腾冲嗜热厌氧杆菌Cas2原核表达及生物信息学分析
2021-07-01 11:04刘原子魏亚琴杨宇泽俞海山万学瑞
微生物学杂志 2021年2期
王 川, 刘原子, 魏亚琴, 毛 婷, 杨宇泽, 俞海山, 张 钊, 万学瑞
(1.甘肃农业大学 动物医学院,甘肃 兰州 730070; 2.陕西梅里众诚动物保健有限公司,陕西 西安 712034;3. 甘肃省微生物资源开发利用重点实验室,甘肃省科学院生物研究所厌氧微生物中心,甘肃 兰州 730000;4.北京市畜牧总站, 北京 100101)
嗜热菌是生活在环境温度50 ℃以上的一类微生物。腾冲嗜热厌氧杆菌(Thermoanaerobactertengcongensis)MB4株是科学家在中国云南腾冲地区热泉分离得到的1株嗜热厌氧菌,其最适生长温度为75 ℃[1],我国自主对其全基因组进行了测序[2]。CRISPR/Cas系统是一种细菌抵抗外源噬菌体的免疫机制,长期研究发现在90%的古生菌及40%的细菌基因组中都存在这种系统[3-4]。CRISPR(clustered regularly interspaced short palindromic repeats)系统由重复间隔序列、前导序列和cas基因组成,其最明显的结构特征是由一段不连续的重复序列(Direct repeat, DR)和插入其中的间隔序列(Spacer sequence)构成的重复间隔序列[5-6],相比常温菌,这些系统和短的重复序列在嗜热菌种中更丰富,数目更多。CRISPR/Cas系统根据Cas蛋白分为3种类型即Ⅰ型、Ⅱ型和Ⅲ型,这3种类型又可以分为Ⅰ-A到Ⅰ-F、Ⅱ-A和Ⅱ-B、Ⅲ-B几种亚型[7]。Cas1、Cas2 蛋白在各类型和亚型的CRISPR/Cas系统中普遍存在且高度保守,被视为探究 CRISPR/Cas系统进化关系的支架[8]。不同的CRISPR/Cas系统中其Cas蛋白的功能各不相同,例如核糖核酸酶、解旋酶、聚合酶等[9]。最近有研究证明,CRISPR位点具有基因表达调控的功能[10]。Cas2是一种保守的核酸内切酶,并且发挥着重要的作用[11-12],其广泛分布于所有含有CRISPR序列的微生物基因组中[13-16]。前期进行了腾冲嗜热厌氧杆菌在不同温度条件下的转录组分析,全局性分析了该菌的嗜热机制[17]。发现腾冲嗜热厌氧杆菌基因组上有3个短的重复序列CRISPR,分别为 C65、C25和C216(数字表示重复数)。腾冲嗜热厌氧杆菌CRISPR/Cas系统属于Ⅰ-A和Ⅲ-B亚型,cas2基因属于Ⅰ-A 亚型[18]。本研究通过生物信息学方法研究Cas2在腾冲嗜热厌氧菌热适应过程的作用。
1 材料与方法
1.1 材料
1.1.1 菌株与质粒 腾冲嗜热厌氧杆菌(Thermoanaerobactertengcongensis)由谭华荣研究员惠赠;质粒pET-28a和pBluescriptⅡKS(+),菌株E.coliDH5α和表达菌株E.coliBL21为本实验室保藏。
1.1.2 培养基 TTE培养基用于腾冲嗜热厌氧杆菌培养[19];培养大肠埃希菌DH5α、BL21的培养基为LB培养基,2×YT培养基用于重组菌的诱导。
1.1.3 主要试剂 限制性内切酶BamHⅠ和XhoⅠ购自TaKaRa公司;T4 DNA Ligase购自Thermo fisher公司;镍柱购自GE公司;氨苄青霉素、卡那霉素、异丙基-β-D-硫代半乳糖苷(Isopropyl-β-D-1-thiogalactopyranoside,IPTG)、EasyPfu DNA polymerase购自北京全式金生物技术有限公司;质粒DNA小提试剂盒、胶回收试剂盒购自天根生化科技有限公司;其他试剂为国产分析纯。
1.2 方法
1.2.1 引物设计 利用primer5参照GenBank中腾冲嗜热菌MB4(AE008691.1)cas2的序列设计引物,克隆引物见表1。
1.2.2 腾冲嗜热厌氧杆菌基因组DNA的提取 腾冲嗜热厌氧杆菌在TTE培养基中75 ℃培养12 h,使用CTAB法提取嗜热菌基因组DNA。
1.2.3cas2基因的扩增 以提取的基因组DNA为模板进行扩增。反应条件:95 ℃预变性6 min;95 ℃ 35 s;50.7 ℃ 45 s;72 ℃ 2 min;共35个循环。将cas2和pET-28a双酶切,16 ℃连接,将其转化到DH5α中,提取质粒后酶切鉴定并送至生物公司测序。

表1 试验分析所用引物
1.2.4 重组蛋白的表达及纯化 将测序正确的重组质粒转化至大肠埃希菌BL21(DE3)感受态细胞中,挑取单菌落过夜培养后按1∶100比例转接入100 mL 2×YT(Kan+)培养基中,37 ℃培养2.5 h至菌液OD值(600 nm)达到0.4~0.6,加入终浓度为1 mmol/L的IPTG在26 ℃诱导表达7 h,7 000 r/min离心10 min收集菌体,PBS缓冲液洗涤2次后超声波破碎30 min(40%功率,工作5 s,间歇5 s),7 000 r/min离心10 min[20]。收集上清并用GE公司的Ni-NTA柱进行蛋白纯化,以诱导前的重组菌株为对照进行SDS-PAGE电泳,分析重组蛋白表达情况。
1.2.5 实时荧光定量PCR 在50、60、75和80 ℃条件下培养腾冲嗜热厌氧杆菌。RNA提取及RT-PCR方法参照Liu等[19]的文章。引物序列见表1,以16S RNA作为内参基因,用相对定量的方法检测不同温度下cas2基因的表达量变化[21]。荧光定量数据的计算方法以2-△△Ct方式计算[22],采用SPSS软件进行ANOVA单因素方差分析,P<0.05表示有显著性差异。
1.2.6cas2及编码蛋白的生物信息学分析 选取腾冲嗜热厌氧杆菌(AAM25776.1)、沙门氏菌(AEB71782.1)、嗜酸革兰杆菌(KJE29018.1)、单核细胞增生李斯特氏菌(KFL19553.1)、嗜盐杆菌(AXH10449.1)cas2基因,利用在线软件Prot-Param (http://www.expasy.org/tools) 分析该基因序列编码蛋白Cas2氨基酸序列组成与理化性质;应用软件ProtScale (https://web.expasy.org/cgi-bin/protscale/protscale.pl) 分析其氨基酸残基疏水性;应用软件TMHMM Server v.2.0 (http://www.cbs.dtu.dk/services/TMHMM) 预测跨膜区;利用软件SignalP 3.0 (http://www.cbs.dtu.dk/services/SignalP-3.0) 预测蛋白信号肽;利用软件PredictProtein (https://www.predictprotein.org/home)预测蛋白质二级结构;运用 STRING 数据库(http:/ /string-db.org),设置为高置信度0.7,不限制数量,构建与 Cas2相互作用的蛋白网络。
2 结果与分析
2.1 cas2的克隆和表达载体的构建
通过PCR方法克隆得到了腾冲嗜热厌氧杆菌cas2基因,测序结果表明cas2大小为264 bp。所得cas2基因序列比对同源性为100%,将cas2连接到pET-28a,转化大肠埃希菌DH5α,构建的重组质粒pET-28a::cas2经BamHⅠ和XhoⅠ酶切验证无误后转化大肠埃希菌BL21,cas2PCR结果和质粒双酶切验证图谱见图1。
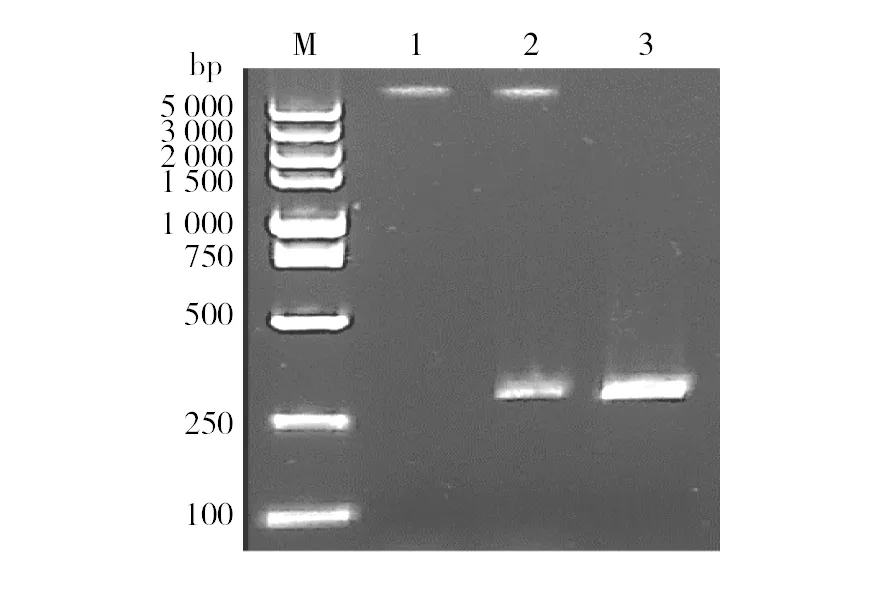
图1 cas2基因的PCR扩增产物及重组质粒双酶切Fig.1 Electrophoresis of PCR products of cas2 gene and recombinant plasmid digested M: DNA分子质量标准DL5000;1: pET-28a载体; 2: pET-28a::cas2重组质粒双酶切;3: cas2 pcr产物M: DL5000 DNA Marker; 1: pET-28a vector; 2: Double endonuclease digestion product ofpET-28a::cas2;3: PCR products of cas2 gene
2.2 Cas2蛋白诱导表达及SDS-PAGE电泳
pET-28a::cas2重组菌株经IPTG诱导,超声破碎后进行纯化,得到蛋白Cas2并进行SDS-PAGE电泳检测。SDS-PAGE分析表明,诱导前无特异条带,诱导后出现特异条带,纯化后在10 ku附近出现单一条带(图2)。
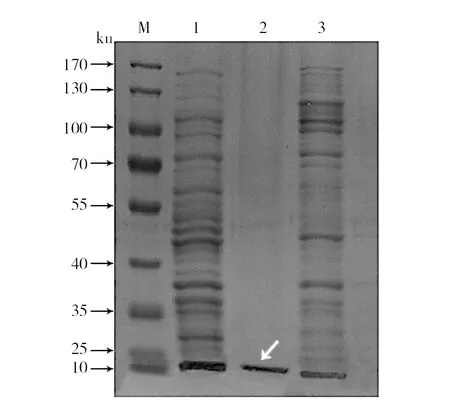
图2 Cas2表达产物的SDS-PAGE分析Fig.2 SDS-PAGE analysis of the Cas2 expressed productM: 蛋白分子质量标准;1: 诱导后的pET-28a::cas2菌液;2: 纯化后的pET-28a::Cas2蛋白; 3:未诱导的pET28a::cas2菌液M: Protein molecular weight Marker; 1: pET-28a::cas2 induced with IPTG; 2: Purification recombinant expressed protein of Cas2; 3: pET-28a::cas2 not induced with IPTG
2.3 cas2在不同温度下的转录分析
将50 ℃的表达量定义为1,结果显示在60、75和80 ℃表达量是相对于50 ℃表达量的22.2、39.3和12.8倍(图3)。可以看出在50到75 ℃,cas2表达量随温度升高而升高,cas2基因在60 ℃和75 ℃高表达,说明cas2表达量与温度有关。

图3 cas2在50、60、75和80 ℃下的表达分析Fig.3 The expression analysis of cas2 under50, 60, 75 and 80 ℃ 16S RNA作为内参基因,50 ℃时表达量作为116S RNA was used as internal control, and the transcription at 50 ℃ was arbitrary assigned as relative 1
2.4 cas2基因及其编码蛋白氨基酸的生物信息学分析分子特征
cas2基因序列及编码蛋白序列的分子特征:通过在线软件EXPASY分析得到腾冲嗜热厌氧杆菌cas2完整ORF全长264 bp,编码84个氨基酸,蛋白分子质量约为9.9 ku,等电点为9.31,分子式为C473H691N115O120S1。通过生物信息学分析比较了cas2基因在腾冲嗜热厌氧杆菌、沙门氏菌、嗜酸革兰杆菌、单核细胞增生李斯特氏菌和嗜盐杆菌所编码氨基酸的基本理化性质(表2)。
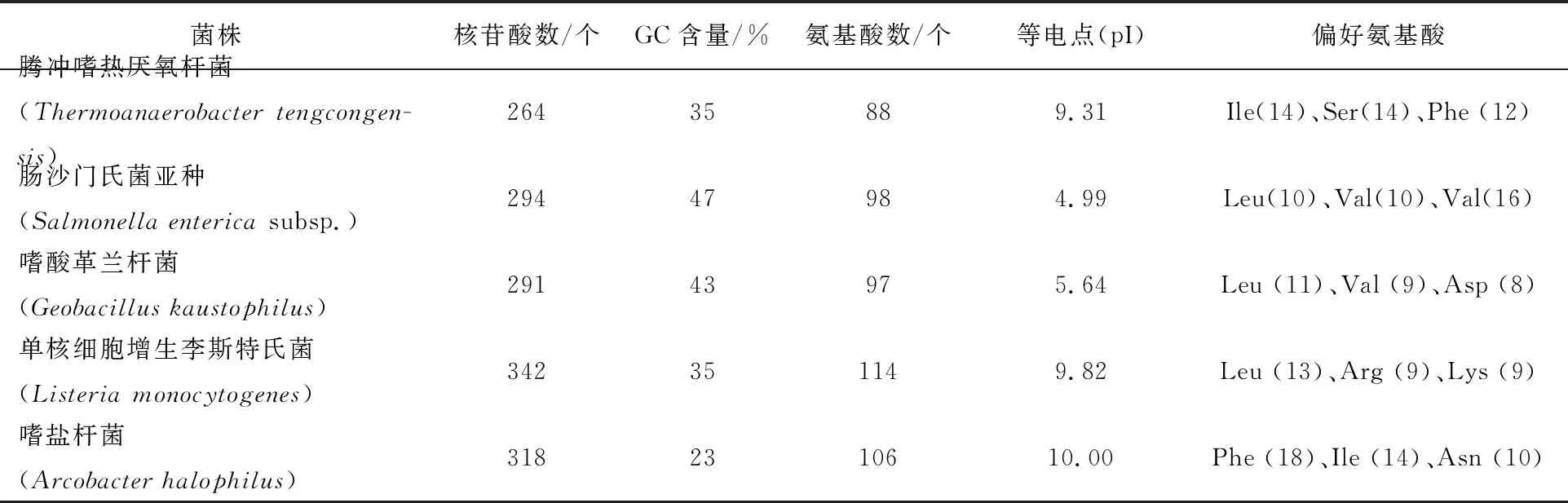
表2 cas2基因及编码蛋白的生物学特征
2.5 cas2基因编码蛋白质的疏水性分析
结果显示,Cas2蛋白的最大疏水性为1.733,最小为-1.233;不稳定系数为40.16,脂肪系数为96.31,最终亲水性的平均值(GRAVY)为0.280(见图4A),且不存在跨膜区域(见图4B)。
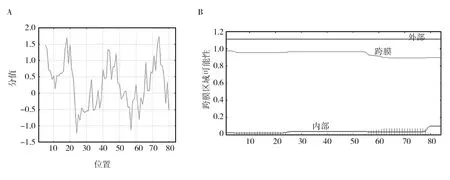
图4 腾冲嗜热厌氧杆菌Cas2蛋白疏水性分析和跨膜结构预测Fig.4 Hydrophobicity analysis and prediction of transmembrance structures of Cas2 protein of Thermoanaerobacter tengcongensisA: Cas2蛋白的疏水性分析; B: Cas2蛋白的跨膜结构预测A: Analysis of hydrophilic/hydrophobic properties of Cas2; B: Prediction of transmembrance structures of Cas2
2.6 腾冲嗜热厌氧杆菌Cas2蛋白二级结构分析
使用SOPMA软件对Cas2蛋白的二级结构进行预测,发现该蛋白主要由α-螺旋、β-折叠、β-转角和无规则卷曲4 种常见的蛋白质二级结构组成。其中β-折叠所占比例最高,为34.52%;其次是α-螺旋,所占比例为28.57%;β-转角和无规则卷曲所占比例分别为13.10%和23.81%(图5)。
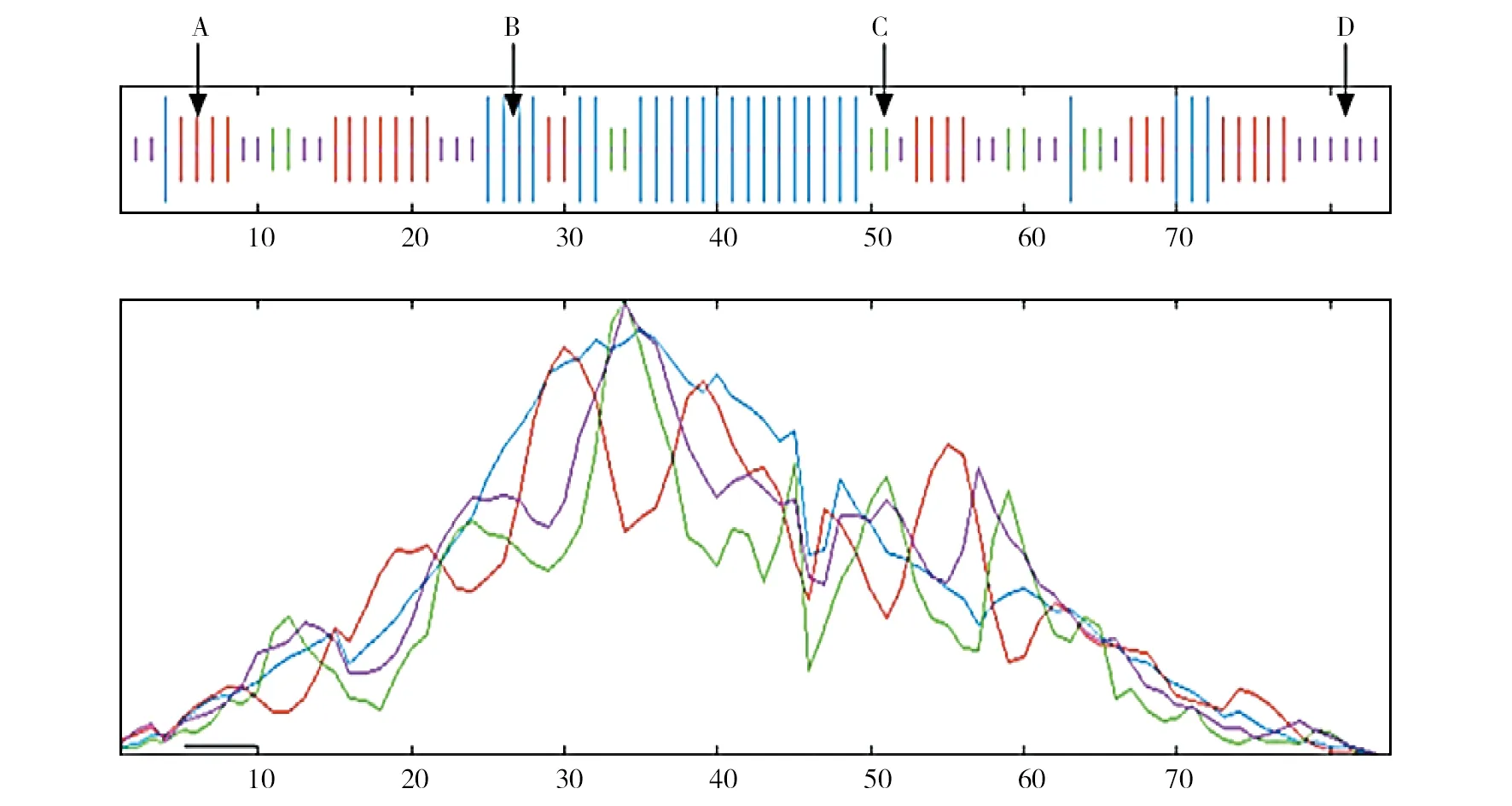
图5 腾冲嗜热厌氧杆菌Cas2蛋白二级结构预测Fig.5 Prediction of secondary structure of Cas2 protein in Thermoanaerobacter tengcongensisA:α-螺旋;B:β-折叠;C:β-转角;D:无规则卷曲A: represents the alpha helix; B: represents beta fold; C: represents beta turn;D: represents the Random curls
2.7 腾冲嗜热厌氧杆菌Cas2蛋白质相互作用分析
使用 STRING 数据库搜索与 Cas2(TTE2659) 相互作用的蛋白质信息,设置为高置信度 0.7,不限制数量,构建Cas2蛋白相互作用网络,结果显示Cas2 (TTE 2657)与 Cas1(TTE 2658)、Cas4 (TTE 2659)、Cas3 (TTE 2660)、Cas5 (TTE 2661)、Cas7 (TTE 2662)、Cas6 (TTE 2664)、 Cas8a (TTE 2666)和 Cas6a (TTE 2667)等存在互作关系(图6)。
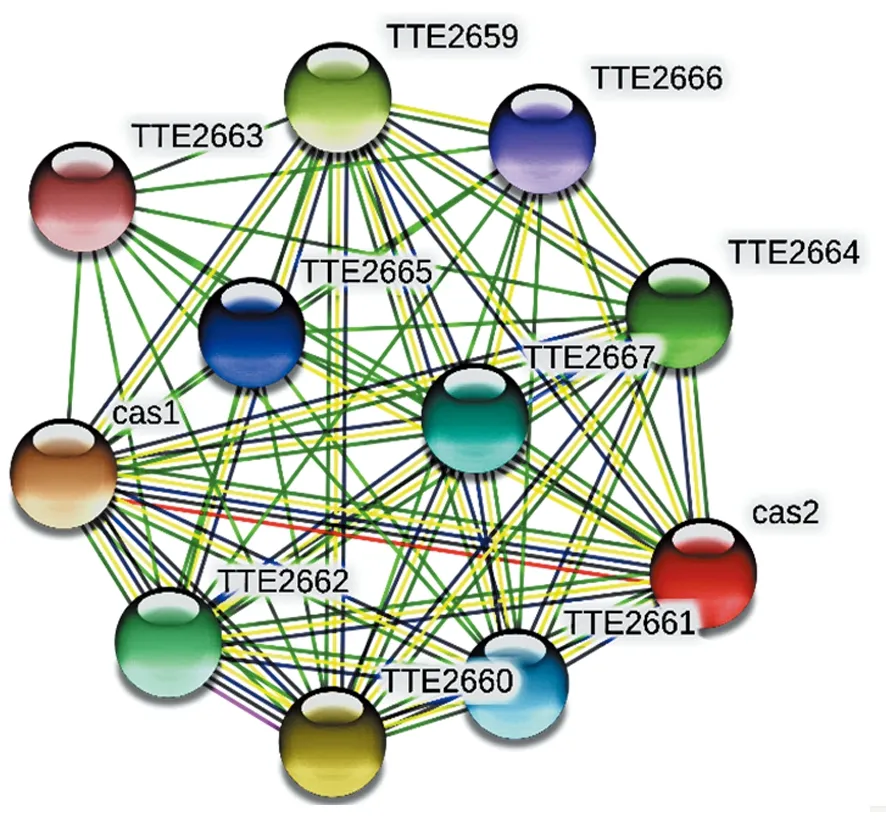
图6 腾冲嗜热厌氧杆菌Cas2蛋白互作蛋白网络Fig.6 Cas2 protein interaction protein network of Thermoanaerobacter tengcongensis
2.8 cas2基因的遗传进化分析
选取不同嗜性的菌株包括肠炎沙门菌(Salmonellaentericasubsp.entericaserovarTyphi)、发酵乳杆菌(Lactobacillusfermentum)、青春双岐杆菌(Bifidobacteriumadolescentis)、嗜盐杆菌(Arcobacterhalophilus)、嗜酸菌(Delftiaacidovorans)、嗜热芽胞杆菌(Geobacilluskaustophilus)、嗜热链球菌(Streptococcusthermophilus)和腾冲嗜热厌氧杆菌(Thermoanaerobactertengcongensis);嗜热芽胞杆菌(Anoxybacillussp. B7M1)和单核细胞增生李斯特氏菌(ListeriamonocytogenesFSL F6-684)等菌株,以这些菌株的cas2基因建立系统进化树,菌株信息结果腾冲嗜热厌氧杆菌cas2基因与嗜热芽胞杆菌(An-oxybacillussp. B7M1)同源性最高(图7)。
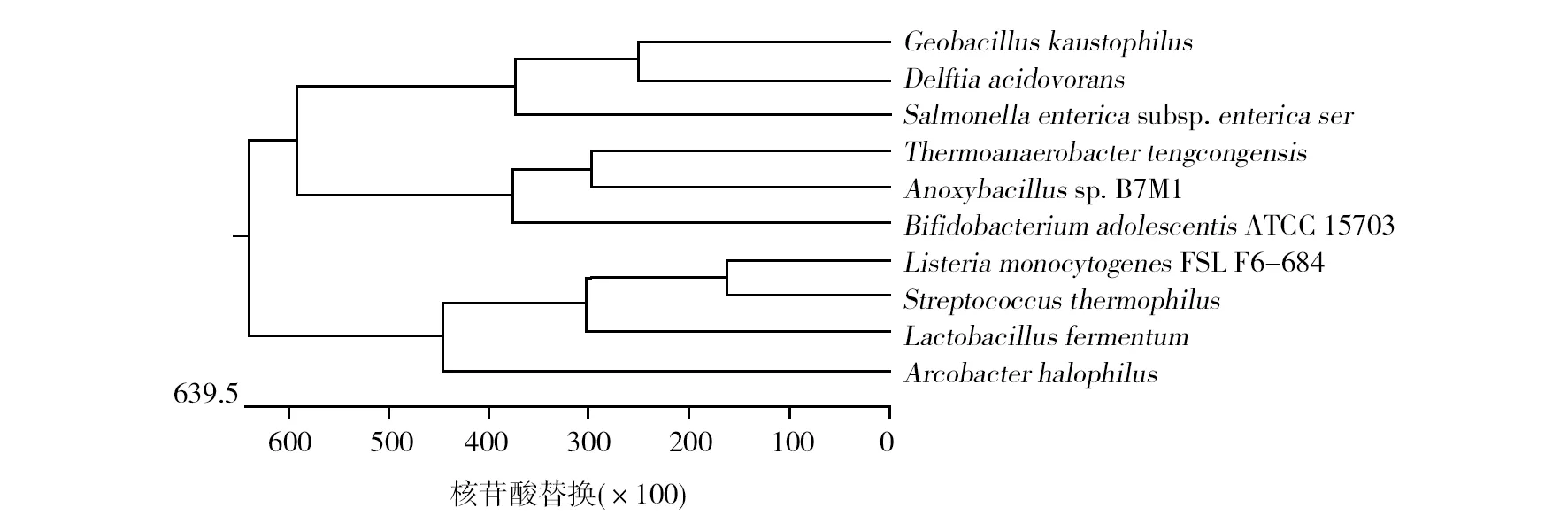
图7 cas2基因系统进化树Fig.7 Phylogenetic tree of cas2
3 讨 论
CRISPR系统对外源DNA的入侵具有获得性免疫功能,这种免疫功能又被称为干扰作用,其发挥功能分为适应、表达和干扰三个阶段[23]。除了Ⅲ-C型,Ⅲ-D型和Ⅳ型CRISPR系统外,在CRISPR系统的适应阶段主要是由Cas1和Cas2发挥作用[24]。Cas1是一种整合酶,Cas2是mRNA干扰素的同源物,两者能识别入侵的外源病毒或质粒DNA的PAM序列[25],在Cas2蛋白复合物作用下,Cas1切割识别的DNA序列并在其他酶的辅助下整合到CRISPR序列中,从而完成外源DNA的捕获[26]。
本研究从生物信息的角度出发分析腾冲嗜热厌氧杆菌cas2及其编码蛋白的理化性质,结果显示Cas2蛋白为9.9 ku左右的亲水性蛋白,不存在跨膜区域,氨基酸组成以Ile(14)、Ser(14)、Phe (12)为主,且该蛋白主要由α-螺旋、β-折叠、β-转角和无规则卷曲4 种常见的蛋白质二级结构组成。使用 STRING 数据库搜索与 Cas2 相互作用的蛋白质信息结果显示Cas2与 Cas1、Cas4、Cas3、Cas5、Cas7、Cas6、 Cas8a 和Cas6a等存在互作关系,值得注意的是上述Cas蛋白构成 CRISPR-Cas I-A 亚型系统,说明Cas2发挥作用是一个复杂的过程。本研究发现腾冲嗜热厌氧杆菌在50~75 ℃时,cas2表达量随温度升高而升高,cas2基因在60 ℃和75 ℃条件下高表达,暗示cas2表达量与温度有关。根据cas2基因建立的进化树显示腾冲嗜热厌氧杆菌与厌氧菌芽胞杆菌同源性高,说明Cas2蛋白在嗜热菌中发挥着特殊的作用。本研究结果对腾冲嗜热厌氧杆菌CRISPR及其相关蛋白质在热稳定性机制中的研究提供了重要的信息,为今后嗜热菌嗜热机制的研究提供参考。
猜你喜欢
四川轻化工大学学报(自然科学版)(2021年1期)2021-06-09
江西农业学报(2021年4期)2021-04-20
今日农业(2020年13期)2020-12-15
云南画报(2020年11期)2020-12-14
汉字汉语研究(2020年2期)2020-08-13
电子制作(2019年22期)2020-01-14
动漫星空(兴趣百科)(2019年4期)2019-04-09
疯狂英语·新读写(2018年3期)2018-11-29
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01