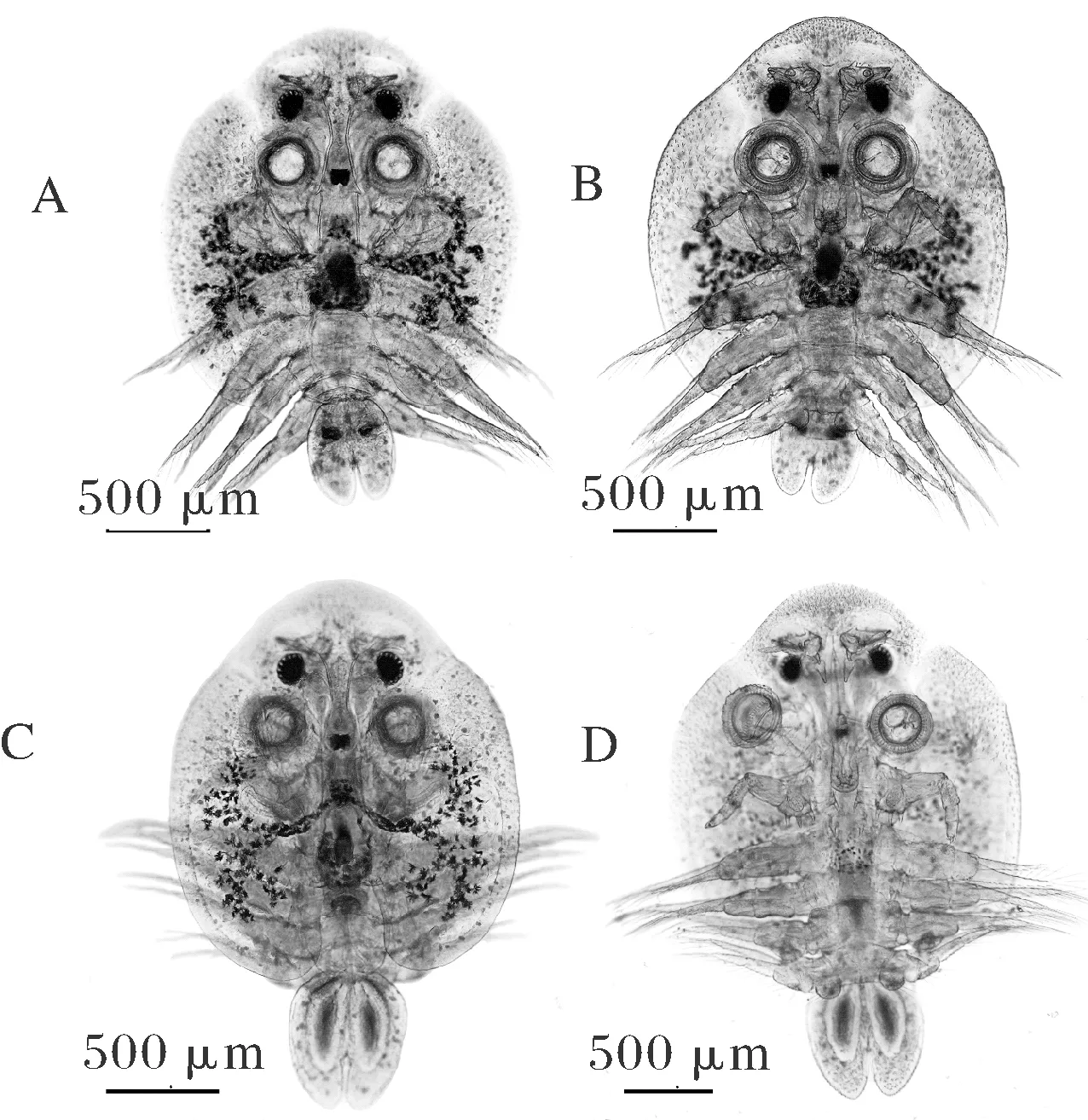
A.雌性背面观; B.雌性腹面观; C.雄性背面观; D.雄性腹面观。A. Dorsal view of the female; B. Ventral view of the female; C. Dorsal view of the male; D. Ventral view of the male.
寄生虫各部位形态描述如下:
第一触角(Fa)的基节由2节组成。第1节和第2节的基部各着生1根刺。第2节的基部外侧有一凸起。第2节末端着生1个尖端向腹面的钩状刺。鞭由2节组成,末端着生5根刚毛(图3A)。
第二触角(Sa)由5节组成。在第1节基部有1根刺,在第1到第4节的末端各着生3~4根刚毛。第5节末端着生5根刚毛(图3B)。
小颚特化为吸盘。雌性吸盘几丁质条数为44~52,每条几丁质条由1枚长基片和4~5枚圆形几丁质片组成。雄性吸盘几丁质条数为46~47,每条几丁质条由1枚长基片和6~7枚圆形几丁质片组成(图3C)。
颚足:共5节,第1节基部有3根粗壮的刺,在第1节的腹面有一椭圆状的突起,凸起的表面分布有尖端向后的小刺,在凸起的后缘着生有中间的3根长刚毛和两侧的5根短刚毛。第2节至第5节长度逐渐减小,在第2节的前端至第4节,表面有分叉的倒刺。在第5节末端,着生有2根倒刺和1个指状凸起(图3D)。
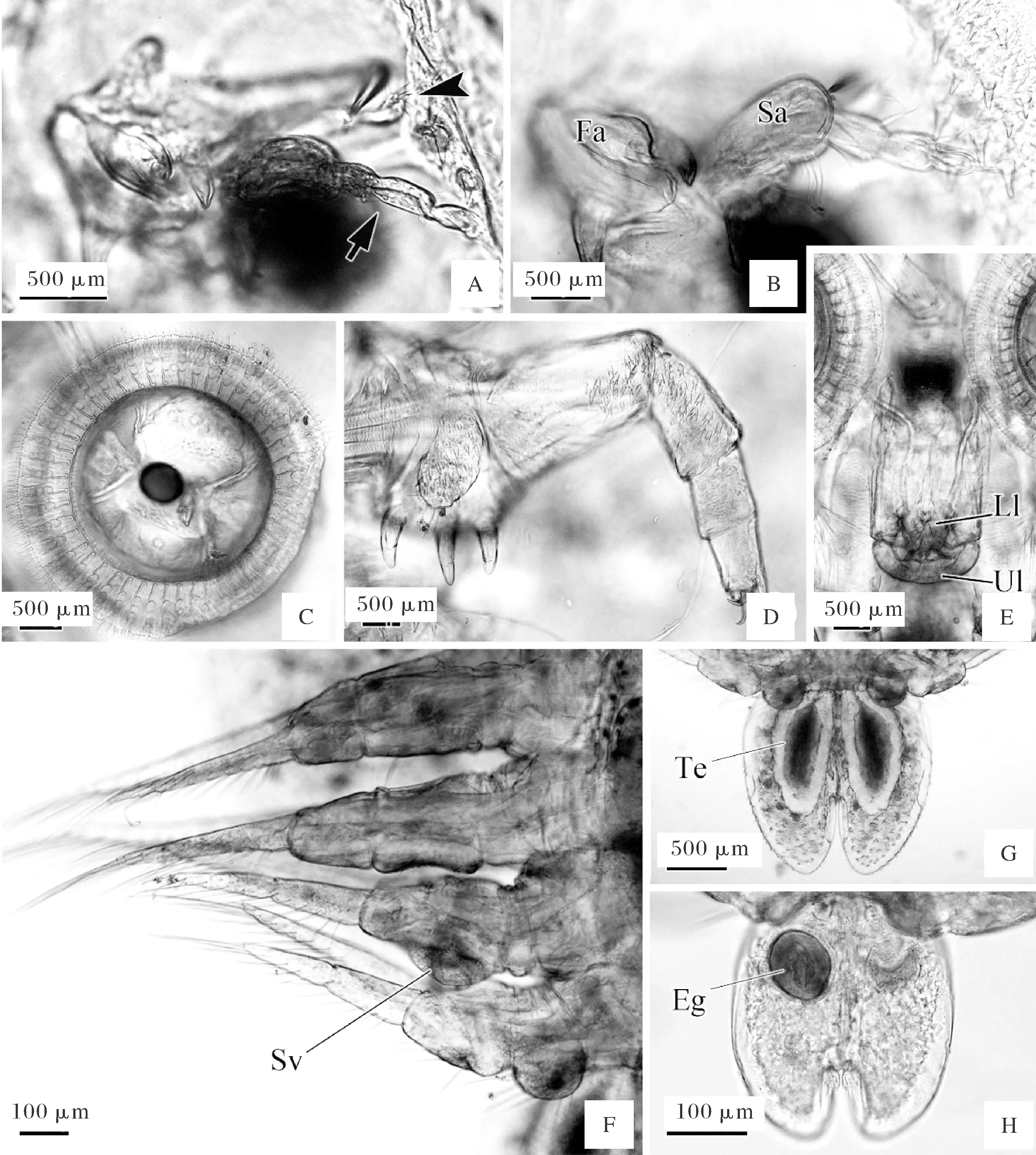
A: 触角,无尾箭头示第一触角鞭,有尾箭头示第二触角; B:第二触角; C:吸盘; D:颚足; E:口管; F:游泳足(雄性); G: 雄性腹部; H:雌性腹部。Eg:卵; Fa:第一触角; Ll:下唇; Sa:第二触角; Sv:精囊; Te:精巢; Ul:上唇。A:Antenna,arrowhead showed the scourge of first antenna,arrow indicated the second antenna; B:Second antenna; C:Sucker,which supportedby sclerites; D:Maxilla; E:Mouth tube; F:Male swimming legs; G:Male abdomen; H:Female abdomen. Eg:Egg; Fa:First antenna; Ll:Lower lip; Sa:Second antenna; Sv:Seminal vesicle; Te:Testis; Ul:Upper lip.
口管呈短圆筒形,位于两吸盘之间。口管由上、下唇组成,上唇(Ul)向腹面呈半圆形,下唇(Ll)形状不规则,在两侧及中间有凸起(图3E)。
游泳足4对。每对游泳足可分为4节,且表面分布有倒刺。每对游泳足在第4节出现2个分支,分支上着生有8根刚毛。在第1对游泳足第2节的内侧,着生有1根刚毛(图3F)。雄性第2游泳足基节腹面有基节突,第三游泳足的内侧有一直径约为200 μm的隆起,为精囊(Sv)。雌、雄虫体在第四游泳足的基节内侧均有一乳头状凸起,在凸起以及第3节上均着生许多刚毛(图3F)。
腹部长度与全长的比值为0.21~0.29,长度略大于宽度(长0.36~0.83 mm,宽0.31~0.65 mm)。中央裂深度为腹长的1/2~1/4。尾叉基位,其上着生4根刚毛。雄性腹部中可见两对称的卵圆形的精巢(T)(图3 G),雌性成虫腹部中可见未孵化的卵(E)(图3H)。
本研究采集到的物种与鲺属内其他物种的形态测量学指标比较见表1。
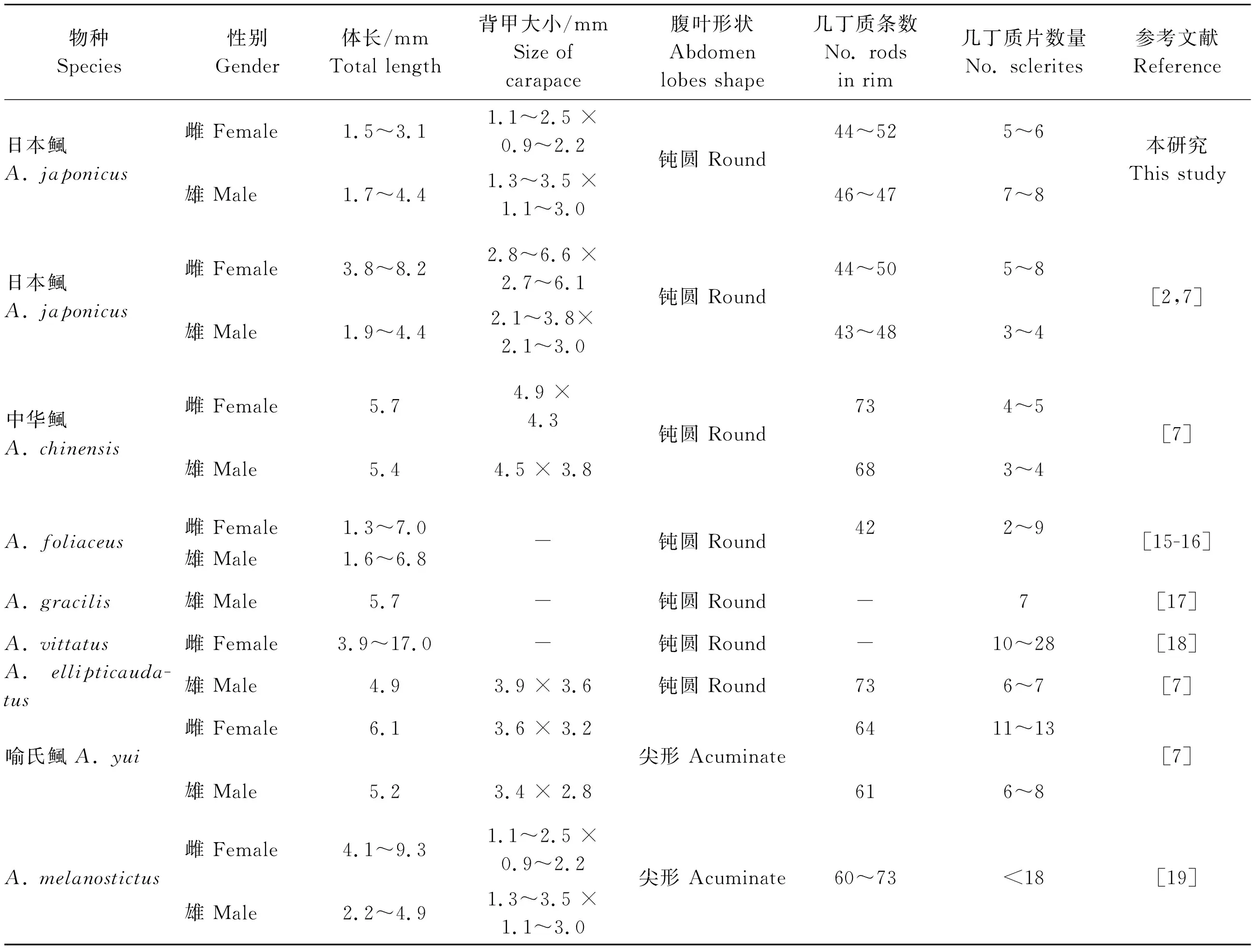
表1 本研究物种与鲺属其他物种的形态测量学数据 Table 1 Measurements of Argulus sp. in this study with other species of Argulus
2.3 系统发育分析
获得SSU rDNA基因片段长度1 800 bp,经BLAST比对发现,本研究中日本鲺株系(MW866569)与Arguluscoregoni(JQ740820)、Argulusfoliaceus(JQ740819)、Argulussp. (JN558648)的相似度均为99.84%,与日本鲺(JN558647)和Argulusnobilis(MH458748)的相似度为99.77%,与鲺属内其他物种的相似度在94.00%~99.70%。基于SSU rDNA序列构建ML系统进化树,结果显示,鲺属内物种存在3个支系(Clade),除JN558647株系外,日本鲺已报道株系均聚为一支(CladeⅠ),本研究采集的日本鲺(Argulussp.)位于CladeⅠ基部,与日本鲺KF747859和KF747860株系亲缘关系最近(图4)。同时,A.foliaceus(KF747861)和A.rhipidiophorus(747862)也聚于CladeⅠ中,与日本鲺在进化树中呈并系关系。
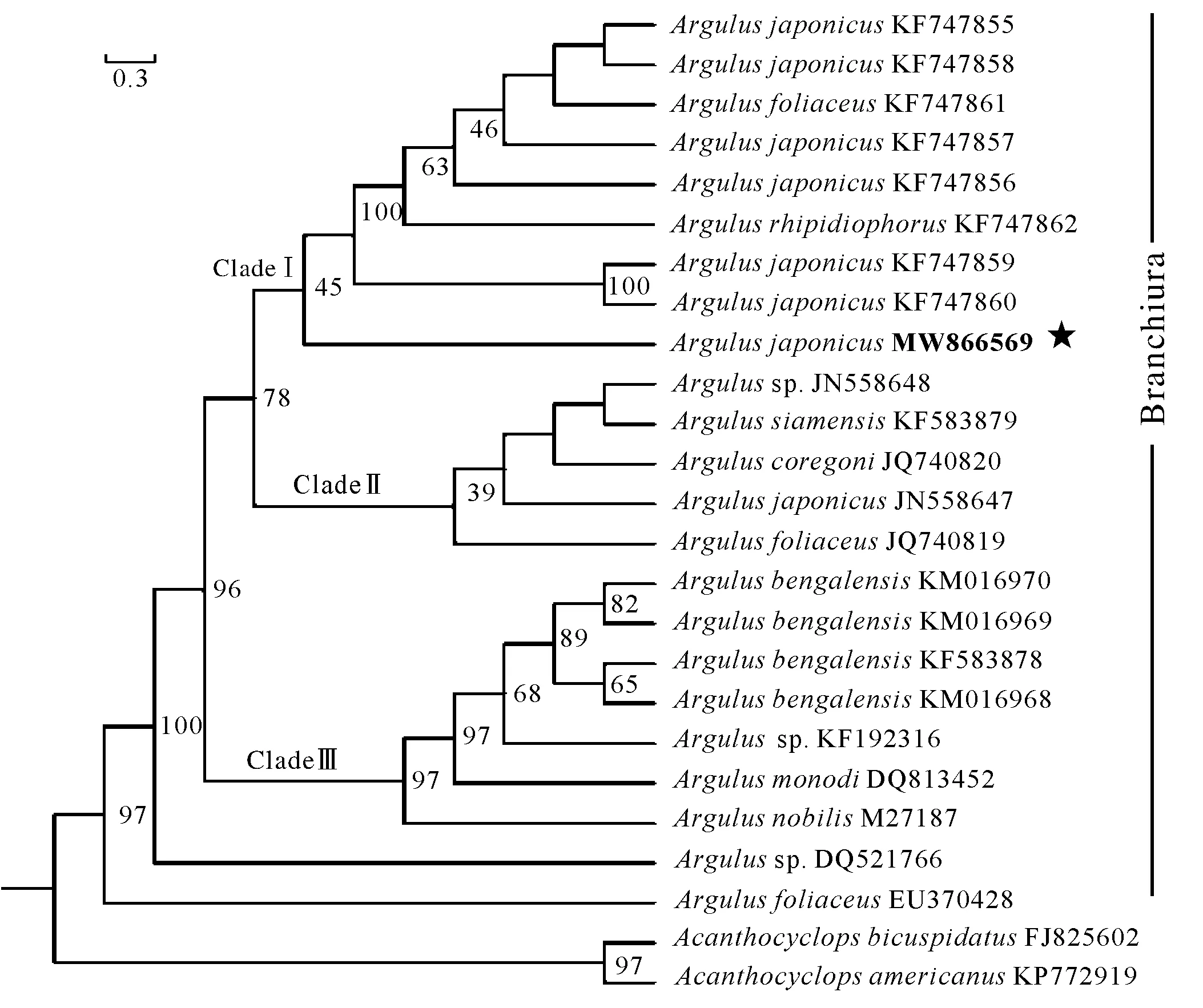
图4 基于SSU rDNA序列构建的ML系统进化树(本研究物种标星号表示)
3 讨 论
日本鲺引起的鲺病常导致多种水产动物的大量死亡,给水产养殖业造成严重的经济损失[21-23]。大口黑鲈是我国重要的淡水养殖品种,然而,在其寄生虫病的研究中,鲜有鲺病的相关报道。Bauer[24]、Angelina等[25]、Ingham等[26]以及邓国成等[27]开展了大口黑鲈的寄生虫病调查,报道了单殖吸虫、复殖吸虫、绦虫、棘头虫、线虫以及其他甲壳动物寄生虫,但均未发现鲺的寄生。Troxel[28]在加利福利亚州13尾大口黑鲈中检测到1只鲺,并将其鉴定为A.flavescens。Cloutman[29]和Mcalister等[30]分别调查了阿肯色州大口黑鲈的寄生虫感染情况,前者发现并报道了A.mississippiensis的寄生和感染,后者分离到一种形态有所差异的鲺,遗憾的是他们未对该鲺作进一步的鉴定。在我国,梁芝源等[31]在300份患病大口黑鲈样本中检测1种鲺的寄生和感染,但该研究仅限于发现报道,尚未对病原在种级阶元上作出鉴定。本研究在湖北省武汉市的大口黑鲈体表分离到1种鲺,并采用形态学和分子生物学的方法,将其鉴定为日本鲺(A.japonicus),通过检索资料发现,这是日本鲺寄生大口黑鲈的首次报道。
鲺的物种鉴定主要是通过比较形态学特征的差异,常用的形态特征包括腹部侧叶的形状[1]、吸盘几丁质条数以及几丁质片的数量[7]等。日本鲺腹部侧叶钝圆,可明显区别于腹部侧叶尖形的物种,如A.coregoni[21]以及喻氏鲺(Argulusyui)[7]等(表1)。另外,王耕南[7]、Wadeh等[2]发现日本鲺吸盘的几丁质条通常由3~8个数量不等的几丁质片组成,雄性(3~4)少于雌性(5~8)。本研究发现,雌性个体几丁质片的数量(5~6)与前人研究相符,但是雄性个体几丁质片数量高达7~8,这可能是由于日本鲺处于的发育阶段不同,导致在形态上有所差异[32-33]。A.foliaceus在形态上与日本鲺相似,肉眼观察难以将二者区分。Yildiz等[34]及Noaman等[35]经过观察发现日本鲺和A.foliaceus的区别在于前者背甲的侧叶末端盖过腹部,且腹部中央裂的深度可延伸至中部,这与本研究结果相符。另外,Soes等[5]发现日本鲺的雄性个体的游泳足基部有基节突,而A.foliaceus无此特征。本研究发现雄性个体的第二游泳足有明显的基节凸起(图3F),进一步证实了Soes等[5]的观点。日本鲺在体色方面也表现出较大差异,本研究中的日本鲺体色淡黄,而在Zoysa等[1]、Wadeh等[2]和王耕南[7]的研究中表现为乳白色,体色的差异可能与宿主的种类及养殖生境有关。
因此,传统的形态学方法鉴定鲺存在一定的难度,尤其是对处于幼体阶段的鲺,几乎无法进行准确鉴定[32]。近年来,分子生物学技术已广泛用于寄生虫的物种鉴定。Sahoo等[36]利用随机扩增多态DNA技术分析了印度13个地区鲺的遗传多样性,发现该技术可以准确区分日本鲺和Argulussiamensis。Patra等[37]利用SSU rDNA序列分析了鲺属内物种的系统发育关系,发现SSU rDNA序列能有效区分该属内的物种。本研究基于SSU rDNA序列构建了ML系统进化树,发现鲺属内物种主要聚为3个支系,拓扑结构与Patra等[37]的相同。然而,日本鲺所在支系(CladeⅠ)中其株系与A.foliaceus和A.rhipidiophorus呈多系类群,日本鲺JN558647株系甚至聚于CladeⅡ中,这给确定日本鲺的系统进化位置带来困难。检索GenBank数据库中的序列信息发现,大部分鲺物种的SSU rDNA序列仅限于序列信息,缺乏相关文献报道的支持,存在误定种的可能。因此,在今后的工作中,应补充更多鲺物种分子数据,同时,结合其他的分子标记探究鲺属物种的系统进化关系。
综上所述,本研究首次报道日本鲺在大口黑鲈体表的寄生,丰富了日本鲺生物学特性的基础数据,增补了大口黑鲈的寄生虫病名录,可为今后大口黑鲈鲺病的诊断和防治提供理论依据。
猜你喜欢
基层中医药(2021年6期)2021-11-02
生物技术通报(2021年4期)2021-05-14
新农民(2020年15期)2020-06-22
创新作文(小学版)(2018年10期)2018-08-20
中国酿造(2017年8期)2017-09-03
广东饲料(2016年7期)2016-12-01
中国环境科学(2015年7期)2015-08-30
浙江柑橘(2013年2期)2013-01-22
华南农业大学学报(2012年2期)2012-11-10
食品工业科技(2011年6期)2011-10-09
