
秒尺度温室番茄作物-环境互作模型构建与验证
2021-06-29 01:45:14徐立鸿孟凡峥蔚瑞华
农业工程学报 2021年8期
徐立鸿,孟凡峥,蔚瑞华
秒尺度温室番茄作物-环境互作模型构建与验证
徐立鸿,孟凡峥,蔚瑞华
(同济大学电子与信息工程学院,上海 201800)
为了解决现有温室模型时间尺度不统一的问题,该研究建立了一个时间尺度统一的温室番茄作物-环境互作模型,描述作物与环境之间的相互作用,提高模型的精准性。首先,将番茄作物生长模型拆分成SUPPLY、PARTITION、GROWTH 3个子模块,针对3个模块在由天数量级时间尺度到秒数量级时间尺度变换时存在的问题,通过模型替换、结构改造、参数辨识等方法对时间尺度进行了转换,并利用EFAST敏感性分析算法将模型中的不确定参数分为敏感参数和不敏感参数两类。然后,在秒时间尺度番茄作物生长模型的基础上,考虑番茄作物对温室环境的实时反馈,结合小气候模型形成包含未知参数的“通用”的互作模型结构。最后,利用贝叶斯优化方法及番茄生产温室的实际数据,分别对互作模型中生长模型和小气候模型的未知参数进行参数辨识,确定互作模型全部结构与参数,得到可用的互作模型。利用该研究得到的秒时间尺度生长模型对2015—2018年上海崇明A8温室番茄产量进行模拟,其与真实产量值间的均方根误差在7.34~18.85 g/m2之间,平均相对误差在5.8%~18%之间,均小于TOMGRO模型与Integrated模型,可以更好地预测产量变化。含作物反馈的小气候环境模型经参数辨识后,模拟番茄作物3个不同生长时期(幼苗期、开花坐果期、结果期)的环境因子(温室内温度、湿度、CO2浓度)变化的平均相对误差均在3%~6%之间,且相较于未考虑作物反馈的一般小气候模型有更好的模拟效果。互作模型的建立将作物与温室小气候环境统一成一个模型,可以为温室环境控制提供模型基础。
温室;模型;番茄生长模型;小气候模型;敏感性分析;贝叶斯优化
0 引 言
中国的温室设施面积已突破210万hm2,稳居世界第一[1],但目前国内大多数的温室,存在着依赖人工经验控制等问题,导致温室生产经济效益的低下。以世界种植面积最广泛的番茄为例,2016年荷兰的温室番茄年产量达到了90 kg/m2,而同年中国温室番茄年产量仅有34.5 kg/m2[2]。中国大多数温室都缺乏对温室环境的有效调控,模型与控制算法缺失是温室环境无法有效调控的重要原因之一。
温室是一个复杂的多输入多输出的时延系统,具有参数多、非线性、强耦合的特点[3],其具有高复杂性的主要原因在于温室系统中时间尺度不统一[4-6],作物生长模型是为了描述作物生长过程中的动态变化,需要反映出作物所处的环境变量、不同生长阶段等因素对作物生长的影响。机理模型如HORTISIM模型[7]、TOMGRO模型[8]、TOMSIM模型[9]等,都能够反映温室内温度、CO2浓度、光照等环境变量对作物状态变量如器官数量、干物质量、叶面积指数、产量等的影响。Lin等[10]综合比较了TOMGRO模型、Vanthoor模型以及相关生长模型,建立了泛化性更好的番茄作物产量预测模型(后称Integrated模型)。除了机理模型之外,还有很多基于经验与数据建立的模型,如倪纪恒等[11]建立了辐热积与番茄作物分配指数和收获指数的关系,对番茄干物质分配与产量进行了预测。大多数的生长模型都是以天为时间尺度,因此一般认为是一个天时间尺度的慢过程。小气候模型是提高温室环境控制水平的重要基础,是温室控制的直接控制对象,也作为生长模型的输入从而影响最终产量。Vanthoor等[12]建立了有17个状态变量的复杂模型,并使用多地的温室数据进行了验证;胥芳等[13]基于质热交换的物理过程建立了玻璃温室中温湿度的动态模型,能够有效预测室内的温湿度。相比于作物模型,小气候模型则是一个秒时间尺度的快过程,室内外气候以及温室执行机构动作以分钟甚至秒时间尺度变化,因此室外气候的快速波动以及控制系统的动态变化无法通过作物生长模型体现。Dong等[14]指出,只有当集成作物模型时,温室优化控制系统才能获取经济最优的控制效果。Jones等[8]在TOMGRO模型中提到,为了满足控制需要,应该建立短时间步长计算的模型。仅考虑温室中某一个子模型不能达到控制效果的“最优”,如Schmidt等[15]只考虑了慢子过程,基于步长为9d的温室离散模型,每9d计算一次最优控制解来校正模型与室外气象预测的误差,虽然结果显示可以提高产量,但显然不是经济效益最优的策略。邓璐娟等[16]提出将控制系统分为管理层、优化层和实施层,将一天分为4个阶段,用遗传算法来求解最优设定值。Henten等[17-18]利用奇异摄动理论进行温室系统时间尺度分解的方法,将慢过程进行伪静态化计算后得到的慢状态传递到快子问题的控制目标进行求解,建立起了2个时间尺度层级之间的联系,但这会给控制器的设计带来一定的复杂性。
综上所述,目前温室环境与作物模型存在的问题集中在以下几方面:
1)温室作物与环境系统中存在多种时间尺度,“天”时间尺度的作物生长模型难以将“秒”时间尺度的温室小气候环境模型的动态变化直接反映在作物生长上。
2)目前大多数模型仅将小气候环境模型的输出作为作物模型的输入,在温室环境模型与作物模型联合使用时,忽略了作物生长对小气候的实时反馈作用;
3)多时间尺度增加求解温室最优控制问题的控制器设计的复杂性。
因此,本研究针对当前模型中存在的问题,提出秒时间尺度的温室番茄作物-环境互作模型。互作模型包括生长模型与小气候模型2部分,二者相互耦合,时间尺度统一。以秒为计算步长的番茄生长模型,旨在将作物生长与温室环境的时间尺度进行统一,因此可以将快速变化的外界气象以及执行机构动作直接反映在作物状态变化上,由此在进行在线环境控制时可以简化因时间尺度不统一带来的控制器设计复杂性。
1 秒时间尺度生长模型的建立
1.1 模型结构
秒时间尺度生长模型目的为在秒数量级的时间尺度描述番茄作物的动态生长过程。天数量级时间尺度的生长模型,如Integrated模型、TOMGRO模型等均基于源库理论将模型分为SUPPLY(源)和DEMAND(库)两部分,由各个器官的库强来调节干物质分配。这样的模型注重天级别的干物质积累和分配,而并不适用于在秒级别描述作物动态。因此,为了克服上述问题,本研究在保证秒时间尺度生长模型继承了天时间尺度模型中一些有重要机理意义的参数与结构(如作物器官的分组、茎节数等)的同时,将模型分为3个子模块:干物质生产子模型(SUPPLY)、干物质分配子模型(PARTITION)和作物生长子模型(GROWTH)。各个模块与其他天时间尺度模型不同点在于:
1)SUPPLY子模型需要能够从秒级别描述作物的光合和呼吸作用,反映实时环境状态如温度、光照与二氧化碳等对光合产生同化物的速率的影响。
2)PARTITION子模型需要能够模拟SUPPLY子模型产生的干物质实时累积和分配给各个器官的动态过程。
3)GROWTH子模型能够模拟作物各个器官的数量与干物质量、叶面积指数等作物状态变量在生长期的动态变化。
因此本研究针对3个子模块的不同点做出如下的模型建立和调整:
1)选择秒级别的光合模型如FvCB模型描述干物质生产过程,对光合模型参数根据实际温室场景进行参数辨识。
2)采用基于异速分配[19]理论的干物质分配模型等,引入同化池结构,建立实时变化的环境变化(秒时间尺度)和长期的作物状态(天时间尺度)与分配的关系,使得光合模型产生的同化物也可以在秒级别实时地进行分配。
3)作物器官数量变化缓慢,对器官干物质量的影响微乎其微,且在基于模型的控制中一般不作为评价控制的指标,因此放在天时间尺度描述;而作物干物质量、叶面积则作为模型输出可以直接对控制效果进行评价,因此需要在秒时间尺度进行描述,其中秒数量级别的参数如器官的发育率通过参数辨识进行确定。
最终秒时间尺度生长模型的状态变量可以表示为
式中W、W、W分别为果实、叶片和茎干的干物质量, g;N、N、N分别为果实、叶片和茎节的数量,LAI为叶面积指数,m2/m2。模型整体可以表示为
式中为温室内的小气候环境状态变量,x为温室作物的状态变量,为温室结构的输入向量。
1.2 子模型
1.2.1 SUPPLY子模型
SUPPLY子模型模拟作物产生并进入到同化池的干物质量,包含光合作用产生同化物和呼吸作用消耗同化物两部分。光合速率通过Farquhar等[20]提出的FvCB模型计算得到每秒的光合速率P,g/m2·s。番茄作物呼吸作用每秒消耗的同化物R可以表示为
式中()为维持呼吸的温度影响因子,与温度直接相关;RESP为叶片与茎干的维持呼吸系数,RESP为果实的维持呼吸系数,g/g·s。每秒的净光合作用即为总光合与呼吸的差,转换为进入同化池的干质量SUPPLY为
式中为同化物与干物质的转换系数;root为分配给根用于根生长的干物质比例。
1.2.2 PARTITION子模型
描述干物质分配的源库模型通过分析源器官产生和库器官生长需要干物质的比例来模拟分配过程,但秒数量级别的参数如库强度难以获取。并且当干物质生产>干物质分配时,源库模型一般将多余库强的干物质抛弃掉,在快速变化的秒时间尺度,这种做法会带来较大的误差。因此,在秒时间尺度生长模型中引进了同化池的概念,以及选用异速分配法来描述作物的干物质分配。同化物由源器官产生并放入同化池,各个库器官每秒从这个同化池获取同化物,同化池中同化物量的变化为
式中MCPoolLeaf、MCPoolStem、MCPoolFruit分别为同化池分配给叶片、茎干和果实器官的分配系数。
同化池中同化物的分配受实时的环境变化如瞬时温度和当天平均温度T影响,也受长期的作物状态如作物发育阶段(Development Stage, DVS)影响,可以表示为
式中Pool为同化池中同化物量,g;()、(T)、DVS分别为瞬时温度、平均温度和作物发育阶段对分配系数的影响因子,Leaf、Stem、Fruit分别为20 ℃时叶片、茎干和果实每秒的潜在分配系数,g/s;LPN(Leaf Per Node)为作物稳定产出果实时叶片与茎节的数量比。
在作物开花坐果之前,果实的干物质量时刻为0,同化池向果实器官分配同化物的系数也为0,因此DVS=0;当作物进入果期并稳定产出果实时,发育阶段影响因子DVS接近于1。因此总茎节数N与第一个果实所在的茎节数First存在负指数关系
式中First取值16,由现场人工获取;为不确定参数,需要进行参数辨识。同化池分配给各器官后,器官还需要将分配的干物质再分配给各个年龄组。
1.2.3 GROWTH子模型
番茄作物的结构包括叶片、茎干、果实和根4个部分,叶片、茎干和果实为作物的地上部分,它们的数量与干物质量是生长模型进行模拟的最主要的参数。作物器官数量一天内的变化较少,且基于模型的控制最终目标倾向于产量(干物质量)最大化,因此对于作物器官数量完全可以放在天数量级别描述,而与控制目标直接相关的叶面积以及器官干物质量,则应该在秒数量级别描述。以果实为例,其数量与干物质量和茎节的关系可以表示为
式中△N(,)、△N(,)分别是第天第个年龄组茎节和果实的数量变化;FPN(Fruit Per Node)是稳定产出果实时新生果实与新生茎节的比例,取值为2.67,由人工获取;MCPoolFruit()为分配给第个年龄组的干物质;r分别为秒级别的果实发育率,n是作物的生理年龄组数;P是收获导致的果实数量损失。
叶面积的增长与叶片干物质量及比叶面积SLA相关,当叶面积指数大于临界叶面积指数XLAIM时,会通过剪枝操作使叶面积指数LAI维持在XLAIM不变,因此可以表示为当LAI超过XLAIM时,其导数为0
式中XLAIM取值为3.5,由人工设定。
1.3 生长模型参数敏感性分析
作物生长与其所处的地域环境密切相关,生长模型参数需要根据所处的环境数据进行调整以达到最小的预测误差。因此在生长模型结构确定后,需要对模型中大量不确定的参数(见表1)进行分类,一部分对结果影响较大的参数需要根据实际数据进行辨识,另一部分对结果影响较小的参数可以采用固定值,以降低模型辨识的难度。
敏感性分析用于分析相关变量对模型输出的影响程度,以区分敏感参数和不敏感参数[21-22],进而来帮助我们区分变量是否需要辨识。
借助专业敏感性分析软件Simlab进行试验,使用拓展傅里叶幅度检验法(Extended Fourier Amplitude Sensitivity Test,EFAST)对生长模型进行敏感性分析,表1中参数的参数取值范围均为均匀分布。得到的各个参数的一阶敏感性指数和全局敏感性指数(如图2):
采用Dejonge[23]在EFAST法中界定的S>0.05,S,i>0.1来确定敏感性参数,S、S,i分别为参数的一阶敏感度和全局敏感度。从敏感性分析的结果来看,生长模型中13个不确定参数可以分为敏感参数和不敏感参数两部分,敏感参数包括、E、c、、MAX25,Leaf、Fruit、Leaf、,它们需要根据实际温室的数据进行辨识后确定;不敏感参数包括r、r、r、RESPL、RESPF,可以直接从表1中选取固定值,本文选取的参数值见表2。
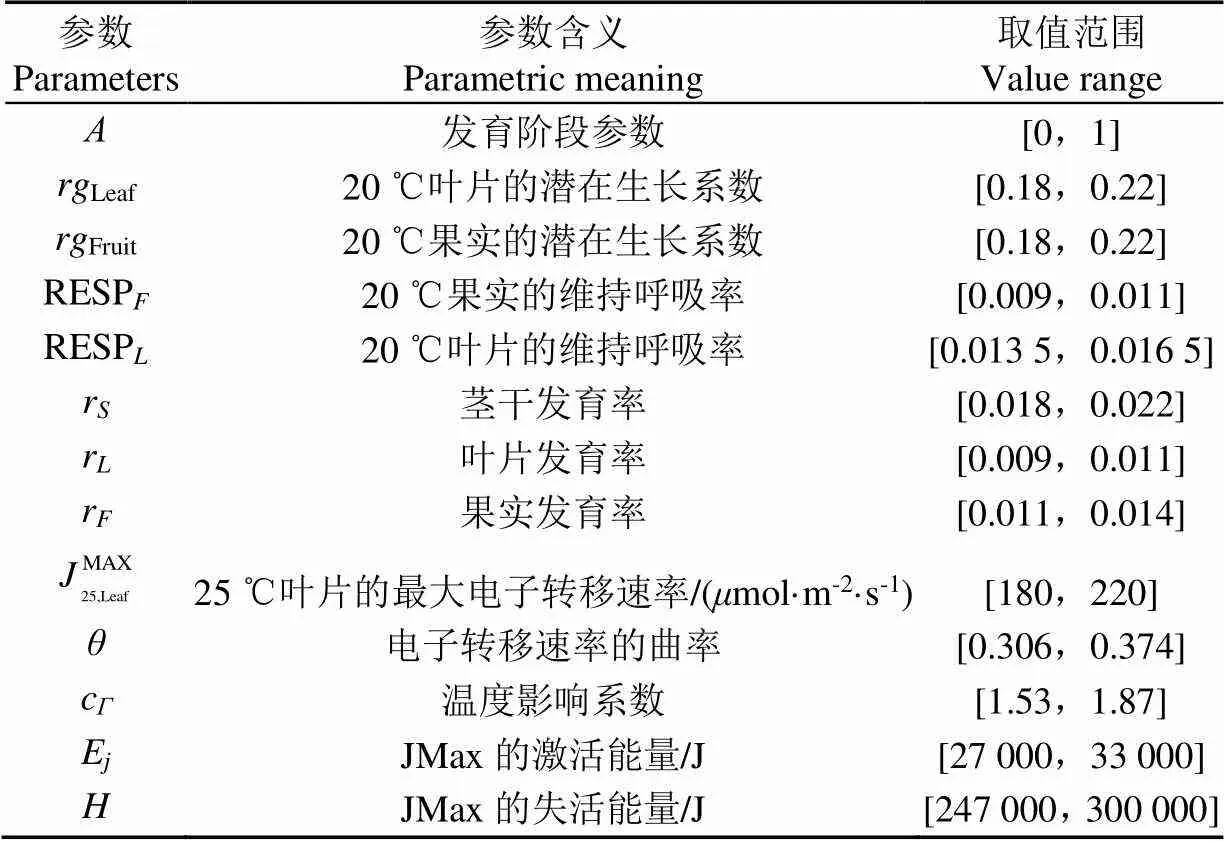
表1 秒时间尺度作物生长模型参数

表2 不敏感参数选取的固定值
2 含作物反馈影响的小气候环境模型
2.1 含作物反馈的小气候模型结构
温室小气候机理模型的研究已经相对成熟,但小气候模型在环境控制时只作为作物模型的输入而存在,即考虑了小气候模型对作物模型的作用,而未考虑作物模型对小气候模型的反馈作用。已有典型的小气候模型可以表示为
式中、、分别为执行机构、室外气候、温室结构的输入向量,小气候状态包含冠层温度Can、温室内空气温度Air、温室地表温度Flr、温室内空气湿度Air、温室内CO2浓度CO2,Air
由于缺乏对作物对环境实时反馈这部分机理的量化,大多数小气候模型在温室中长期使用时会出现误差变大等问题。而互作模型结合作物模型x与小气候模型,将作物对小气候环境的实时反馈进行量化,可以解决上述小气候模型存在的误差变大的问题。因此,含作物反馈影响的小气候模型可以表示为以下形式
2.2 作物对温度动态的反馈
作物对温度动态的反馈过程反映在作物冠层温室内辐射的拦截,以及作物蒸腾作用对温室内热量传递的影响上。随着作物的生长,叶面积指数不断变大,作物冠层会对太阳辐射进行不同程度的拦截,从而导致太阳辐射在温室中不同的传输规律。冠层的光学参数受叶面积指数影响
冠层吸收的总的辐射能量为
式中为冠层的消光系数,α、τ、ρ分别为冠层的吸收系数、透射系数和反射系数,ρ为地表的反射系数,η为温室内空气的吸收率,η为冠层的吸收率,I为温室内总辐射,W/m2。地表吸收的辐射能量包括从冠层透射的辐射以及经二次反射回到地表的辐射,最终地表吸收的辐射能量为

2.3 作物对湿度动态的反馈
温室中的湿度会动态的变化,与作物的蒸腾作用都有很大的关系,是作物对环境反馈的一个重点体现。尤其在作物发育后期叶面积比较大时,作物蒸腾作用对湿度的影响不可忽略。由于现代温室为了预防病虫害,大多数会选择铺设水泥地面或者覆盖膜,因此不考虑土壤蒸腾对室内湿度的影响,即地表与空气的水汽交换φ,flr恒为0。
根据Stanhellini[24]提出的基于P-M模型的番茄蒸腾速率模型,作物的蒸腾量为
式中为湿度计算常数,r、r分别为边界层和叶片气孔的阻抗,VP为冠层的饱和蒸气压。
2.4 作物对CO2动态的反馈
作物产生或消耗的CO2取决于生长模型中的光合作用与呼吸作用,作物与温室空气交换的CO2的量为
式中,CO2、CH2O分别为CO2和同化物的摩尔质量。
3 温室番茄作物-环境互作模型
温室番茄作物-环境互作模型包含番茄生长与温室小气候环境两部分,两者之间存在相互作用,不可剥离。其中番茄作物生长模型(x)描述以温室内小气候为输入的作物动态生长过程,小气候模型()用于描述温室内部小气候受室外气候、温室结构、执行机构、作物生长等因素的影响而动态变化。模型结构可以通过微分方程表示,见形式表达式(26)和具体表达式(27)。
4 互作模型的使用和验证
本研究中首先利用上海市崇明温室的一年的番茄产量数据对秒时间尺度番茄生长模型进行了参数辨识确定。辨识后的生长模型结合到温室环境小气候模型中,再利用崇明温室的一段时间的实际气象与执行机构数据对加入了作物反馈的小气候模型进行参数辨识确定,由此得到崇明温室番茄作物-环境的互作模型。再利用崇明温室的其他年份的实际数据,对互作模型进行验证。
4.1 试验场景介绍
试验数据采集自上海崇明国家设施农业工程技术中心示范基地半封闭式Venlo型A8号温室(31°34'N,121°41'E),温室为东西朝向,长35 m,宽25 m,屋脊高8.25 m,屋檐高7.3 m,屋顶倾角22°,屋脊南北两侧各有3个长2.5 m宽1.2 m的天窗。温室配备内外遮阳网、保温幕、顶部补光系统、加热设备及循环通风系统,可以提高温室内部空气温湿度以及补充CO2的均匀性。温室配备荷兰的Priva自动控制系统,可以采集室内外的气候数据,室外气象站可以采集室外太阳辐射强度、室外温度、湿度、风速、降雨量等数据,室内传感器布置与作物冠层上方0.3 m,并随着作物的生长而向上移动,可以采集的室内数据包括室内温度、湿度、CO2浓度等数据。
温室内种植的作物为无限生长型番茄,番茄品种为Goriso,种植面积为680 m2,18个种植列,种植密度约为2.5株/m2。番茄作物种植于稻壳炭有机基质种植盒中,通过滴箭进行灌溉供水,并且定期供给营养液和氮肥。营养液的正常供给间隔为7~10 d,而在番茄根系吸收水分和养分能力较弱的定植前期,3~5 d浇施速效氮肥促进番茄植株营养生长。番茄植株定植于每年9月20日,经过100 d的生长期已开始稳定产出果实,在生长期间减去侧枝只留主茎干,定期修剪最下层叶片将叶面积指数维持在3.3~3.5 m2/m2,成熟的果实人工采摘称质量并记录。
4.2 模型评价指标
采用均方根误差(Root Mean Square Error,RMSE)、平均相对误差(Mean Relative Error,MRE)、平均绝对误差(Mean Absolute Error,MAE)作为参数辨识与模型精度验证的评价指标,它们的计算公式为
模型的求解和计算基于Matlab工具箱的ode45函数,该函数采用变步长四/五阶Runge-Kutta法对模型进行求解。
4.3 秒时间尺度生长模型验证

未知参数经辨识后的值见表3。
为了验证辨识后得到的番茄生长模型的有效性,利用2015—2018年的每一年定植日期(9月20日)起100d内的累积产量数据分别对辨识后的本文秒时间尺度生长模型进行验证,并与Integrated模型、TOMGRO模型进行对比。原始产量数据为人工采集并记录的果实鲜质量,每次采摘后记录从定植起至今已采收的累计果实鲜质量,前100d累积平均产量鲜质量约为4.1 kg/m2。由于不具备进行破坏性试验如烘干称量等条件,因此通过干鲜比来获取试验所需的干质量。图4中实际产量数据点为从定植起到采摘当日的累积干质量,由累积鲜质量换算而来,换算公式如下
式中DMF和FMF分别为所采摘果实的累积干质量和鲜质量,g;η为干鲜比,取值为6.3%[33]。模拟拟结果与实际产量值如图4,3个模型的模拟值与实测值间的误差见表4。
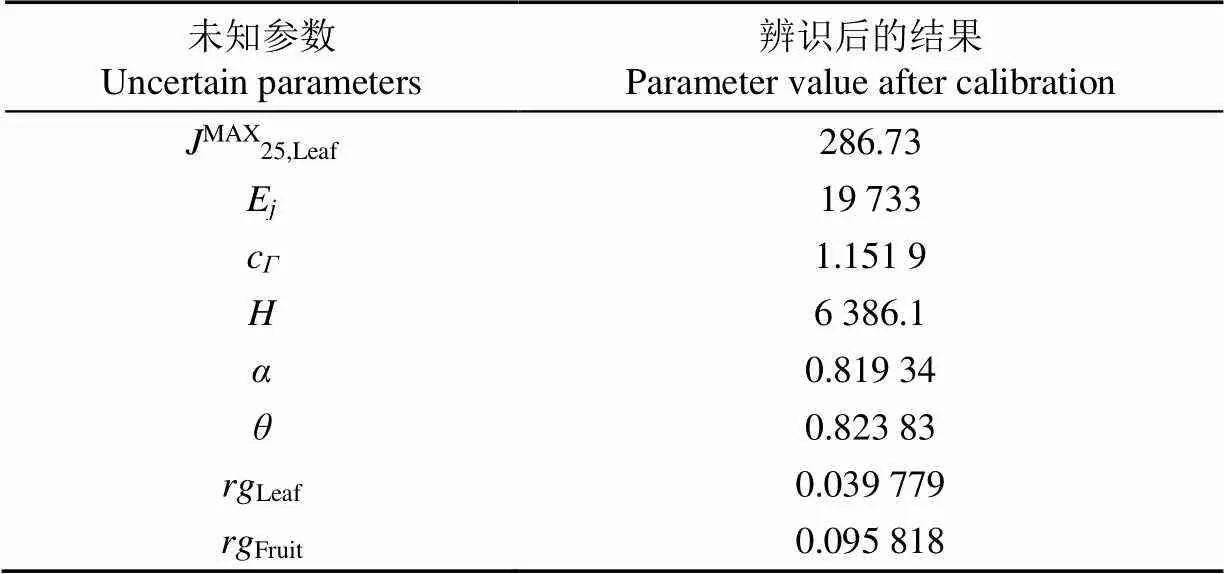
表3 生长模型未知参数辨识结果
从图表可以看出,利用2015—2018年的室内气候数据作为模型输入,秒时间尺度生长模型对作物产量的模拟与预测平均相对误差在5.8%~18%之间,RMSE在7.34~18.85 g/m2,准确性均优于TOMGRO模型与Integrated模型,可以更加准确地预测作物产量的动态增长。但由2015—2018年的对比图中可以看出模型预测结果总体高于实际产量值,且这种现象在作物生长的中前期(首次采收果实)更加显著,造成这种误差的原因经分析后可能在于:1)敏感性分析中呼吸模型中的系数被认为是不敏感参数未进行辨识,导致不能很好地模拟作物中前期的呼吸作用。2)干物质产量由实际产量乘以鲜质量到干质量的转换系数,未进行破坏性试验进行测量,导致产量值本身存在一定误差。3)因温室实际种植过程生产者栽培与管理导致的误差。

表4 3种模型仿真误差
4.4 小气候环境模型验证
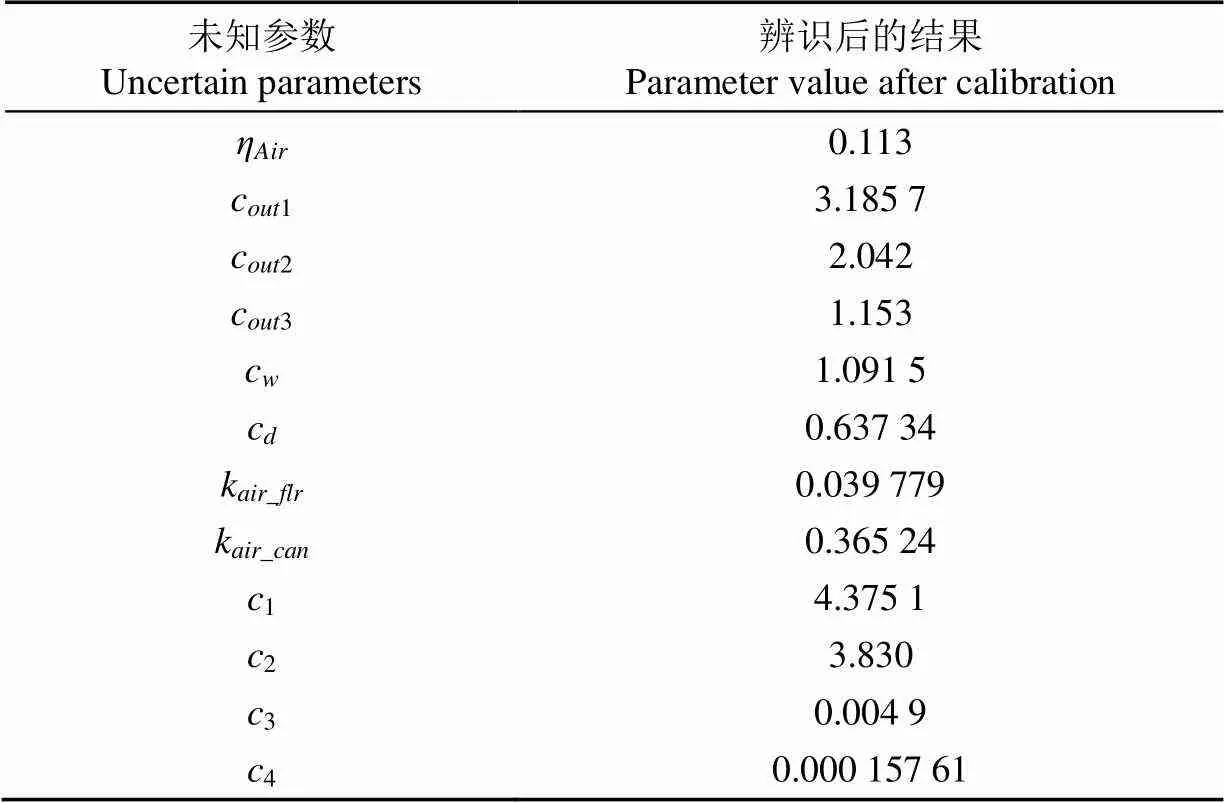
表5 小气候模型未知参数辨识结果
参数辨识后的小气候模型对辨识数据预测输出如图 5所示。该模型模拟温度动态的RMSE为0.68 ℃,模拟湿度动态的RMSE为0.84 g/m3,模拟CO2浓度动态的RMSE为23.47 mg/m3,平均相对误差分别为0.03、0.04、0.04,这说明该模型对3个环境因子的模拟效果都是比较好的。
互作模型是小气候模型与作物模型时间尺度统一之后模型,不仅能将室内环境的变化实时反映在作物上,也将作物对于环境的反馈机理考虑在内,如作物的光合、呼吸、蒸腾、生长等过程对环境的影响。在作物的不同生长阶段,其对环境的相互作用程度是不同的,在作物生长前期,叶面积和干物质量较小,对环境的影响较小几乎可以忽略,但在其生长后期,叶面积和干物质量较大,作物的多个生理过程,如光合、呼吸、蒸腾等,会严重影响温室内的环境动态。因此想要验证互作模型对作物对环境反馈这一复杂机理过程的模拟是否准确,需要在作物生长的全周期进行验证。因此本研究选取了番茄作物处于幼苗期(移栽后16d,LAI=0.1)、开花坐果期(移栽后51d,LAI=1)、结果期(移栽后79d,LAI=2)的3种情况,分别与未考虑作物反馈——叶面积指数与作物干物质量都为0的情况进行对比,来说明考虑作物对环境的实时反馈对于温室作物-环境系统建模的重要性,并验证模型在作物的整个生长周期中是否能准确模拟环境因子的动态变化。作物不同生长阶段的模型模拟结果如图6所示,模拟结果与测量值的误差如表6所示。
从图6与表6可以看出,建立的包含作物反馈小气候模型作物不同的生长阶段对温室内部环境因子动态变化的平均相对误差在3%~6%之间,可以很好地模拟环境因子动态变化,且模拟误差均小于未考虑作物对环境反馈的模型。以湿度与CO2浓度为例,误差随着作物的生长(LAI的增大)更加明显,这都说明了作物与环境的互相作用会影响模型准确性,在温室建模中这部分机理过程不可忽略。互作模型在作物不同的生长阶段均可较好地对温室内环境动态变化进行模拟,LAI=1和2时温度预测误差略大于LAI=0.1的幼苗期,分析原因在于LAI=1与LAI=2时由加热泵组成的加热系统已经开始为温室供热,温室中实际的加热升温过程是一个复杂的非均匀的过程,而在互作模型中,室内空气被视为均匀的,加热泵释放到室内的热能被空气均匀地吸收并升温;此外,因为加热过程的非均匀性,室内温度传感器与加热泵的相对位置也会影响温度测量值。
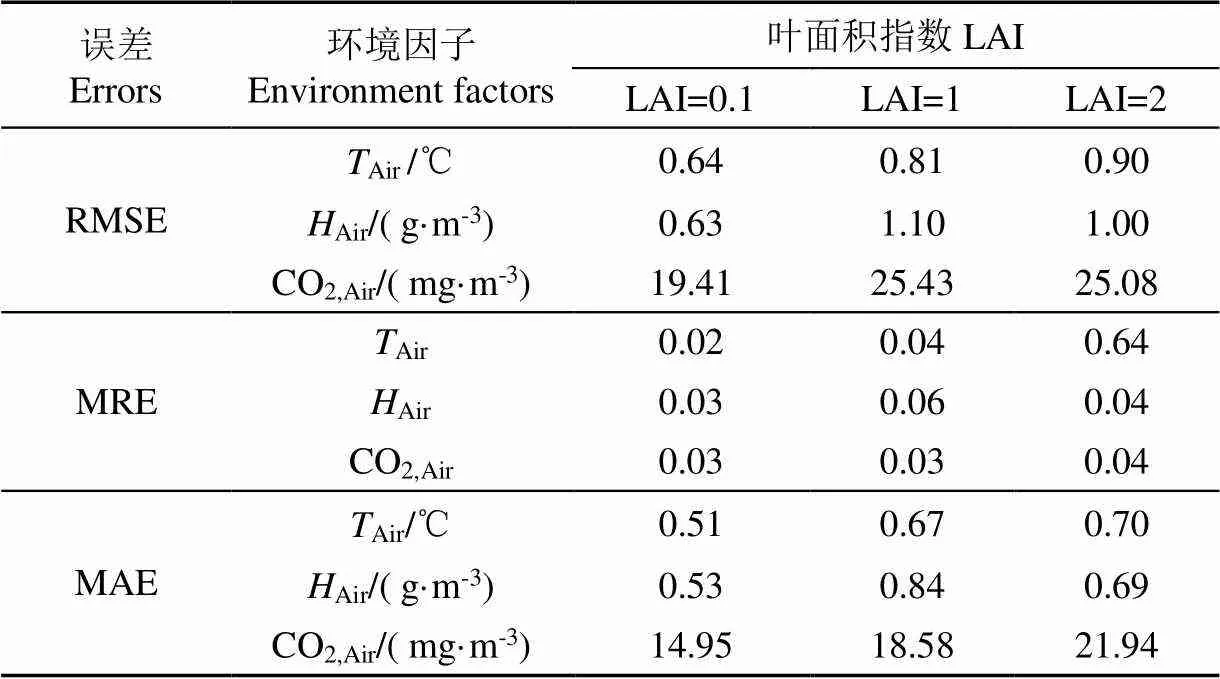
表6 不同生长阶段仿真误差
5 结 论
针对目前温室系统建模中存在的问题,本研究建立了时间尺度统一的温室番茄作物-环境互作模型,利用实际温室和产量数据对模型进行了参数辨识和验证,并与其他2个代表性模型进行了对比验证。
1)改变现有基于源库理论的天时间尺度作物生长模型的SUPPLY、DEMAND结构,建立由SUPPLY、PARTITION、GROWTH 3部分组成的作物生长模型,并通过模型替换、参数辨识等方法将3个子模型的时间尺度由天转化为秒,通过敏感性分析对模型不确定参数进行分类,得到秒时间尺度作物生长模型。
2)在秒时间尺度温室番茄生长模型的基础上,综合考虑作物对环境的反馈作用,建立温室番茄作物-环境互作模型。
3)利用上海市崇明A8温室2014年的实际番茄产量数据,通过贝叶斯优化方法对互作模型中的秒时间尺度生长模型进行参数辨识,并利用2015—2018年的数据进行模型验证。最终模拟结果表明秒时间尺度生长模型在2015—2018年产量模拟值与真实值的平均相对误差均在5.8%~18%之间,相较于Integrated模型与TOMGRO模型有更小的误差,2018年受不确定气候影响,3个模型的误差均偏大。
4)利用崇明温室连续5 d的实际运行数据对互作模型中加入了作物反馈的小气候环境模型进行参数辨识,并进行模型验证。在番茄作物3个不同生长阶段(幼苗期、开花坐果期、结果期),加入了作物反馈的小气候环境模型对温室内温度、湿度和CO2浓度动态变化的模拟平均相对误差均在3%~6%之间,相较于未考虑作物反馈的小气候模型有更小的误差。
互作模型的建立,将番茄作物生长模型和小气候环境模型统一为一个模型,其时间尺度也统一为秒。与以往温室模型不同的是,互作模型将小气候动态的作用直接通过作物生长反映,也将作物生长对小气候环境的实时反馈作用在小气候动态模型中反映。从而使得模型机理更加完善,因而模型也更加精准。它使得作物不同生长阶段模型对温室小气候环境的预测误差小于其他模型,从而保证模型更能够满足小气候环境控制的需要。互作模型中的未知参数需利用实际温室数据进行参数辨识后确定,以保证所得到的互作模型适合该地域该温室。未来可以在此温室互作模型基础上,进一步研究温室能耗模型与二氧化碳消耗等模型,从而研究基于作物产量和能耗优化的温室环境效益优化控制算法,提高温室栽培与控制管理的经济效益。
[1]紫琳. 我国主要温室设施面积已突破210万公顷未来将朝超低能耗方向发展[J]. 中国食品,2017(19):173
Zi Lin. The area of major greenhouse facilities in my country has exceeded 2. 1 million hectares and will develop towards ultra-low energy consumption in the future[J]. China Food, 2017(19): 173. (in Chinese with English abstract)
[2]贺冬仙. 国内植物工厂发展的思考[J]. 农业工程技术,2016(19):24-25.
[3]徐立鸿,苏远平,梁毓明. 面向控制的温室系统小气候环境模型要求与现状[J]. 农业工程学报,2013,29(19):1-15.
Xu Lihong, Su Yuanping, Liang Shuming. Requirement and current situation of control-oriented microclimate environmental model in greenhouse system[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2013, 29(19): 1-15. (in Chinese with English abstract)
[4]Straten G V, Challa H, Buwalda F. Towards user accepted optimal control of greenhouse climate[J]. Computers & Electronics in Agriculture, 2000, 26(3): 221-238.
[5]杜尚丰,陈俐均,徐丹,等. 温室生产系统优化控制方法综述[J]. 河北工业大学学报,2017,46(5):85-93.
Du Shangfeng, Chen Lijun, Xu Dan, et al. Survey of optimal control on greenhouse cultivation system[J]. Journal of Hebei University of Technology, 2017, 46(5): 85-93. (in Chinese with English abstract)
[6]Albright L D, Arvanitis K G, Drysdale A E. Environmental control for plants on earth and in space[J]. Control Systems IEEE, 2001, 21(5): 28-47.
[7]Gijzen H, Heuvelink E, Challa H, et al. Hortism: A model for greenhouse crops and greenhouse climate[J]. Acta Horticulturae, 1998(456): 441-450.
[8]Jones J W, Dayan E, Allen L H, et al. A dynamic tomato growth and yield model (TOMGRO)[J]. Transactions of the ASAE, 1991, 34(2): 663-672.
[9]Gijzen H, Vegter J G, Nederhoff E M. Simulation of greenhouse crop photosynthesis: validation with cucumber, sweet pepper and tomato[C]//IV International Symposium on CO2in Protected Cultivation. 1989: 71-80.
[10]Lin D, Wei R, Xu L. An integrated yield prediction model for greenhouse tomato[J]. Agronomy, 2019, 9(12): 873.
[11]倪纪恒,罗卫红,李永秀,等. 温室番茄发育模拟模型研究[J]. 中国农业科学,2005,38(6):1219-1225.
Ni Jiheng, Luo Weihong, Li Yongxiu, et al. Simulation of the development of tomato in greenhouse[J]. Scientia Agricultura Sinica, 2005, 38(6): 1219-1225. (in Chinese with English abstract)
[12]Vanthoor B H E, Stanghellini C, Henten E J V, et al. A methodology for model-based greenhouse design: Part 1, a greenhouse climate model for a broad range of designs and climates[J]. Biosystems Engineering, 2011, 110(4): 363–377.
[13]胥芳,张立彬,陈教料,等. 玻璃温室小气候温湿度动态模型的建立与仿真[J]. 农业机械学报,2005,11:108-111.
Xu Fang, Zhang Libin, Chen Jiaoliao, et al. Modeling and simulation of subtropical greenhouse microclimate in China[J]. Transactions of the Chinese Society for Agricultural Machinery, 2005, 11: 108-111.
[14]Dong Q, Yang W, Yang L, et al. Crop model-based greenhouse optimal control system: Survey and perspectives[C]. //Computer and Computing Technologies in Agriculture VI, 2013, 216-224.
[15]Schmidt M, Reinisch K, Puta H, et al. Determining climate strategies for greenhouse cucumber production by means of optimization[J]. IFAC Proceedings Volumes, 1987, 20(5): 345-350.
[16]邓璐娟,张侃谕,龚幼民,等. 温室环境多级控制系统及优化目标值设定的初步研究[J]. 农业工程学报,2005,21(5):119-122.
Deng Lujuan, Zhang Kanyu, Gong Youmin, et al. Preliminary study on hierarchical greenhouse environment control system and setting of the optimized target values[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2005, 21(5): 119-122. (in Chinese with English abstract)
[17]Henten E J V, Bontsema J. Time-scale decomposition of an optimal control problem in greenhouse climate management[J]. Control Engineering Practice, 2009, 17(1): 88-96.
[18]徐丹. 双时间尺度温室生产最优控制算法研究[D]. 北京:中国农业大学,2018.
Xu Dan. Two Time-scale Optimal Control of Greenhouse Cultivation[D]. Beijing: China Agricultural University, 2018. (in Chinese with English abstract)
[19]Marcelis L F M, Heuvelink E, Goudriaan J. Modelling biomass production and yield of horticultural crops: A review[J]. Scientia Horticulturae, 1998, 74(1): 83-111.
[20]Farquhar G D, von Caemmerer S, Berry J A. A biochemical model of photosynthetic CO2assimilation in leaves of C3species[J]. Planta, 1980, 149(1): 78-90.
[21]邢会敏,相诗尧,徐新刚,等. 基于EFAST方法的AquaCrop作物模型参数全局敏感性分析[J]. 中国农业科学,2017,50(1):64-76.
Xing Huimin, Xiang Shiyao, Xu Xingang, et al. Global sensitivity analysis of AquaCrop model parameters based on EFAST method[J]. Scientia Agricultura Sinica, 2017, 50(1): 64-76. (in Chinese with English abstract)
[22]张静潇,苏伟. 基于EFAST方法的CERES-Wheat作物模型参数敏感性分析[J]. 中国农业大学学报,2012,17(5):149-154.
Zhang Jingxiao, Su Wei. Sensitivity analysis of CERES-Wheat model parameters based on EFAST method[J]. Journal of China Agricultural University, 2012, 17(5): 149-154. (in Chinese with English abstract)
[23]Dejonge K C, Ii J C A, Ahmadi M, et al. Global sensitivity and uncertainty analysis of a dynamic agroecosystem model under different irrigation treatments[J]. Ecological Modelling, 2012, 231: 113-125.
[24]Stanghellini C. Transpiration of greenhouse crops: an aid to climate management[D]. Netherlands: Wageningen University, 1987.
[25]Su Yuanping, Xu Lihong. Towards discrete time model for greenhouse climate control[J]. Engineering in Agriculture, Environment and Food, 2017, 10(2): 157-170.
[26]Chen J, Yang J, Zhao J, et al. Energy demand forecasting of the greenhouses using nonlinear models based on model optimized prediction method[J]. Neurocomputing, 2016, 174: 1087-1100.
[27]苏远平. 温室环境控制与鲁棒控制[D]. 上海:同济大学,2015.
Su Yuanping, Greenhouse Climate Control and Robust Control[D]. Shanghai: Tongji University, 2015. (in Chinese with English abstract)
[28]Shen Y, Wei R, Xu L. Energy consumption prediction of a greenhouse and optimization of daily average temperature[J]. Energies, 2018, 11(1): 65.
[29]Graamans L, van den Dobbelsteen A, Meinen E, et al. Plant factories: Crop transpiration and energy balance[J]. Agricultural Systems, 2017, 153: 138-147.
[30]Impron I, Hemming S, Bot G P A. Simple greenhouse climate model as a design tool for greenhouses in tropical lowland[J]. Biosystems Engineering, 2007, 98(1): 79-89.
[31]Kittas C, Boulard T, Papadakis G. Naturl ventilation of a greenhouse with ridge and side openings: Sensitivity to temperature and wind effects[J]. Transactions of the ASAE, 1997, 40(2): 415-425.
[32]崔佳旭,杨博. 贝叶斯优化方法和应用综述[J]. 软件学报,2018,29(10):3068-3090.
Cui Jiaxu, Yang Bo. Survey on bayesian optimization methodology and applications[J]. Journal of Software, 2018, 29(10): 3068-3090. (in Chinese with English abstract)
[33]应建阳. 基于能耗与作物产量预测的温室经济效益研究[D]. 杭州:浙江工业大学,2018.
Ying Jianyang. Research on Economic Benefits of Greenhouse Based on Prediction of Energy Consumption and Crop Yield[D]. Hangzhou: Zhejiang University of Technology, 2018. (in Chinese with English abstract)
Development and verification of tomato crop-environment interaction model in second timescale greenhouse
Xu Lihong, Meng Fanzheng, Wei Ruihua
(,,201800,)
Greenhouse cultivation has a strong impact of crops on the complex process, because various time scales existed in the controlled environment. The greenhouse model can be divided into two types, including the crop growth and microclimate model. The crop growth model was usually used to simulate daily change of crop, where the variables of crop is updated at each time step within a day. The microclimate model has a shorter calculating step, because the climate in a greenhouse changed quickly, due mainly to rapid fluctuation of weather outside. In general, the climate physics is considered as a fast process, while the crop physics is considered as a slow one. The difference of time scales has brought a great challenge at the level of crop state, such as the rapid fluctuations of greenhouse climate or the elusive ambient inputs in the monitoring system of a greenhouse. In this study, taking tomato as a research object, a crop-climate interactive model at small timescale was established to balance the time scales of crop and climate in greenhouse. Firstly, the growth model was divided into three sub-modules, including the SUPPLY, PARTITION, and GROWTH. The replacement, structural transformation were implemented in the model, when three modules were transformed from a long timescale (day level) to a small timescale (second level). Two types of uncertain parameters were divided in the model under a global sensitivity analysis (Extended Fourier Amplitude Sensitivity Test, EFAST), such as sensitive and insensitive parameters. Insensitive parameters were fixed in the model, whereas, the sensitive parameters needed to be identified, according to real production data in specific greenhouses. Secondly, the general interactive model was obtained to combine small time-scale crop growth model and greenhouse microclimate model. Microclimate in the interactive model was different from other microclimate model, because it fully considered the reaction between the microclimate and crop, where the microclimate model was be considered as an input for the crop model. The proposed interactive model was also calibrated and validated in the field test. The real data was collected from A8 Venlo type greenhouse at Chongming Island, Shanghai of China. The 4-year (2015-2018) observed data of tomato yield was used in the model. It was found that the root mean square error (RMSE) between the simulated and real yield value was 7.3-18.85, and the average relative error was between 5.8% and 18%, both less than TOMGRO and Integrated model. The data demonstrated that the interactive model presented a better performance on the yield prediction of tomato. The microclimate simulation result also proved that the interactive model behaved a higher accuracy at different crop growth stages than that without considering the influence of crop growth. The average relative error was less than 10% for the prediction of microclimate environment at three stages of crop growth (growing, blooming and setting fruiting), indicating high efficiency to simulate the real dynamics of greenhouse microclimate. Nevertheless, there were relatively larger deviations in the small part of simulation from actual data, such as simulated yield in 2018 and temperature trajectory when LAI=2. Bayesian optimization was also used to identify the uncertain parameters in both crop growth and microclimate model. Model structure and parameters were totally determined after sensitivity analysis and parameter identification. Consequently, the interactive model can provide a theoretical basis for cultivation and environmental control in a greenhouse.
greenhouse; models; tomato growth model; microclimate model; sensitivity analysis; Bayesian optimization
徐立鸿,孟凡峥,蔚瑞华. 秒尺度温室番茄作物-环境互作模型构建与验证[J]. 农业工程学报,2021,37(8):212-222.doi:10.11975/j.issn.1002-6819.2021.08.024 http://www.tcsae.org
Xu Lihong, Meng Fanzheng, Wei Ruihua. Development and verification of tomato crop-environment interaction model in second timescale greenhouse[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2021, 37(8): 212-222. (in Chinese with English abstract) doi:10.11975/j.issn.1002-6819.2021.08.024 http://www.tcsae.org
2020-11-18
2021-01-23
上海市科技兴农重点攻关项目(沪农科创字(2018)第3-2号);国家自然科学基金项目(项目号:61973337)
徐立鸿,教授,博士生导师,研究方向为温室环境建模与控制、预测控制和智能控制。Email:xulihong@tongji.edu.cn
10.11975/j.issn.1002-6819.2021.08.024
S626
A
1002-6819(2021)-08-0212-11
猜你喜欢
商丘师范学院学报(2023年9期)2023-09-06 06:03:42
农业灾害研究(2022年1期)2022-05-07 01:31:04
农业工程技术(2022年1期)2022-04-19 13:58:20
力学学报(2021年10期)2021-12-02 02:32:04
云南农业(2021年9期)2021-09-24 11:57:06
云南农业(2021年8期)2021-09-06 11:36:44
能源工程(2021年1期)2021-04-13 02:06:12
英语文摘(2019年2期)2019-03-30 01:48:28
湖南林业科技(2017年1期)2017-02-06 05:29:18
水利技术监督(2016年6期)2017-01-15 14:01:30
