
卤醇脱卤酶研究进展
2021-07-08 03:13丁俊辉侯亚利杨忠华
发酵科技通讯 2021年2期
丁俊辉,袁 林,喻 晨,侯亚利,杨忠华,全 灿
(1.武汉科技大学 化学与化工学院,湖北 武汉 430081;2.安琪酵母股份有限公司,湖北 宜昌 443000;3.中国计量科学研究院 化学计量与分析科学研究所,北京 100013)

对卤醇脱卤酶的机制及其应用已经有相关报道,但对卤醇脱卤酶特别是新发现的卤醇脱卤酶还未见详细的归纳与综述。此外,对卤醇脱卤酶的分子改造及固定化等方面的研究进展还未见相关报道。笔者在对新发现的卤醇脱卤酶归纳总结的基础上,对卤醇脱卤酶的分子改造和固定化进行总结,为卤醇脱卤酶的研究提供新的研究资讯。
1 卤醇脱卤酶的主要来源及亚型
Castro等[6]首次在以2,3-二溴丙醇为唯一碳源的黄杆菌(Flavobacteriumsp.)中发现并分离出卤醇脱卤酶,之后从多种微生物中分离得到大量的卤醇脱卤酶。主要的卤醇脱卤酶的来源及亚型见表1。
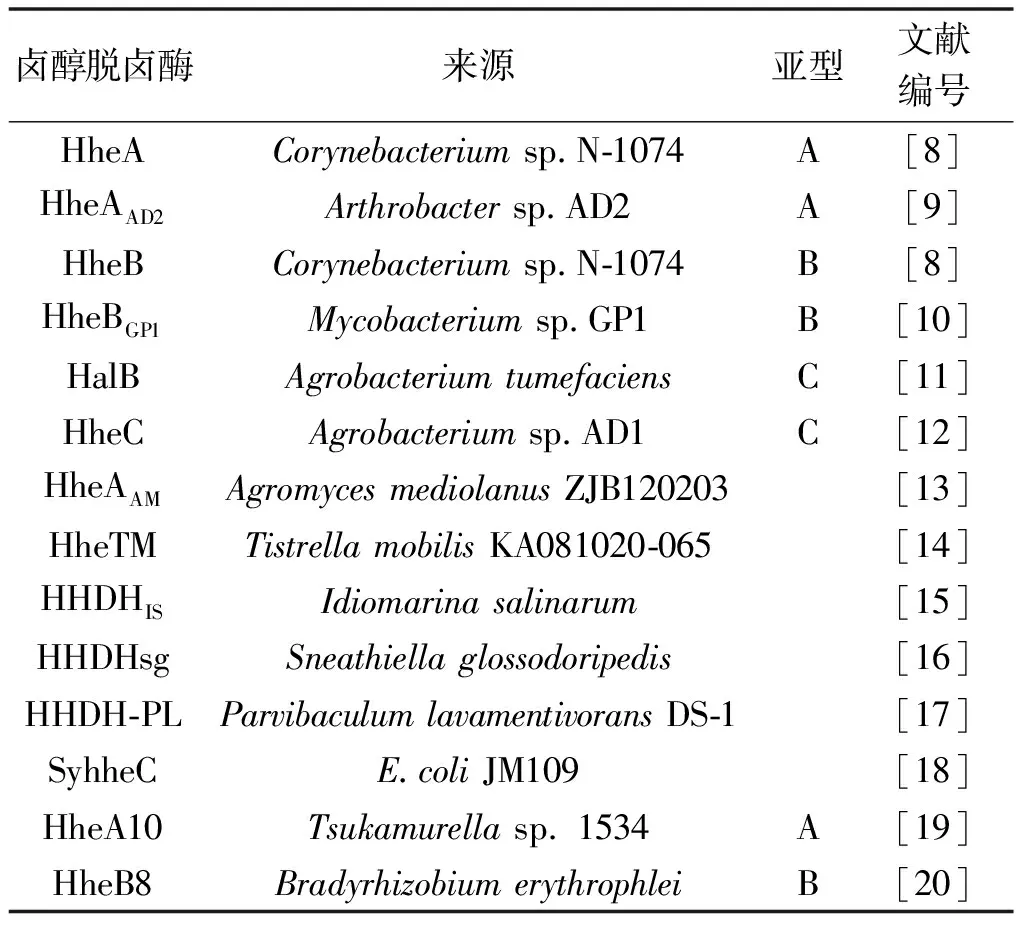
表1 卤醇脱卤酶的产酶微生物及亚型
2014年以前,通常根据其序列同源性将卤醇脱卤酶分为A,B和C 3种亚型,它们之间的氨基酸序列同源性为20%~30%[7]。这3种亚型卤醇脱卤酶的底物范围具有如下特性:A和B亚型卤醇脱卤酶对较长链的邻卤醇(C5,C6)具有较高的催化活性,而C亚型卤醇脱卤酶则适用于催化短链邻卤醇(C2,C3)。此外,C亚型卤醇脱卤酶对脂肪族及芳香族均具有较高的立体选择性,而A和B仅有较低的手性选择性。因此人们对C亚型卤醇脱卤酶的研究更为深入,包括其稳定性、催化机理、动力学反应机制、三级结构及生物催化特性等。
2016年,Koopmeiners等[21]和Anett等[22]利用基因挖掘技术以HHDHs亲核结合口袋结构(T-X4-F/Y-X-G)和第2个催化三联体结构(SX12YX3R)的两个序列挖掘到37种新型卤醇脱卤酶,并对其中17种酶的酶活及立体选择性进行了研究。通过研究其序列同源性,发现部分酶不能分配到A,B和C亚型,通过构建系统发育树分析增加了D,E,F和G 4种亚型。到目前为止,研究最多的是HheC,HheC催化1,3-二氯-2-丙醇生成环氧氯丙烷,底物的转化率可达92.3%[23]。此外,它对2,3-二氯-1-丙醇、3-氯-1,2-丙二醇、2-氯-1-苯基乙醇、对硝基-2-溴-1-苯基乙醇等底物都具有较高的催化活性。当HheC催化2,2-β-氰与叠氮双取代环氧化物生成相应的手性纯环氧化物和β-取代叔醇(e.e.达99%)时,如果甲基在手性中心作为第二取代基将极大增加反应的立体选择性(对映体选择率E>200)[24]。
2 卤醇脱卤酶的分子改造
作为生物催化剂,酶具有优异的区域选择性与立体选择性,在催化合成中日益受到重视。卤醇脱卤酶可以立体选择性地催化合成手性环氧化物与手性邻卤醇,这些手性产物作为手性砌块被广泛应用于精细化工和手性药物的合成。野生型卤醇脱卤酶由于热稳定性不足以及立体选择性不高等原因,不能满足工业应用的要求,因此需要对酶进行分子改造,提高其催化特性,如立体选择性、催化活性、热稳定性、pH稳定性和有机溶剂耐受性等。近几年,国内外研究者采用蛋白质工程技术的定向进化、半理性设计和理性设计等分子进化手段对卤醇脱卤酶进行分子改造。


表2 卤醇脱卤酶的分子进化
目前为止,对卤醇脱卤酶HheC的改造研究最多,这主要是因为人们对其晶体结构及催化机理研究最深,为其理性设计提供了结构与功能关系基础。通常人们对卤醇脱卤酶HheC的活性位点及底物结合口袋进行改造,例如将底物结合口袋部位的氨基酸残基转化为较小的氨基酸以减小空间阻力,或将其转化为疏水基团增加和底物的亲和性,并且可以通过分子对接等手段对其进行理论设计。例如,Wu等[27]采用大引物易错PCR方法和迭代定点饱和突变(ISM)方法对于卤醇脱卤酶HheC构建突变文库,筛选到的最好突变体Var4在65 ℃条件下酶的半衰期提高3 400 倍,在75 ℃下温育30 min后仍保留75%活性。Xue等[28]对卤醇脱卤酶HheC进行理性设计,通过定点突变,所获得的突变体P175S/W249P在pH 8.0下催化1,3-二氯-2-丙醇生成S-环氧氯丙烷的立体选择性从5%提高至95%,在pH 10下其e.e.为92.3%,产率达92.3%。如果再联合环氧化物水解酶,其立体选择性可达99%以上。除了采用现有的突变改造方法,Wang等[29]还创建了高效多位点进化策略,利用该策略对卤醇脱卤酶HheC的碳末端的10个氨基酸进行改造,筛选出的最优突变体PX14半衰期是野生型的17.8倍,kcat是野生型的4倍。该策略不仅大大节约了筛选成本,还使得该酶的催化性能得到了提高。研究表明:在今后的研究中不仅要利用现有的改造策略,也需要根据具体的要求设计新的策略以提高改造效率。除了对卤醇脱卤酶HheC的改造,对其他卤醇脱卤酶如Mikleusevic等[30]进行定点突变,将178位的天冬酰胺转化成丙氨酸,可使得其对苯乙烯氧化衍生物和缩水甘油醚的立体选择性提高。Liu等[31]对来自AgrobacteriumtumefaciensCCTCC M 87071的卤醇脱卤酶进行结构分析,并进行分子对接分析,对活性位点及底物结合口袋周围的14个氨基酸进行定点突变,获得的最优突变体的催化活性比野生型增加26倍,kcat增加18.4倍,45 ℃下半衰期延长3倍。
3 卤醇脱卤酶固定化
游离酶在使用时不易回收,难以重复利用,另外还会对后续产物分离造成影响,同时游离酶也可能对高温、酸碱、离子强度以及有机溶剂等耐受性较差,这些缺陷共同限制了其在工业上的大规模生产应用。卤醇脱卤酶在使用中也存在这些问题。酶的固定化技术是解决上述问题的有效途径之一。通过采用酶固定化技术,可使酶具有更高的稳定性和可重复利用性,具有可操作连续、工艺简单等优点,成为近年来酶工程领域的研究热点[33-35]。
对酶固定化技术的研究具有悠久的历史,而对卤醇脱卤酶固定化的研究是近几年才开始的。酶固定化技术中最重要的是载体选择及固定方法。为了达到不同的目的,人们往往会根据酶的特性及实际应用环境,选择不同的载体及固定化方法。卤醇脱卤酶表面有许多疏水基团,表面疏水性高,故常采用环氧树脂作为固定化载体。由于环氧基团在常温下即可与酶蛋白表面的活性官能团如氨基、羧基、巯基等产生温和的开环共价结合,从而将酶分子固定在载体表面。合成树脂载体在较长的使用周期内具有很强的耐微生物和酸碱腐蚀性以及较强的机械性能,因此当前对卤醇脱卤酶固定化的研究主要是以合成树脂为载体,采用共价结合法进行固定。该方法固定的卤醇脱卤酶具有良好的操作稳定性,可重复利用。2016年,顾恺等[36]将卤醇脱卤酶共价结合在环氧树脂ES-103B上,利用低盐吸附和添加甘油作为酶活中心保护剂,在最佳固定条件下酶活的回收率为74.5%,酶固定化效率为94.2%,比活力为518.6 U/g。在催化1,3-二氯丙醇制备环氧氯丙烷的反应中,重复使用15次后,反应的产物收率还可达到初始收率的91%以上。2018年,Zhang等[37]采用A502Ps树脂固定化卤醇脱卤酶HheC(P175S/W249P)制备S-环氯丙烷,1,3-二氯丙醇的浓度为20 mmol/L时,S-环氧氯丙烷的产率为83.78%,e.e.为92.53%。共价结合固定方法是利用化学方法对酶进行固定。该固定方法所获得的固定化酶虽然稳定性高、重复利用率高,但是固定过程中酶活力损失大。为了避免固定化过程中酶活的损失,利用凝胶、半透膜等材料包埋是一种较好的选择。梁国斌等[38]利用海藻酸钙凝胶包埋卤醇脱卤酶,利用戊二醛进一步交联,制定出固定化脱卤酶微球,催化制备R-4-氰基-3-羟基丁酸乙酯(ATS-5)。在最佳条件下,利用固定化酶微球合成ATS-5,在连续运行10 h后反应转化率可以保持90.6%,ATS-5分离收率达到98.2%,纯度达到99.3%,光学纯度达到99.1%。该固定化酶在4 ℃下保存80 d,无明显的酶活下降。除了常见的固定化方法,还有无载体的交联酶聚集体(CLEAs)固定化技术。2014年,Dvorak等[39]采用该固定化方法,将来自RhodococcusrhodochrousNCIMB 13064的工程化卤代烷脱卤酶、来自AgrobacteriumradiobacterAD1的卤醇脱卤酶与环氧化物水解酶,以纯酶或无细胞提取物的形式进行固定制备CLEAs,使用该固定的酶在填充床反应器内连续降解1,2,3-三氯丙烷(TCP)污染物,反应稳定运行75 d,可以将52.6 mmol/L的TCP连续转化为甘油,TCP转化率为97%,产物甘油的收率为78%。
4 结 论
卤醇脱卤酶已显现出良好的应用前景,开发活性更高、酶学性能更好的新型酶是卤醇脱卤酶研究的热点方向之一。随着宏基因技术、基因挖掘技术等的发展,越来越多新型卤醇脱卤酶被发掘出来,但是很多野生型卤醇脱卤酶因为酶活较低、立体选择性不高和稳定性差等缺点,限制了其在工业中的应用。为解决这一问题,利用定向进化、半理性设计和理性设计等方法对已有卤醇脱卤酶进行分子改造是卤醇脱卤酶研究的另一热点。采用固定化方法可提高酶的稳定性和重复利用率,促进卤醇脱卤酶的工业利用。因此,功能性的卤醇脱卤酶新型固定化载体开发也是将来卤醇脱卤酶研究的方向之一。此外,还应加强在以下几个方面的研究:1)深入解析卤醇脱卤酶的结构,构建其结构与功能之间的关系,以此指导卤醇脱卤酶的进化与改造;2)注重对已有的卤醇脱卤酶新反应的开发,拓宽其催化应用领域;3)结合工业生产特点,根据卤醇脱卤酶的催化特性开发高效的反应器,研究其过程工艺技术。这些工作的开展将极大地推动卤醇脱卤酶的研究及其应用。
猜你喜欢
电气技术(2022年5期)2022-05-23
中国生物制品学杂志(2022年3期)2022-05-13
汽车工程师(2021年12期)2022-01-18
皮肤病与性病(2021年3期)2021-07-30
教育周报·教育论坛(2020年3期)2020-10-21
第一财经(2019年8期)2019-08-26
科技资讯(2018年16期)2018-10-26
科技信息·下旬刊(2018年8期)2018-10-21
作文·初中版(2017年6期)2017-06-16
