
miR-210通过NF-κB信号通路影响宫颈癌Hela细胞恶性生物学行为
2021-06-09 06:29罗茹蓉张桂芸姚文香雒钰花吴梅仙白银市第二人民医院妇科甘肃白银730900
中国老年学杂志 2021年11期
罗茹蓉 张桂芸 姚文香 雒钰花 吴梅仙 (白银市第二人民医院妇科,甘肃 白银 730900)
宫颈癌(CC)是世界上最常见的妇科恶性肿瘤之一,也是女性癌症相关死亡率的主要原因〔1〕。在过去几十年中,随着CC组织的病理检查的改善和早期诊断的应用,CC的发病率和死亡率显著降低,但晚期出现转移的宫颈癌患者5年生存率仅5%~15%〔2〕。目前,CC的主要治疗方法是手术、化疗和放疗,尽管目前人们在治疗和预防方面付出了巨大努力,但患者预后仍不能令人满意。因此,充分了解CC的发生和发展的机制,对识别和筛选CC早期诊断和临床治疗的分子标志物具有重要意义。微小RNA(miRNA)是一类内源性小的非编码RNA,长度18~22个核苷酸,其与相关靶mRNA的3′-UTR相互作用,通过直接和间接影响mRNA的转录过程来调节相关基因的表达。近年来,据报道miRNA在一些参与肿瘤发生的事件中起重要作用,如细胞存活、增殖、凋亡、侵袭和转移〔3,4〕。研究表明,miR-210在人类不同的肿瘤中发挥致癌的作用,如结肠直肠癌、乳腺癌和膀胱癌等〔5~7〕。据报道,miR-210能够促进前列腺癌细胞上皮间质转化和骨转移,该过程与核因子(NF)-κB信号通路密切相关〔8〕。然而,miR-210在CC细胞恶性生物学行为中的作用仍然未知。本实验通过检测miR-210在CC细胞中的表达,探究其对Hela细胞恶性生物学行为的影响及可能的机制,以期为CC的早期诊断和靶向治疗提供一定的实验基础。
1 材料与方法
1.1材料 人正常宫颈上皮细胞H8和CC细胞Hela、caski、siha和c4-1均购自美国ATCC细胞库;胰蛋白酶和DMEM培养基均购自美国HyClone公司;胎牛血清购自美国Gibco公司;青霉素、链霉素购自北京鼎国昌盛生物技术有限公司;miR-210 inhibitor及inhibitor Control购自广州市锐博生物科技有限公司;TRIzol试剂和Lipofectamine 2000试剂购自美国Invitogen公司;逆转录试剂盒、荧光定量聚合酶链式反应(PCR)测定试剂盒和AnnexinⅤ-FITC/PI细胞凋亡测定试剂盒购自日本TaKaRa公司;噻唑蓝(MTT)试剂和二甲基亚砜均购自美国Sigma公司;Transwell小室购自美国Coring公司;细胞裂解液、二喹啉甲酸(BCA)蛋白浓度测定试剂盒、电化学发光(ECL)试剂购自碧云天生物科技研究所;B细胞淋巴瘤(Bcl)-2、Bcl-2相关x蛋白(Bax)和酶切半胱氨酸天冬氨酸蛋白酶(Caspase)-3、IκBα、p-IκBα、NF-κB和p-NF-κB抗体购自美国CST公司;辣根过氧化物酶(HRP)标记的IgG购自北京中杉金桥生物技术有限公司。
1.2细胞培养和转染 人宫颈癌Hela细胞培养在含10%胎牛血清的RPMI1640培养基中,在培养基中分别加入终浓度为1×105U/L的青霉素和100 mg/L的链霉素,将细胞放置在含5%CO2、相对湿度95%、37℃培养箱中培养,细胞贴壁生长密度达80%以上时,用0.25%的胰蛋白酶消化传代。将对数生长期的Hela细胞接种到6孔板中,每孔接种2×105个细胞,置培养箱常规培养,细胞达50%~60%融合时,根据Lipofectamine 2000试剂说明书转染miR-210 inhibitor或inhibitor Control,将细胞分为空白对照(Control)组、阴性对照(anti-NC)组和实验(anti-miR-210)组,各组Hela细胞置37℃培养箱继续培养。
1.3qRT-PCR检测细胞中miR-210的表达水平 分别收集处于对数生长期的H8、Hela、caski、siha、c4-1细胞和转染48 h后各组Hela细胞,TRIzol法提取各细胞中总RNA,采用逆转录试剂盒合成cDNA。以cDNA为模板,使用荧光定量PCR试剂盒进行扩增,反应条件为:94℃ 3 min,94℃ 15 s,56℃ 20 s,72℃ 10 s,共40个循环。以U6为内参,采用相对定量2-△△Ct法计算miR-210相对表达水平。
1.4CCK-8法检测Hela细胞增殖情况 对数期的Hela细胞接种到96孔板中,细胞接种密度为5×103个/孔,待细胞贴壁生长后按照1.2进行转染,每组细胞设置3个复孔,在转染48 h时,向每孔细胞中缓慢加入10 μl CCK-8溶液,避免气泡产生,在37℃培养箱孵育4 h,使用酶标仪测定波长为450 nm处光密度值(OD值),实验重复3次。
1.5Transwell实验检测细胞侵袭和迁移能力 Hela细胞转染48 h后,用不含血清的DMEM培养液饥饿处理16 h,分别收集各组细胞,调整细胞密度为1×106个/ml,每个Transwell小室的上室(侵袭实验Transwell小室的上室预先用Matrigel胶包被,迁移实验Transwell小室的上室不以Matrigel胶包被)加入100 μl细胞悬液,在下室加入500 μl含10%胎牛血清的DMEM培养液,置37℃培养箱继续培养48 h。取出Transwell小室,用90%乙醇固定20 min,用0.1%结晶紫染色20 min,用棉签擦去上室未穿膜的细胞,磷酸盐缓冲液(PBS)洗涤3次,风干,在显微镜低倍镜(×100)下随机选取5个视野进行观察,并记录细胞数,实验重复3次。
1.6流式细胞仪检测细胞凋亡情况 分别收集转染48 h后各组Hela细胞,用预冷PBS洗涤细胞2次,离心收集约1×106个细胞,加入结合缓冲液100 μl重悬细胞,向细胞悬液中加入FITC-AnnexinⅤ溶液5 μl,轻轻混匀,孵育15 min,再向细胞中加入PI染液5 μl,混匀后避光反应15 min,使用上流式细胞仪检测各组Hela细胞凋亡情况,实验重复3次。
1.7Western印迹检测蛋白表达水平 转染48 h后收集各组Hela细胞,加入适量细胞裂解液,置冰上提取细胞总蛋白。BCA法测定蛋白样品浓度,取30 μg蛋白样品行10%十二烷基硫酸钠-聚丙烯酰胺凝胶电泳,蛋白分离后电转至PVDF膜上,在含5%脱脂奶粉的封闭液中封阻1 h,分别加入一抗4℃过夜杂交,Bcl-2一抗(1∶500),Bax一抗(1∶500),酶切Caspase-3一抗(1∶500),IκBα一抗(1∶500),p-IκBα一抗(1∶500),NF-κB一抗(1∶500),p-NF-κB一抗(1∶500)。PBST洗涤3次(每次10 min),加入HRP标记的二抗(1∶2 000),室温杂交2 h。PBST洗涤3次(每次10 min),以ECL试剂显影,以β-actin标定,采用Image J软件分析条带灰度值。
1.8统计学方法 使用SPSS21.0软件进行单因素方差分析及SNK-q检验。
2 结 果
2.1miR-210在CC细胞中表达水平升高 qRT-PCR检测结果显示,Hela细胞(3.52±0.38)、caski细胞(2.44±0.25)、siha细胞(2.59±0.28)和c4-1细胞中miR-210的表达水平(1.94±0.20)显著高于正常宫颈上皮H8细胞(1.00±0.10,P<0.05)。在Hela细胞中的表达水平最高,后续采用Hela细胞开展功能实验。
2.2miR-210 inhibitor转染效率检测 anti-miR-210组Hela细胞中miR-210表达水平(0.31±0.03)明显低于Control组(1.00±0.11)和anti-NC组(1.00±0.09,P<0.05)。Control组和anti-NC组细胞中miR-210表达水平比较,差异无统计学意义(P>0.05)。
2.3敲减miR-210抑制Hela细胞增殖、侵袭和迁移能力 CCK-8实验结果显示,anti-miR-210组Hela细胞OD值明显低于Control组和anti-NC组(P<0.05)。Control组和anti-NC组细胞OD值比较,差异无统计学意义(P>0.05)。Transwell实验结果显示,anti-miR-210组侵袭和迁移细胞数明显低于Control组和anti-NC组(P<0.05)。Control组和anti-NC组侵袭和迁移细胞数相比,差异无统计学意义(P>0.05)。见图1和表1。

图1 敲减miR-210对Hela细胞侵袭和迁移能力的影响(结晶紫染色,×200)

表1 敲减miR-210对Hela细胞增殖、侵袭和迁移能力的影响
2.4敲减miR-210促进Hela细胞凋亡 流式细胞仪检测结果显示,anti-miR-210组Hela细胞凋亡率明显高于Control组和anti-NC组(P<0.05)。Control组和anti-NC组细胞凋亡率比较,差异无统计学意义(P>0.05)。Western印迹检测凋亡相关蛋白结果显示,anti-miR-210组Hela细胞中Bax和酶切Caspase-3蛋白表达水平明显高于Control组和anti-NC组(P<0.05),Bcl-2蛋白表达水平明显低于Control组和anti-NC组(P<0.05)。Control组和anti-NC组细胞中Bcl-2、Bax和酶切Caspase-3蛋白表达水平比较,差异无统计学意义(P>0.05)。见图2,表2和图3。

图2 流式细胞仪检测各组Hela细胞凋亡率
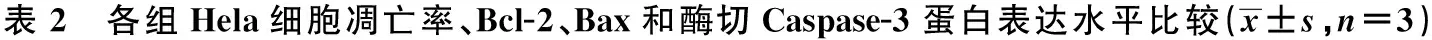
组别凋亡率(%)Bcl-2Bax酶切Caspase-3Control组5.31±0.680.76±0.080.11±0.010.04±0.01anti-NC组5.42±0.660.74±0.070.10±0.010.03±0.01anti-miR-210组19.26±2.111)2)0.30±0.031)2)0.52±0.061)2)0.68±0.071)2)F/P值108.267/0.00049.869/0.000136.026/0.000244.765/0.000

图3 Western印迹检测各组Hela细胞中Bcl-2、Bax和酶切Caspase-3蛋白表达水平
2.5敲减miR-210抑制NF-κB信号通路的激活 Western印迹检测结果显示,anti-miR-210组Hela细胞中p-IκBα和p-NF-κB蛋白表达水平明显低于Control组和anti-NC组(P<0.05),IκBα和NF-κB蛋白表达水平无明显变化(P>0.05);Control组和anti-NC组细胞中IκBα、p-IκBα、NF-κB和p-NF-κB蛋白表达水平比较,差异无统计学意义(P>0.05)。见图4和表3。

图4 Western印迹检测各组Hela细胞中IκBα、p-IκBα、NF-κB和p-NF-κB蛋白水平
表3 各组Hela细胞中IκBα、p-IκBα、NF-κB和p-NF-κB蛋白水平比较
3 讨 论
研究发现,miRNA表达异常不仅能够调控肿瘤细胞无限增殖还可影响细胞侵袭和转移〔9,10〕。因此,深入探究miRNA的生物学功能为恶性肿瘤的早期诊断和靶向治疗提供新的方向。以往研究显示,miR-210在人类多种肿瘤中呈高表达,作为致癌基因,miR-210能够靶向调控基因表达或信号通路的活化。如Liu等〔11〕研究发现,miR-210在骨肉瘤组织中的表达显著高于其配对正常组织,体外功能实验结果显示,miR-210可通过直接靶向成纤维细胞生长因子受体样(FGFRL)1促进骨肉瘤细胞迁移和侵袭。Luan等〔12〕研究显示,miR-210通过靶向Bcl-2腺病毒E1B相互作用蛋白(BNIP)3保护肾上腺嗜铬细胞瘤PC-12细胞免受缺氧诱导的损伤,此外该过程涉及PI3K/AKT/mTOR信号通路。阳宁等〔13〕研究发现,miR-210在前列腺癌组织中表达水平显著高于癌旁组织,其可通过下调BNIP3的表达抑制前列腺癌PC-3细胞凋亡。并且miR-210的表达升高与肾癌、乳腺癌、多发性骨髓瘤等肿瘤淋巴结转移、TNM分期和预后不良有关〔14~16〕。目前研究发现,miR-210在宫颈癌组织中异常高表达,然而关于miR-210在宫颈癌中的功能及作用机制尚不清楚〔17〕。
有研究指出,miR-210作为致癌基因可通过上调Bax、Caspase-3和Caspase-9蛋白表达水平促进胶质瘤U87细胞的凋亡〔18〕。另一项研究表明,miR-210可通过调控Bcl-2和Bax蛋白表达水平影响卵巢癌OVCAR3和SKOV3细胞凋亡〔19〕。本研究结果显示,敲减miR-210后细胞中Bax和酶切Caspase-3蛋白表达水平明显上调,Bcl-2蛋白表达水平明显下调,这与以往研究一致。提示敲减miR-210可能通过调控凋亡相关蛋白表达水平促进细胞凋亡。研究表明,NF-κB信号通路在人类多种类型的癌症中异常激活,这与肿瘤的进展和转移显著相关。例如,在CC中,NF-κB信号的异常激活与CC的预后不良密切相关;抑制NF-κB信号通路能够抑制CC细胞增殖、侵袭和迁移,并促进细胞凋亡〔20~22〕。在静息的细胞中,NF-κB和IκB结合形成复合体,以无活性形式存在于细胞质中,当细胞受外界刺激后,IκB发生磷酸化暴露NF-κB核定位位点,NF-κB转位至细胞核,进而调控相关基因转录及表达〔23〕。本研究结果提示过表达miR-210可能抑制NF-κB信号通路的激活。Zhang等〔24〕研究发现,miR-210通过靶向死亡受体(DR)6抑制NF-κB信号通路影响软骨细胞活力和细胞凋亡。结合Zhang等〔24〕和Ren等〔8〕研究提示miR-210可能通过调控NF-κB信号通路发挥作用。本研究结果表明miR-210可能通过调控NF-κB信号通路影响CC Hela细胞增殖、凋亡、侵袭和迁移。
总之,本研究结果表明miR-210在CC细胞中呈高表达,敲减CC Hela细胞中miR-210的表达能够通过抑制NF-κB信号通路的激活抑制细胞增殖、侵袭和迁移,诱导细胞凋亡。提示miR-210可能有望成为CC靶向治疗的新靶点。
猜你喜欢
保健医苑(2022年5期)2022-06-10
食品安全导刊(2021年21期)2021-08-30
中国临床医学影像杂志(2021年6期)2021-08-14
肝博士(2020年5期)2021-01-18
天津医科大学学报(2019年6期)2019-08-13
现代检验医学杂志(2016年2期)2016-11-14
中国病理生理杂志(2015年8期)2015-12-21
医学研究杂志(2015年7期)2015-06-22
医学研究杂志(2015年3期)2015-06-10
创业家(2015年1期)2015-02-27
