
一类食饵具有Allee 效应的捕食-食饵模型的分歧解
2021-06-19 07:54李艳玲
工程数学学报 2021年3期
曹 倩, 李艳玲
(1- 长安大学理学院,西安 710064; 2- 陕西师范大学数学与统计学院,西安 710119)
1 引言
Allee 效应指的是种群密度低时,种群单位增长率和种群密度之间正相关,该效应是当前反应扩散模型研究中的热点问题.近年来,许多实验表明物种存在双Allee 效应,例如,一种濒临灭绝的物种—岛屿灰狐,它们原产于美国加利福尼亚州海峡群岛的六个岛屿.金雕的捕食是幼年和成年岛屿灰狐的主要威胁.此外,它们在物种密度较低的情况下,雌性繁殖的可能性降低,这是由于雌性怀孕前缺乏配偶以及很难找到新的配偶.因此,岛屿灰狐的生长速度受到双Allee 效应的影响(繁殖困难带来的Allee 效应和捕食驱动的Allee 效应).在海洋生态系统中也存在双Allee 效应[1].然而,形成双Allee 效应的机制有很多,双Allee 效应对生物多样性保护很重要并且给系统带来复杂的动力学行为,因此研究双Allee 效应是十分必要的.本文研究如下食饵带有双Allee 效应的捕食-食饵模型
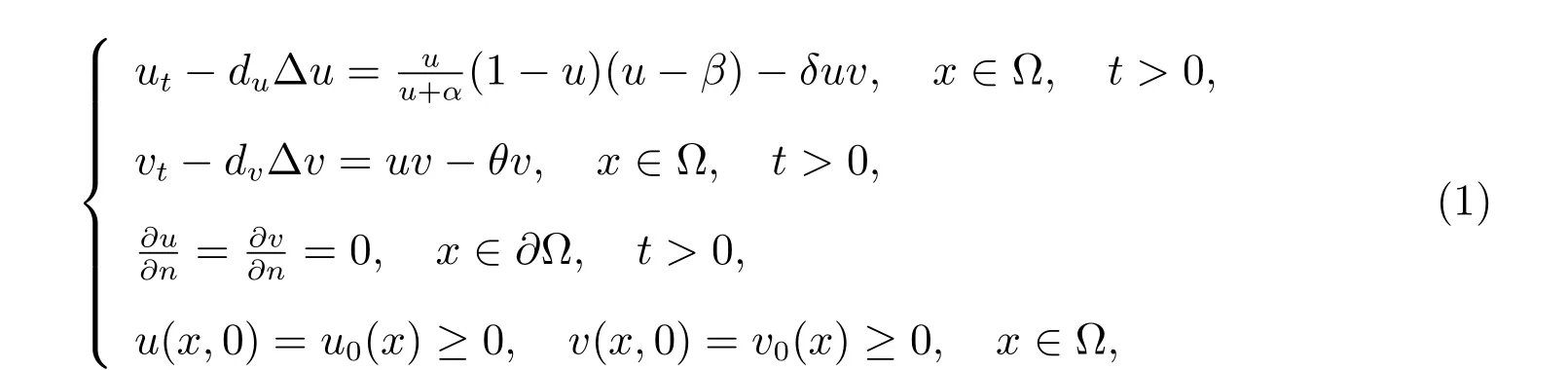

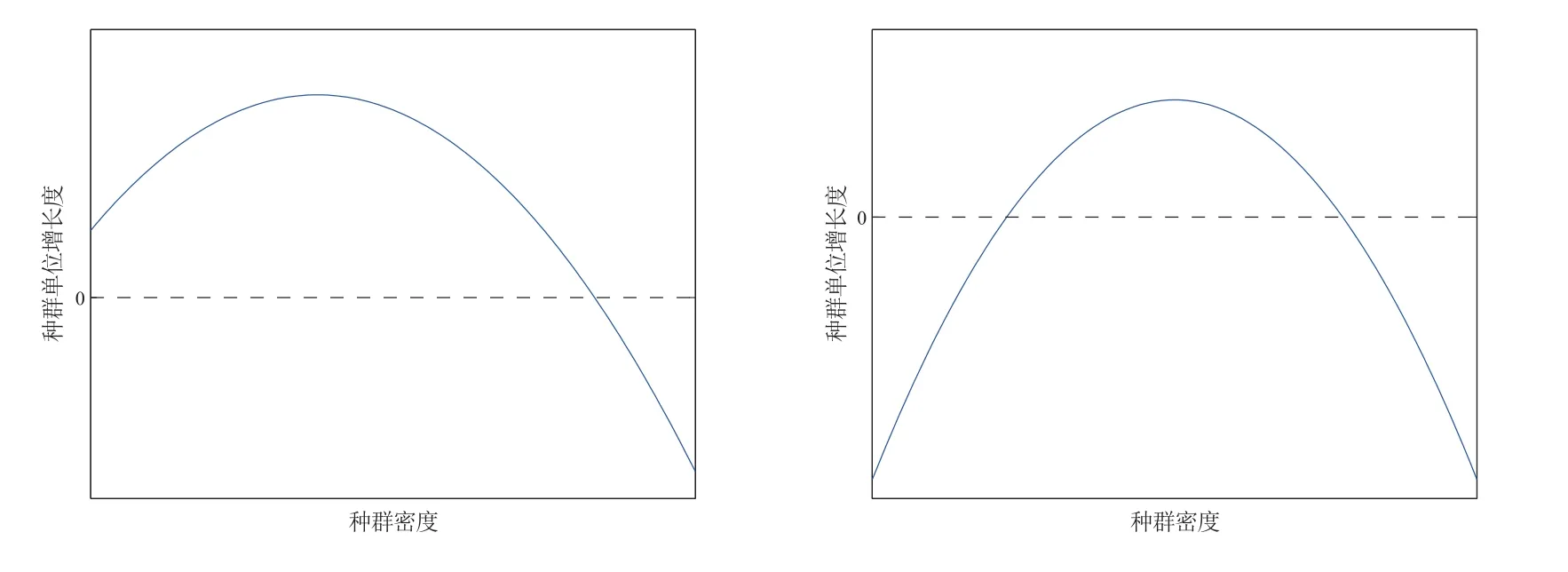
图1 弱Allee 效应和强Allee 效应定义的图示
近几十年来,很多学者已经对捕食-食饵型模型进行了广泛的研究[2–14],其中文献[2]研究了在齐次Dirichlet 边界条件下一类具有强Allee 效应的捕食-食饵模型共存解的存在性.文献[3]研究了一类具有脉冲扰动的随机捕食-食饵系统的周期解和平稳分布.文献[4,5]分别研究了带有加法Allee 效应和乘法Allee 效应的捕食-食饵模型共存解的惟一性和多解性.然而在齐次Neumann 边界条件下带有双Allee 效应的扩散捕食-食饵模型的研究很少见.因此,本文主要研究系统(1)所对应平衡态系统的分歧解的存在性.下面考虑系统(1)相应的平衡态系统
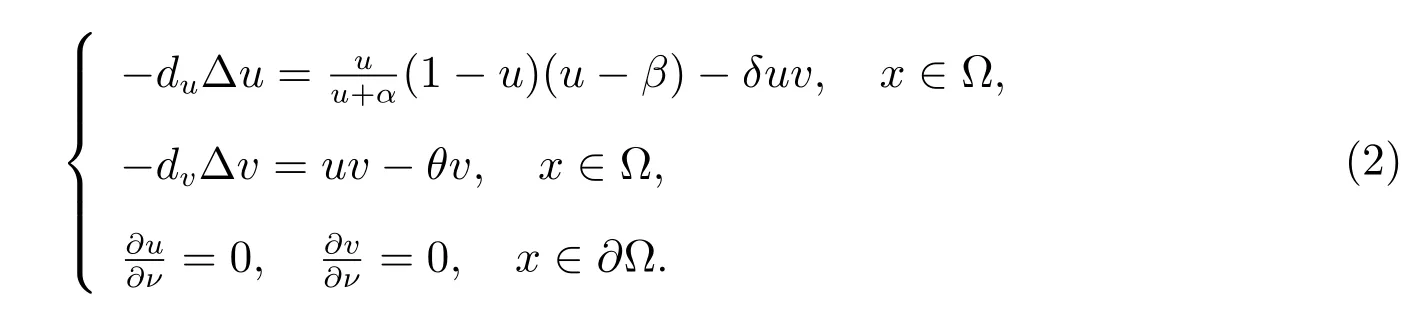
2 稳定性分析
本节利用稳定性理论分别研究强Allee 效应和弱Allee 效应情形下系统(2)常数解的稳定性.这对后面分歧解的研究有着重要作用.
首先,我们研究强Allee 效应(0<β <1)的情形.此时,系统(2)有四个常数解:平凡解(0,0),半平凡解(1,0)和(β,0),唯一的正常数解(θ,vθ),其中
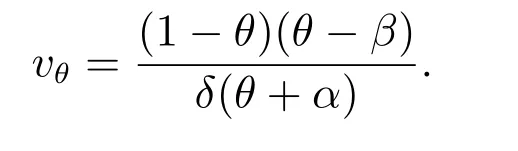
当且仅当β <θ <1 时,正常数解(θ,vθ)存在.
为了记号简单,我们记u=(u,v)和
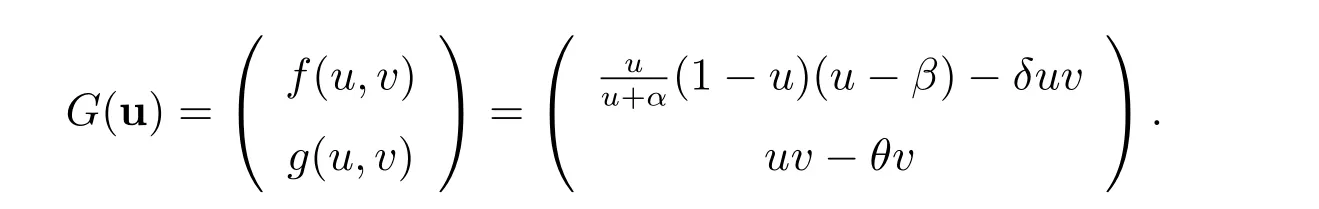
下面我们得到G(u)在u*=(u*,v*)处的线性化
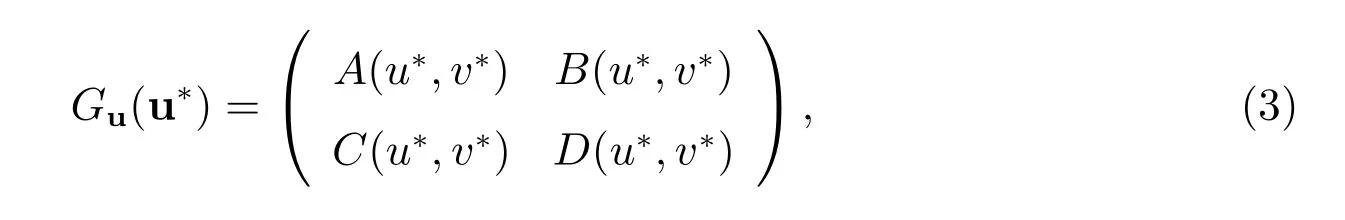
其中
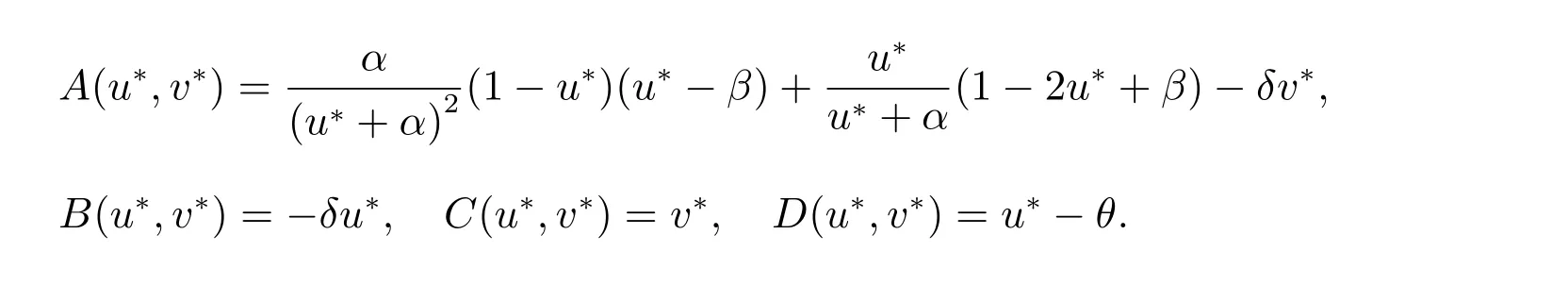
假设{λn}是-Δ 在齐次Neumann 边界条件下的一列可列特征值,满足0 =λ0<λ1<λ2<··· <λj <···.设mj是λj(j= 0,1,2,···)的代数重数并且φjk(1≤k ≤mj)为λj对应的标准化特征函数.下面定义
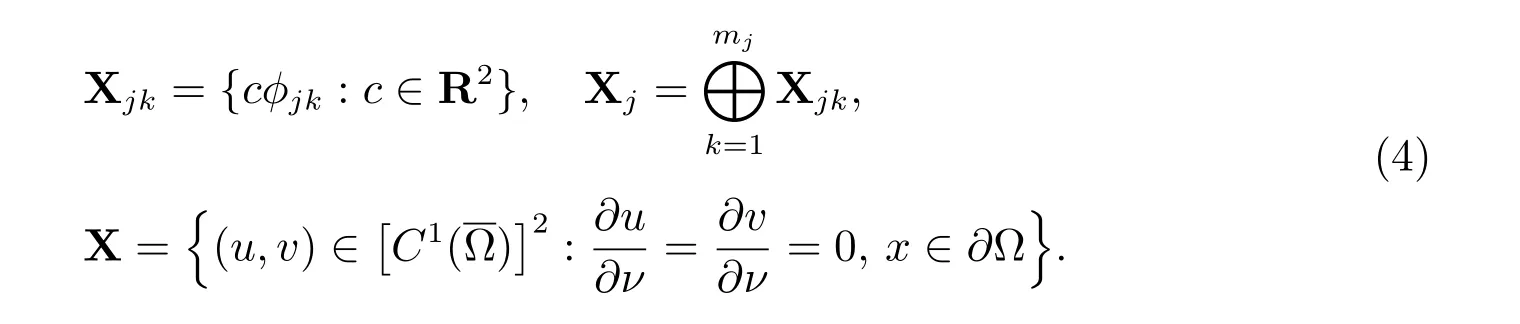
容易得到
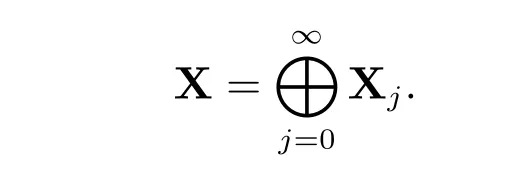
下面将系统(2)在u*处线性化
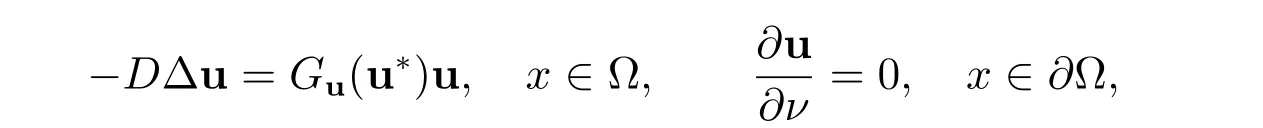
其中
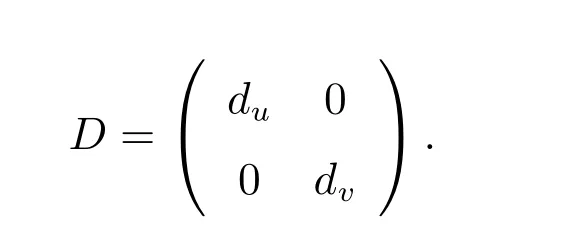
下面记
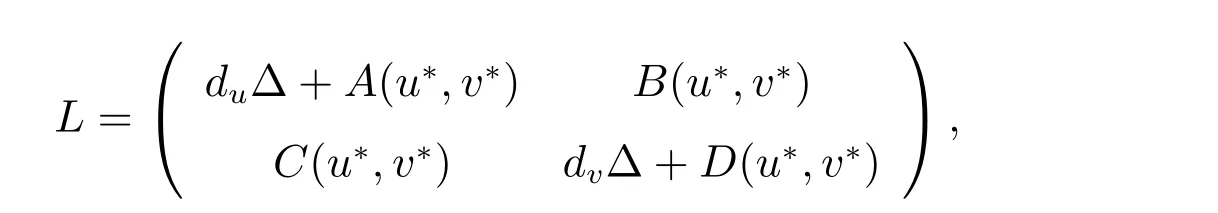
其中A(u*,v*), B(u*,v*), C(u*,v*)和D(u*,v*)如上所述.
假设(Φ(x),Ψ(x))是L算子相应于特征值μ的特征函数.那么有
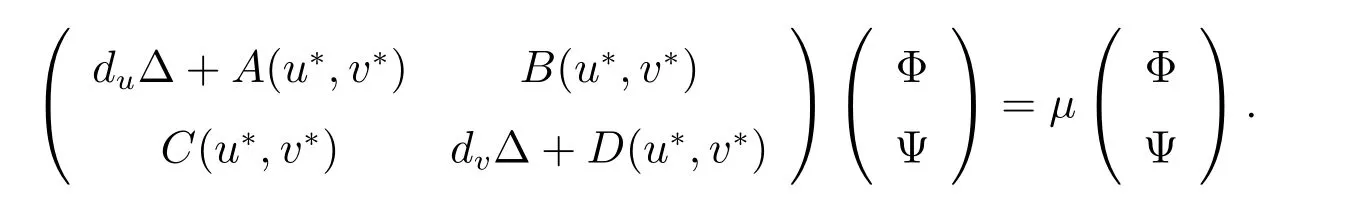
令
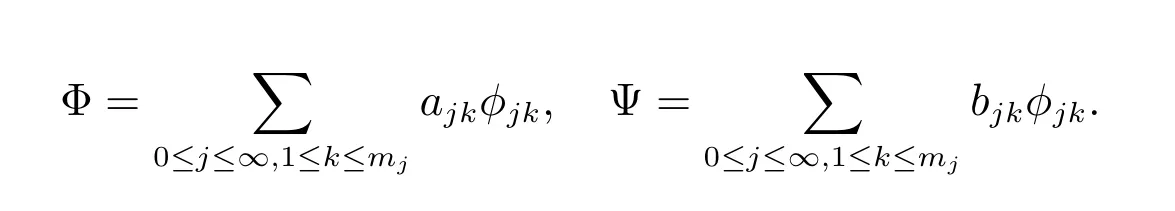
则可得到
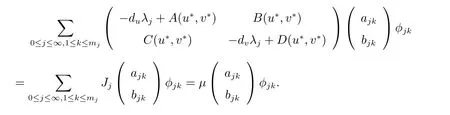
因此,我们知道当且仅当存在某个j ≥0,使得矩阵μI-Jj的行列式等于0,即
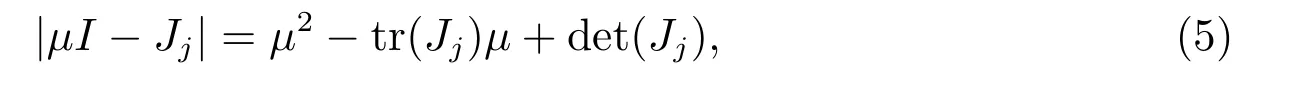
其中

因此,类似于文献[15],我们有以下结论:
(i)
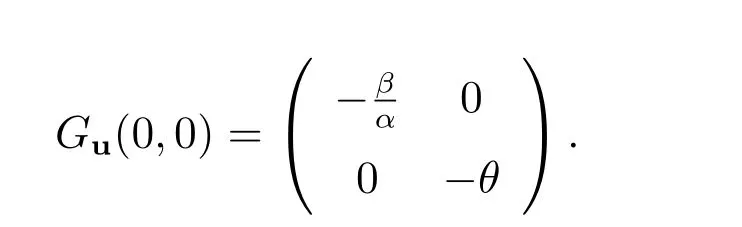
显而易见,(0,0)局部渐近稳定.
(ii)
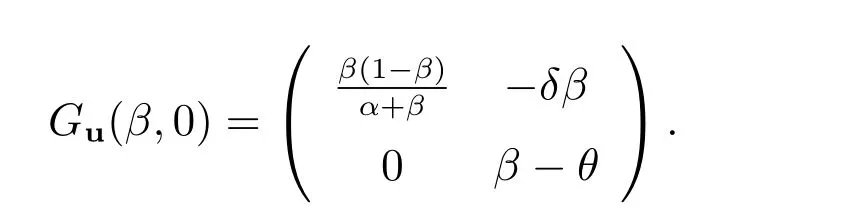
显而易见,(β,0)不稳定.
(iii)
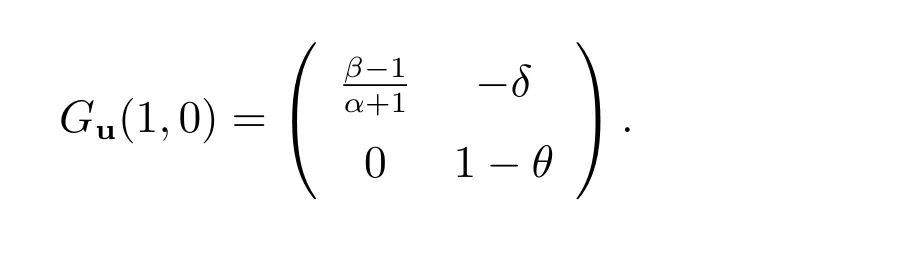
当θ >1 时,我们得到tr(Ji)<0 和det(Ji)>0(i ≥0).因此,(1,0)局部渐近稳定;
当0<θ <1 时,我们得到
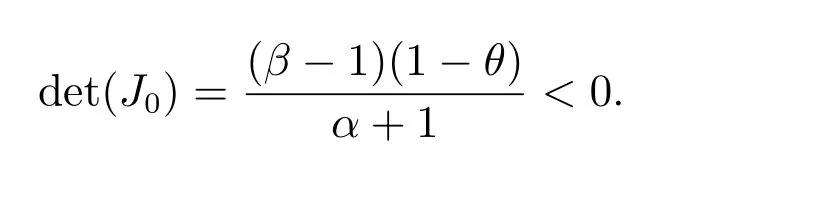
所以(1,0)不稳定.
(iv)
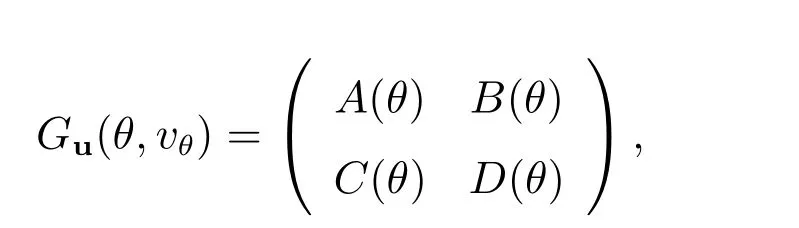
其中
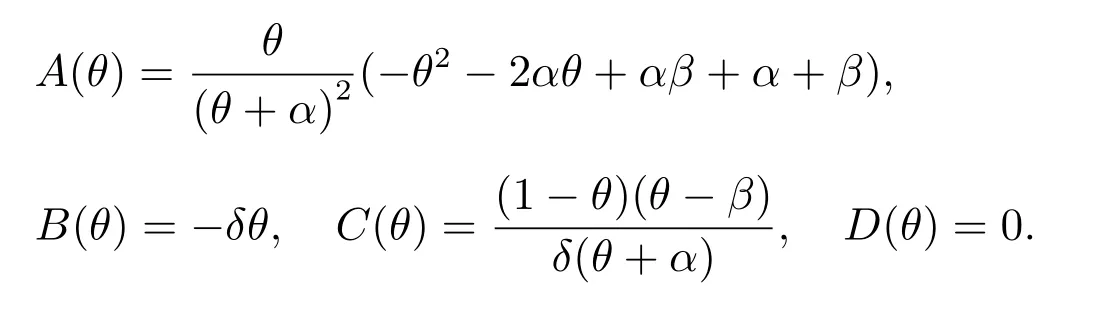
令
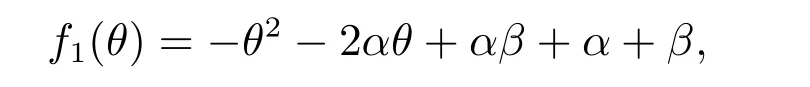
那么A(θ) = 0 等价于f1(θ) = 0.由f1(θ)的表达式知,当θ ∈(-α,+∞)时,f1(θ)单调递减,所以f1(θ)在θ ∈(β,1)⊂(-α,+∞)时单调递减.此外
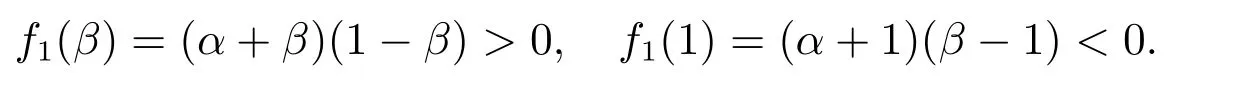
那么存在唯一的
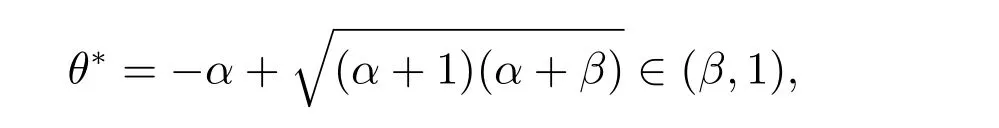
使得
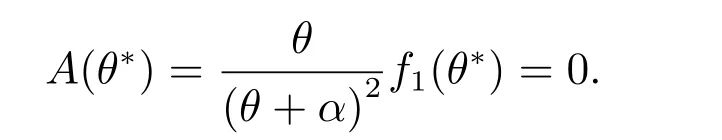
(a) 当β <θ*<θ <1 时,我们得到tr(Ji)<0 和det(Ji)>0(i ≥0).因此(θ,vθ)局部渐近稳定.
(b) 当β <θ <θ*<1 时,我们得到A(θ)>0.对于i=0,我们有tr(J0)>0,这说明(5)至少有一个有正实部的根.因此(θ,vθ)不稳定.
综上可得下面的定理,给出强Allee 效应(0<β <1)情形下常数平衡解(0,0),(β,0),(1,0)和(θ,vθ)的稳定性.
定理1 假设du, dv, α, δ >0,0<β <1.那么有下列结论:
(i) 当θ >0 时,(0,0)局部渐近稳定;
(ii) 当θ >0 时,(β,0)不稳定;
(iii) 当θ >1 时,(1,0)局部渐近稳定;当0<θ <1 时,(1,0)不稳定;
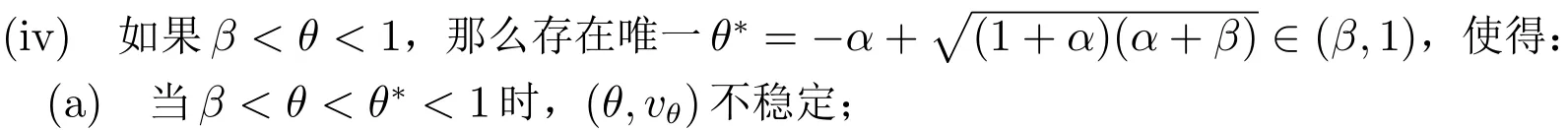
(b) 当β <θ*<θ <1 时,(θ,vθ)局部渐近稳定.
接下来,我们考虑弱Allee 效应(-1<β <0)的情形,此时系统(2)有平凡解(0,0),半平凡解(1,0)和唯一正常数解(θ,vθ),其中当且仅当0<λ <1 时,(θ,vθ)存在.下面我们研究这些常数解的稳定性,下述稳定性定理证明与定理1 类似,此处定理证明省略.
定理2 假设du, dv, α, δ >0,-1<β <0.那么下述结论成立:
(i) 当θ >0 时,(0,0)不稳定;
(ii) 当0<θ <1 时,(1,0)不稳定;当θ >1 时,(1,0)局部渐近稳定;
(iii) 如果

那么(θ,vθ)局部渐近稳定.
3 分歧解的存在性
这一节,我们分别对强Allee 效应(0<β <1)和弱Allee 效应(-1<β <0)的分歧解进行分析.我们以du为分歧参数,并利用局部分歧理论研究正常数解(θ,vθ)邻域上局部分歧解的结构和性质.当研究Ω=(0,l)时,系统(2)对应的如下椭圆系统
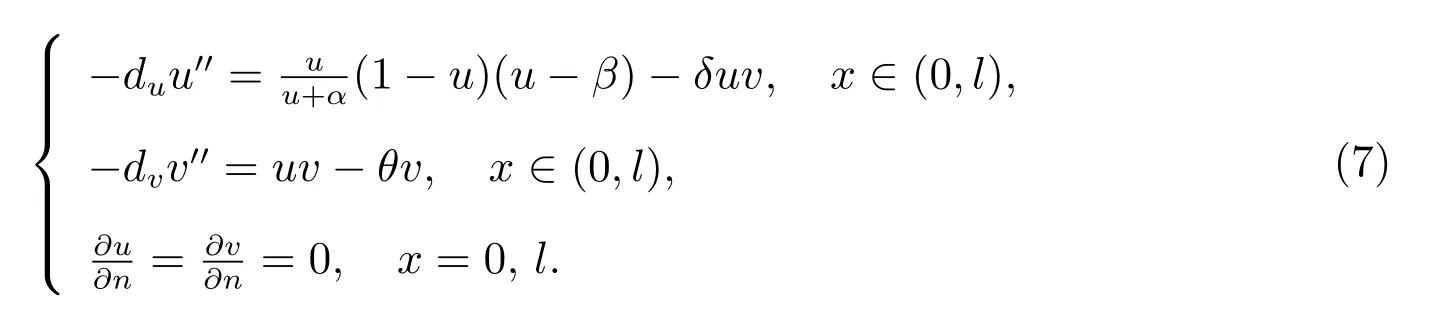
下面,我们考虑特征值问题
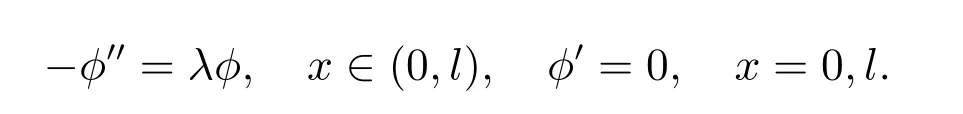
这个特征值问题有一系列简单特征值
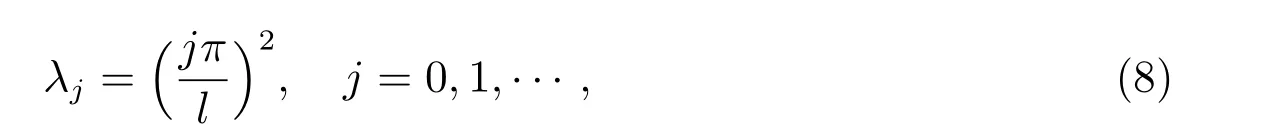
和相应标准化特征函数
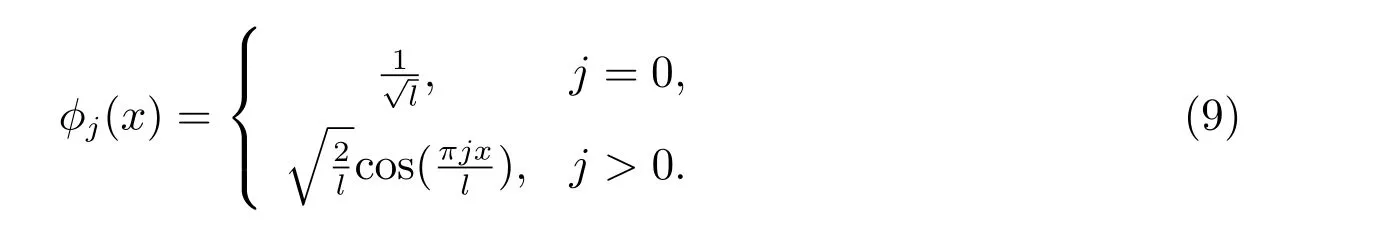
显然,这些特征函数构成L2(0,l)上一组正交基.

并把S看做具有C2(Ω)范数的Banach 空间.易知系统(7)的解是映射ˆF: (0,∞)×S →Y的零点,所以
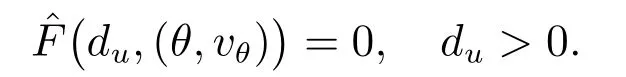
对某个τ >0,如果ˆF(τ,(θ,vθ))在(0,∞)×S上的邻域包含不同于(du,(θ,vθ))(du >0)的零点,那么(τ,(θ,vθ))为方程ˆF= 0 的分歧点.根据文献[16]的定理1.7,如果下述条件成立,则(τ,(θ,vθ))为方程ˆF=0 的分歧点.

综上,我们得到下述强Allee 效应(0<β <1)情形下的局部分歧结论.
定理3 假设
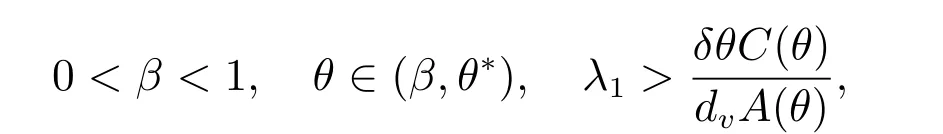
和当k/=j时,dj/=dk,那么(dj,(θ,vθ))是ˆF= 0 的一个分歧点.并且,当|s|足够小时,系统(2)有一个单参数的非常数解Γj(s)=(du(s),u(s),v(s)),其中和du(0)=dj.此外,du(s), u(s), v(s)是关于s连续可微的函数.
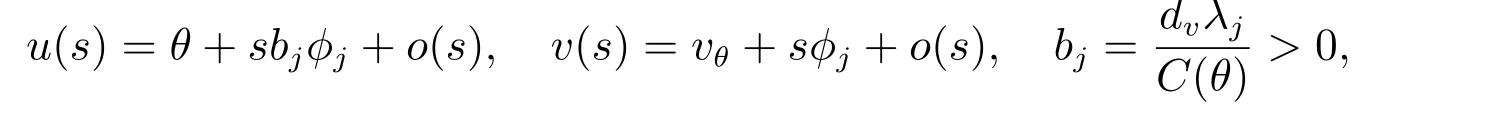
证明 考虑ˆF(du,u)关于u 在正常数解(θ,vθ)处的Fr´echet 导数及ˆFu(du,(θ,vθ))的伴随算子

其中
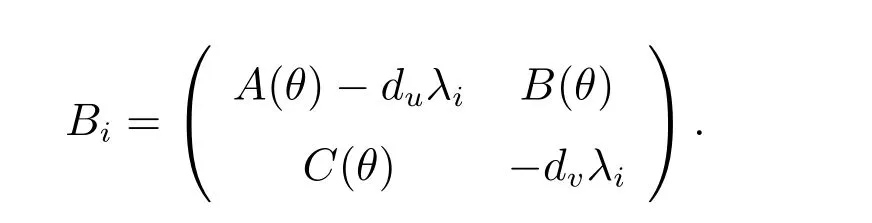
由此可得
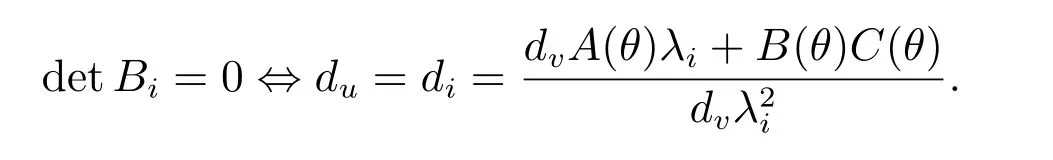
取du=dj,则有

所以,我们得到
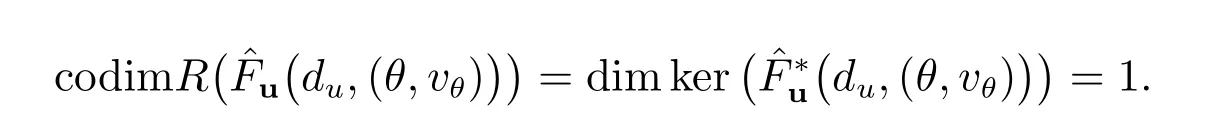
因此条件(b)成立.
最后,因为
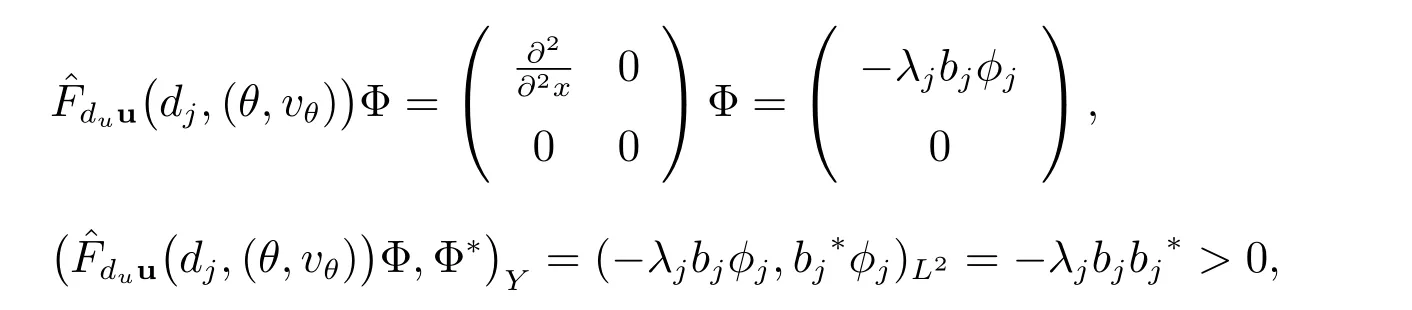
我们得到
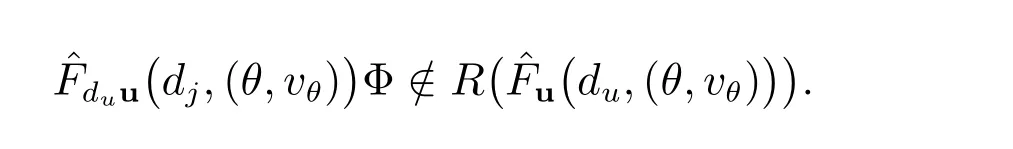
所以条件(c)成立.
类似于定理3 的证明方法,我们可以证明弱Allee 效应(-1<β <0)情形下的局部分歧结论.此处证明过程省略.
定理4 假设
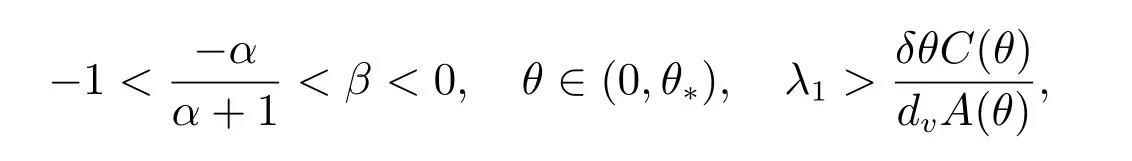
和当k/=j时,dj/=dk,那么(dj,(θ,vθ))是ˆF= 0 的一个分歧点.并且,当|s|足够小时,系统(2)有一个单参数的非常数解Γj(s)=(du(s),u(s),v(s)),其中
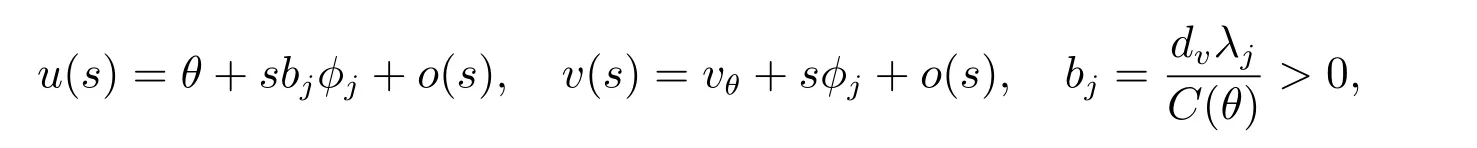
和du(0)=dj.此外,du(s), u(s), v(s)是关于s连续可微的函数.
4 数值模拟
本节利用Matlab 软件进行数值模拟,用以验证前面的理论分析.这里研究强Allee 效应(0<β <1)的情况.弱Allee 效应情形可以用类似的方法研究.
图2 中的参数选取α=1, β=0.2, δ=3, θ=0.4, l=3, t=2000,此时
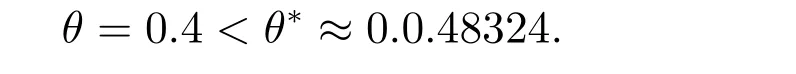
图3 中的参数选取α=1, β=0.2, δ=3, θ=0.8, l=3, t=2000.此时
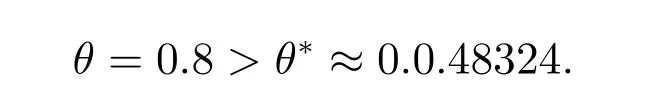
图2 和图3 验证了定理1 和定理3:强Allee 情形下,存在一个临界值θ*,使得当θ ∈(θ*,1)时,(θ,vθ)局部渐近稳定;而当θ ∈(β,θ*)时,(θ,vθ)不稳定,此时系统(2)至少存在一个非常数正解.
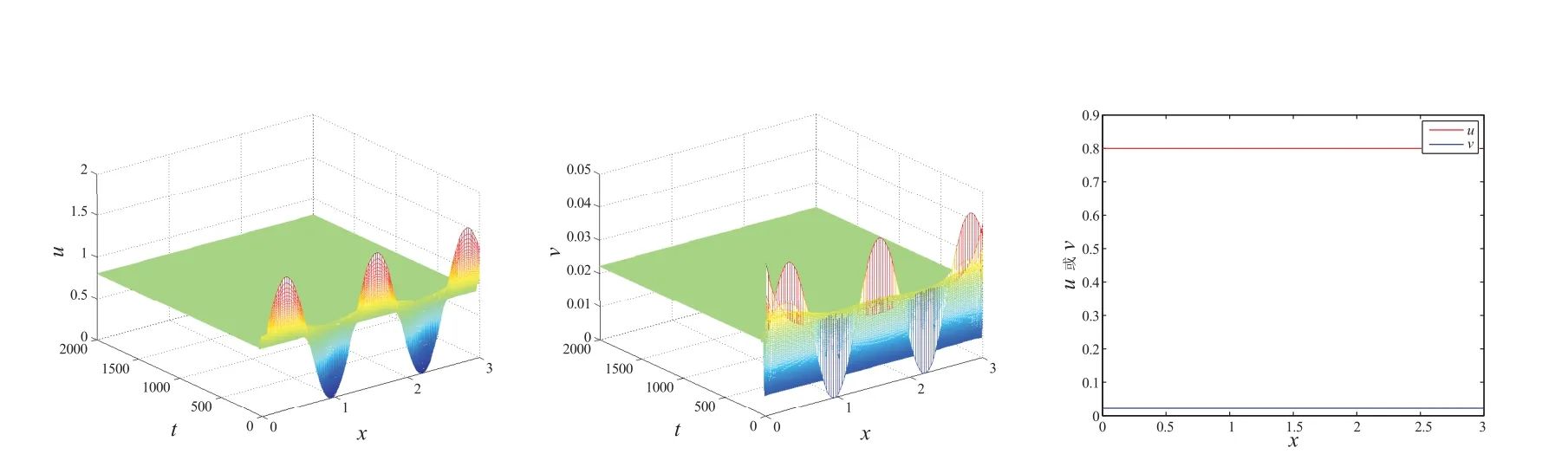
图3 λ >λ*时,系统(1)共存解(u,v)的时空图和系统(1)在t=2000 时,共存解(u,v)的空间分布
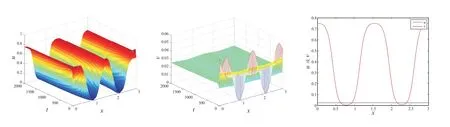
图2 λ <λ*时,系统(1)共存解(u,v)的时空图和系统(1)在t=2000 时,共存解(u,v)的空间分布
5 结语
本文主要运用稳定性理论和局部分歧理论研究了一类食饵具有双Allee 效应的捕食-食饵模型,得到了该模型常数解的稳定性、分歧解的存在性和分歧解的性质.结果表明,当
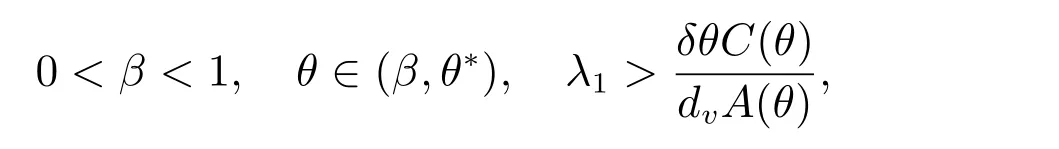
和当k/=j时,dj/=dk,那么(dj,(θ,vθ))是ˆF=0 的一个分歧点;当
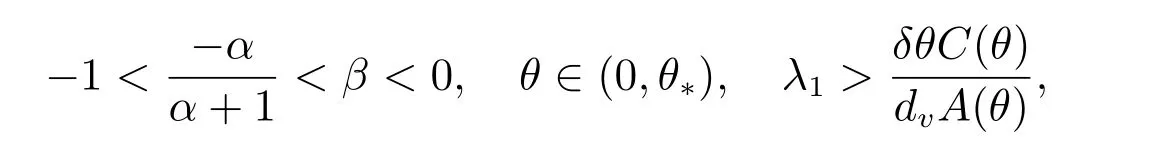
和当k/=j时,dj/=dk,那么(dj,(θ,vθ))是ˆF=0 的一个分歧点.
猜你喜欢
中等数学(2022年6期)2022-08-29
数学物理学报(2022年1期)2022-03-16
中学生数理化·七年级数学人教版(2022年11期)2022-02-22
数学物理学报(2021年2期)2021-06-09
应用数学(2020年4期)2020-12-28
数学物理学报(2019年4期)2019-10-10
校园英语·上旬(2019年6期)2019-10-09
数学物理学报(2019年3期)2019-07-23
中学生数理化·七年级数学人教版(2017年6期)2017-11-09
发明与创新(2016年38期)2016-08-22
