
不同发酵模式下郫县豆瓣后发酵过程动力学模型的建立与分析
2021-06-07 08:24赵晓燕吴晓霞林洪斌车振明丁文武
中国食品学报 2021年5期
谢 思,赵晓燕,吴晓霞,谢 磊,林洪斌,吉 礼,吴 韬,车振明,丁文武*
(1 西华大学食品与生物工程学院 成都610039 2 四川省食品生产安全协会 成都610056)
郫县豆瓣属中国传统发酵食品,以其味辣香醇、黏稠绒实、红棕油亮、酱香浓郁等特点在我国酱类产品中独树一帜[1],是川菜制作过程中不可或缺的一种调味品,对郫县豆瓣进行相关研究具有非常重要的意义。目前,国内许多研究机构尤其是川内的各大科研单位已对郫县豆瓣进行了大量的研究工作,其研究内容主要包括发酵过程中风味物质的形成以及影响因素[2-3],后发酵过程中菌落的变化和相关微生物分析[4-7]以及有害物质如黄曲霉毒素的变化与关键控制点分析[8]等,这些研究工作有力地促进了郫县当地豆瓣产业的健康、快速发展。
然而,目前郫县豆瓣的后熟发酵大多采用传统的“翻晒露”工艺生产,即将物料置于陶缸或条形池中,于露天环境下,白天利用太阳光进行热晒升温发酵,夜间则低温发酵,无论白天、黑夜,有雨则加盖防雨,无雨则敞开露晒,同时每天早、晚各翻搅1 次以保证物料均匀反应。该工艺生产方式落后,机械自动化控制水平低,生产效率低下,导致生产成本增加,食品安全卫生控制难度增大,生产规模难以扩大,产品质量标准统一困难。随着我国劳动力成本的增加和科技的飞速发展,工业生产已逐渐进入智能时代。根据郫县豆瓣传统生产工艺的特点,设计新型现代化生产工艺和设备,提高自动化生产水平,促进产业升级已成为郫县豆瓣生产行业的迫切需求。
工业生产过程的自动控制离不开动力学模型的构建,且动力学模型在中小工艺放大过程中具有非常重要的作用[9]。国内外学者对生产过程中动力学模型的构建进行了大量研究,在食品生产方面涉及鱼类[10-12]、蔬菜[13-15]、水果[16-18]等方面的研究,并且都取得了较好的研究成果。然而,针对郫县豆瓣在发酵过程中的动力学模型尚未见研究报道。本研究通过构建郫县豆瓣半封闭恒温与传统自然后熟发酵过程动力学模型,探讨两者的关系,以期为郫县豆瓣的现代化生产工艺的设计和改进提供参考。
1 材料与方法
1.1 材料
甜瓣子与椒醅均购于四川郫县豆瓣股份有限公司,购回后立即按一定比例混合,分别置于自然与恒温发酵装置中发酵。
1.2 试剂
氢氧化钠、亚硫酸钠、酒石酸钾钠、3,5-二硝基水杨酸、葡萄糖、甲醛、酚酞、乙醇、邻苯二甲酸氢钾、丙酮、三氯乙酸、盐酸、17 种氨基酸标样、茚三酮,以上试剂均为分析纯级,购于成都市科龙化工厂。
1.3 仪器与设备
30+氨基酸自动分析仪,英国Biochrom 公司;SpectraMax i3x 酶标仪,美国Molecular Devices,LLC 公司;BS110S 型电子天平,福州科锡仪器设备有限公司;7200 可见光分光光度计,上海尤尼柯仪器有限公司;磁力搅拌器,常州越新仪器制造有限公司。
1.4 郫县豆瓣自然与恒温发酵
郫县豆瓣发酵的试验装置如图1所示。整套装置由空气过滤装置、自动控温装置、恒温发酵装置、测温装置、自然发酵装置组成。自动控温装置是一种采用机械制冷降温、电加热升温的液体循环设备,本次试验中加入循环液体水对恒温发酵装置进行控温。恒温发酵装置是一个50 L 带夹套的玻璃发酵罐,用纱布将封头上的管口封住,形成一个半封闭的发酵环境。自然发酵装置是豆瓣厂常用的晒缸。测温装置是一个采用多路温度巡检仪测定2 个不同发酵装置里郫县豆瓣实时温度的设备。发酵罐与晒缸里都平均分布有上、中、下3层,每层平均5 个测温点,测温装置对每个测温点每隔2 min 记录1 次温度,在整个发酵期间全程不间断记录。发酵开始时,将从工厂取回的成熟甜瓣子与成熟椒醅立即以质量比1∶3 混匀,分别置于自然发酵装置与恒温发酵装置中发酵。自然发酵工艺采取传统“日晒夜露”工艺,不进行人工控温,每天早上8 点搅拌1 次;恒温发酵工艺采取的发酵温度为40℃,发酵过程中每隔3 d 搅拌1 次。2 种工艺发酵过程中,当豆瓣中的水分含量降至50%以下时进行适量补水,使豆瓣中水分含量维持在50%~60%之间。取样时,2 种发酵工艺均为搅拌后立即取样,每隔6 d 取样1 次,整个发酵时间为8月到11月,共90 d。
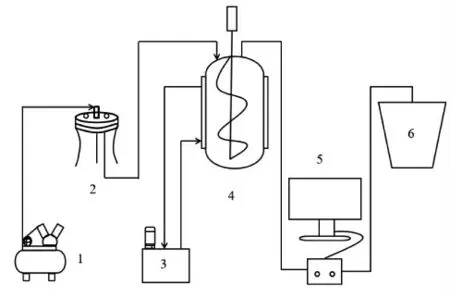
图1 郫县豆瓣自然与恒温后熟发酵装置图Fig.1 Schematic of natural and constant temperature post-fermentation of Pixian broad-bean paste
1.5 方法
1.5.1 水分含量的测定 采用GB 5009.3-2010《食品安全国家标准 食品中水分的测定》[19]中的直接干燥法测定水分含量。
1.5.2 总酸与氨基酸态氮含量的测定 采用GB 5009.235-2016《食品安全国家标准 食品中氨基酸态氮的测定》[20]中的酸度计法测定总酸和氨基酸态氮含量。
1.5.3 还原糖含量的测定 采用3,5-二硝基水杨酸(DNS)法[21]测定还原糖的含量。
1.5.4 色价的测定 采用GB 1886.34-2015《食品安全国家标准 食品添加剂 辣椒红》[22]中的分光光度法测定色价。
1.5.5 氨基酸含量的测定 采用GB/T 30987-2014《植物中游离氨基酸的测定》[23]中的全自动氨基酸分析仪法测定氨基酸的含量。
1.5.6 数据处理采用Excel 和Origin 8.5 对试验数据进行处理与分析以及绘制相关图表。
2 结果与分析
2.1 豆瓣发酵过程中水分的动力学变化
由自然和恒温发酵过程中的水分变化情况可知,其水分随时间变化分别符合指数与线性关系,选用的动力学方程如式(1)和式(2)所示。

式(1)和(2)中,y 分别为自然与恒温发酵的水分含量(g/100 g);ym,y0分别为自然与恒温发酵的水分最终含量和初始含量(g/100 g);b,c 为速率常数(d-1);a 为自然发酵水分减少的最大量(g/100 g)。拟合参数见表1,拟合曲线如图2所示。

表1 水分动力学参数Table 1 Kinetic parameters of moisture
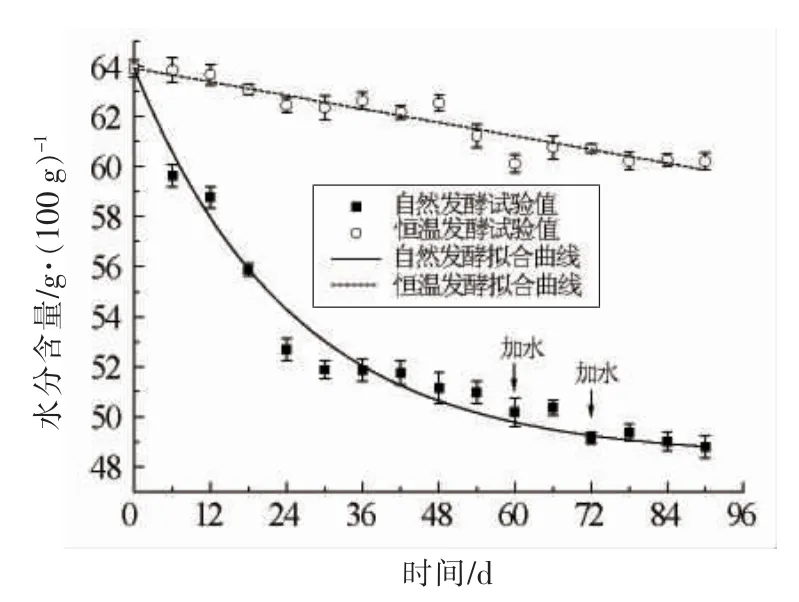
图2 不同发酵模式下水分含量随时间的变化及拟合曲线Fig.2 Variations of moisture content with time and fitting curves in different fermentation modes
由模拟结果可以看出,自然发酵模式下的水分变化基本符合指数模型,水分散失速率不断降低,其平均值为0.04 g/100 g d。从整个发酵过程可以看出,水分减少速率呈现出先快后慢直至接近于零的变化趋势,这是由于自然发酵模式采用敞口发酵,空气流动量大,发酵初期环境温度较高,且豆瓣中含水量也较高,其表面的水汽分压与空气中的水汽分压差距较大,从而使豆瓣中的水分能够大量、快速地散失到空气中,因此发酵前期物料的水分散失速率较快;而随着发酵的进行,环境温度不断降低,并且随着豆瓣中含水量的减少,其与空气中含水量的差值逐渐减小,即豆瓣的含水量逐渐接近平衡含水量,因此水分的传质推动力不断降低,进而导致水分散失率不断下降直至趋近于零。恒温发酵模式下水分变化基本符合一次函数,水分减少速率为定值(0.05 g/100 g d),造成这种现象的原因是恒温发酵采取的是半封闭发酵,空气流通量小,限制了罐内水分向外界传递,进而降低了水分的传递速率,同时恒温发酵过程中温度不变,且豆瓣内自由水含量较高,使得水分传质推动力基本不变,导致水分传递速率几乎保持不变。从以上结果分析可以看出,虽然2 种发酵工艺的水分变化速率值相差不大,但是由于自然发酵受环境因素影响较大,导致其工艺更为繁杂,水分变化情况更多样化,因此自然发酵拟合得到方程也相对复杂,说明恒温发酵更容易在实际生产中实现对于水分的控制。
此外,根据表1的拟合结果可得到,按照目前的发酵方式,自然发酵模式下,豆瓣水分的最低含量(ym)预测值为48.39 g/100 g,发酵过程中减少的水分含量预测值(a)为15.52 g/100 g。然而,发酵90 d 时得到的水分含量试验值为48.80 g/100 g,减少的水分含量试验值为15.11 g/100 g,仅比预测的结果高0.41 g/100 g,说明即使再延长发酵时间,水分含量也不会有明显变化。由于在实际生产中发酵时间越长,人力物力成本越高,因此90 d 的发酵时间基本符合产品要求,延长发酵时间可能反而会导致资源的浪费,造成生产成本的增加。
2.2 豆瓣发酵过程中总酸的动力学变化
2 种发酵方式的总酸含量变化情况均符合指数关系,选用的动力学方程如式(3)所示。

式(3)中,y——发酵过程中总酸含量(g/100 g);ym——发酵过程中总酸的最大生成量(g/100g);a——发酵过程中总酸增加的最大量(g/100 g);b——速率常数(d-1)。拟合参数见表2,拟合曲线如图3所示。
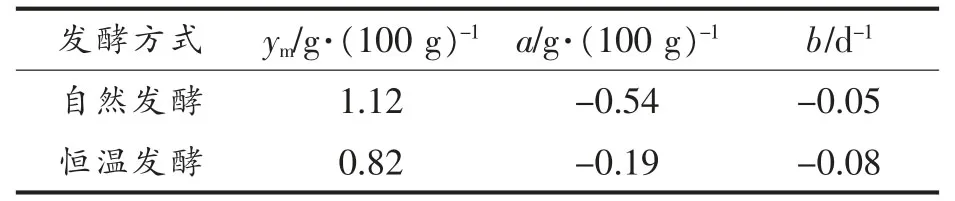
表2 总酸动力学参数Table 2 Kinetic parameters of total acid
由式(3)和表2可知,总酸含量变化的平均速率b 在自然与恒温发酵下分别为-0.05 d-1和-0.08 d-1,表明自然发酵模式下总酸生成速率更快,这可能是由于自然发酵过程中发酵温度变化极为复杂,且晒缸温度分布不均匀,导致可适应发酵条件的微生物种类和数量更多,各种生化反应更剧烈,造成了总酸生成速率更快。发酵过程中总酸增加的最大量a 在自然发酵模式下为0.54 g/100 g,约为恒温发酵(0.19 g/100 g)的3 倍,说明自然发酵模式下郫县豆瓣总酸的生成量更多;并且从拟合方程还可以预测得到自然发酵模式下总酸的最大生成量ym为1.12 g/100 g,大于恒温发酵(0.82 g/100 g)。实际生产中国标要求总酸含量不能过高,因为一旦偏高就有可能是发酵过程中微生物群落失衡,腐败酸含量增加,导致产品质量不合格[24],所以采用恒温发酵更有利于总酸的控制。
由图3可知,自然与恒温发酵模式下总酸含量变化均呈先增加后不变的趋势,且分别在60 d和30 d 由加速期进入稳定期,最终达到预测最大值,由于恒温发酵模式下的总酸增长速率和加速期的时间均小于自然发酵,使得总酸的最大生成量小于自然发酵,同时也说明恒温发酵模式下总酸的发酵周期更短,由于发酵周期越短,实际生产效率越高,因此在对总酸进行调控发酵时,可以适当缩短发酵时间,进而提高生产效率。

图3 不同发酵模式下总酸含量随时间的变化及拟合曲线Fig.3 Variations of total acid content with time and fitting curves in different fermentation modes
2.3 豆瓣发酵过程中氨基酸态氮的动力学变化
根据2 种模式下氨基酸态氮含量的动态变化,选用Logistic 曲线[25]描述其生成过程,方程见式(4)。

式(4)中,y——氨基酸态氮含量(g/100 g),ym为氨基酸态氮最大生成量(g/100 g),y0——氨基酸态氮初始量(g/100 g),μ——比生成速率(d-1),拟合参数见表3,拟合曲线如图4所示。

表3 氨基酸态氮动力学参数Table 3 Kinetic parameters of amino nitrogen
由表3可知,自然发酵下,氨基酸态氮最大生成量ym为0.28 g/100 g,比恒温发酵的0.22 g/100 g 高0.06 g/100 g,均满足一级豆瓣高于0.2 g/100 g 的要求。由图4可知,2 种模式下氨基酸态氮含量的拟合曲线有显著差异,造成这种差异的直接原因主要有2 个:一是自然发酵下的增长速率明显高于恒温发酵,由拟合参数也可知自然发酵的比生成速率μ是恒温发酵的近3 倍,使得自然发酵的氨基酸态氮最大生成量明显高于恒温发酵;二是自然发酵下的延滞期明显少于恒温发酵,从方程可知,氨基酸态氮随时间的生成曲线是一个典型的S 型曲线,一般可将其分为延滞期、加速期和稳定期,恒温发酵模式下的延滞期长达24 d,而自然发酵模式下几乎没有延滞期,发酵开始直接进入加速期,从而更快的进入稳定期同时得到更高的累积量,这也提示若要在发酵过程中获得更高的氨基酸态氮生成量,则需进一步明确影响氨基酸态氮生成的因素,缩短发酵过程的延滞期并延长其加速期,从而提高氨基酸态氮的比生成速率。
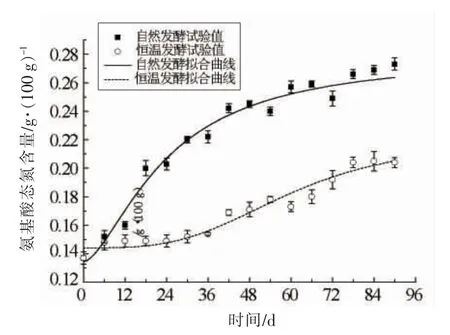
图4 不同发酵模式下氨基酸态氮含量随时间的变化及拟合曲线Fig.4 Variations of amino nitrogen content with time and fitting curves in different fermentation modes
2.4 豆瓣发酵过程中还原糖的动力学变化
根据还原糖含量的动态变化,选用GaussAmp模型[26]描述其生成过程,动力学方程为:

式(5)中,y——还原糖含量(g/100 g);y0——还原糖最小终止生成量(g/100 g),A 为时间变化对还原糖含量影响的总量因子,xC——还原糖含量最大时对应的时间(d),ω——发酵时间范围(d)。拟合参数见表4,拟合曲线如图5所示。

表4 还原糖动力学参数Table 4 Kinetic parameters of reducing sugar
由图5和表4可知,自然发酵和恒温发酵还原糖含量变化速率为零的点即拐点,分别出现在第51 天和第27 天,说明自然发酵还原糖含量的增长速率时间约是恒温发酵的2 倍,这提示如果恒温发酵要得到更高的还原糖含量,则需要改变工艺条件来促进相关微生物和酶作用的发挥,从而增加还原糖累积的时间。自然发酵下总量因子A 是恒温发酵的10 倍,说明自然发酵更容易受环境变化的影响,也进一步表明环境的差异对豆瓣发酵的影响较大,从而出现了2 种发酵模式下还原糖累积量差异显著的现象,这也提示在工艺设计过程中,研究如何改变发酵条件以增大A 值促进还原糖的生成具有重要意义。由拟合结果可知,还原糖最小终止生成量y0在自然发酵下为1.02 g/100 g,恒温发酵下为2.13 g/100 g,这提示自然发酵如果要得到更高的还原糖含量,可以通过缩短还原糖含量下降的时间实现。
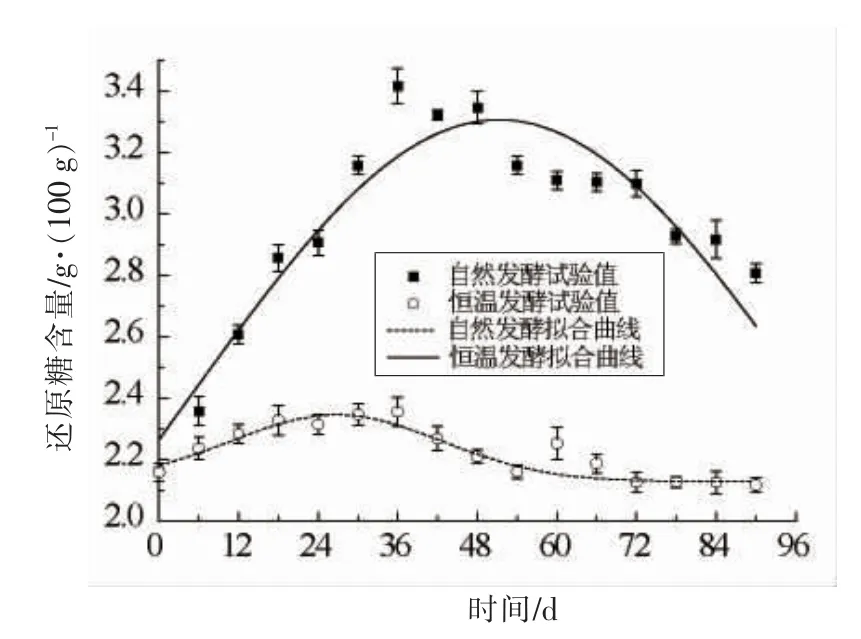
图5 不同发酵模式下还原糖含量随时间变化及拟合曲线Fig.5 Variations of reducing sugar content with time and fitting curves in different fermentation modes
此外,由图5及表4可知,还原糖生成量的变化曲线均呈现出先增加后减少的趋势,然而自然发酵下增加和减少的速率均明显高于恒温发酵,这是因为在后发酵阶段高盐环境让霉菌停止生长甚至死亡,但它所分泌的淀粉酶和糖化酶等仍能作用于淀粉产生还原糖。贾瑞博等[27]研究发现米曲霉所产的淀粉酶最适反应温度为50℃,由于自然发酵模式下豆瓣能达到的最高温度(50℃)高于恒温发酵(40℃),因此其酶活性高于恒温发酵,导致其还原糖的生成速率高于恒温发酵。孙常雁[28]认为淀粉酶有一定的耐盐性,活性虽然受到部分抑制,却仍能发挥作用,如果长时间处于高盐环境中酶活性会明显减弱。赵晓娟[29]认为豆酱发酵后期酸度增加,淀粉酶和糖化酶活力降低。本研究中,郫县豆瓣酱中的食盐含量高达20%,且豆瓣酱中的酸度也在不断上升,因此随着发酵的进行,酱醅中的淀粉酶和糖化酶等酶的活性逐渐减弱,还原糖增长速率逐渐减慢,直至为零。之后还原糖含量的逐渐减少,则是由于自然发酵模式下充足的氧环境加速了美拉德反应[30],还原糖作为反应物被快速消耗,因此还原糖含量的下降速率也更快,最终出现了自然发酵下还原糖含量的大幅增长和大幅降低的现象,即通过模型预测分析得到,自然发酵下可得到的还原糖最大生成量比恒温发酵高0.96 g/100 g,最小终止生成量比恒温发酵低1.11 g/100 g。恒温发酵下的还原糖含量变化相对稳定,且发酵周期更短,但也有可能限制了美拉德反应的进行,进而使得恒温发酵产品的风味不足,因此进一步改进恒温发酵的发酵条件,促进还原糖的生成,也将是完善恒温发酵工艺的一个重要研究点。
2.5 豆瓣发酵过程中色价的动力学变化
根据试验结果,自然发酵和恒温发酵模式下的色价变化趋势基本符合线性关系,选用的动力学方程如式(6)所示。

式(6)中,y——色价值;y0——发酵过程中色价的初始值,a——速率常数(d-1)。拟合参数见表5,拟合曲线见图6。
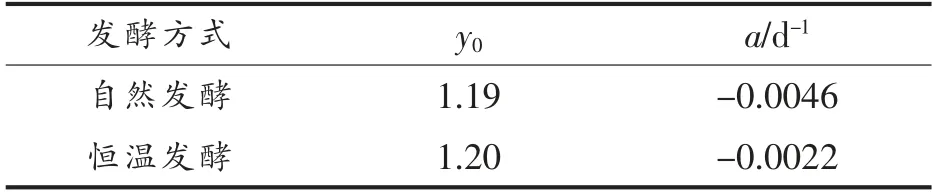
表5 色价动力学参数Table 5 Kinetic parameters of chromaticity
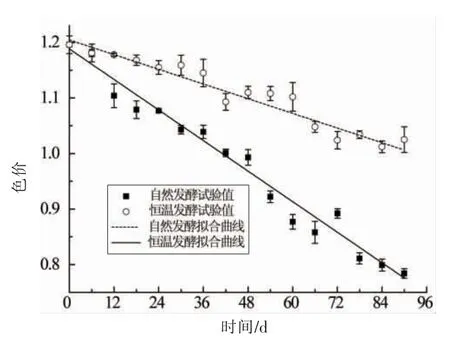
图6 不同发酵模式下中色价随时间的变化及拟合曲线Fig.6 Variations of chromaticity with time and fitting curves in different fermentation modes
由拟合方程(6)及表5可知,自然发酵下色价下降速率a 为0.0046 d-1 约为恒温发酵(0.0022 d-1)的2 倍,自然发酵下色价下降速率更快,这是因为辣椒红色素是影响豆瓣色价的主要因素,而影响辣椒红色素稳定性的因素有很多,如温度、水分、光照、pH 值、空气中的氧和金属离子等[31],Jar én-Gal án 等[32]研究表明,辣椒红色素含量随温度升高而降低,这与本试验结果一致。自然发酵过程温度较高,最高时能超出恒温发酵10℃,使得辣椒红色素含量下降得更快,Shin 等[33]研究认为辣椒色素不耐光,强光下很快分解,因此光照强度更大的自然发酵模式下辣椒红色素减少更多,同时自然发酵的开放式发酵让豆瓣表面能更充分的接触氧气,使得自氧化反应更剧烈,因此自然发酵的色价损失更多,使得最终色价值更小,说明半封闭式恒温发酵更有利于豆瓣产品色价的保持。色价越高,颜色越油润有光泽,产品质量越好,提示采用封闭式发酵可能会提高豆瓣品质。
2.6 豆瓣发酵过程中氨基酸含量随时间的变化
根据Tseng 等[34]对氨基酸滋味的描述,将其分为鲜味、甜味、苦味和无味,自然与恒温发酵分别选用指数和线性模型描述其生成过程,其方程分别为:

式(7)和式(8)中,y——分别为自然与恒温的氨基酸含量(mg/g);ym,y0——分别为自然与恒温发酵的氨基酸最终生成量和初始生成量(mg/g);b,d——速率常数(d-1);a——自然发酵随发酵时间增加的氨基酸变化量(mg/g)。拟合参数见表6,拟合曲线见图7。
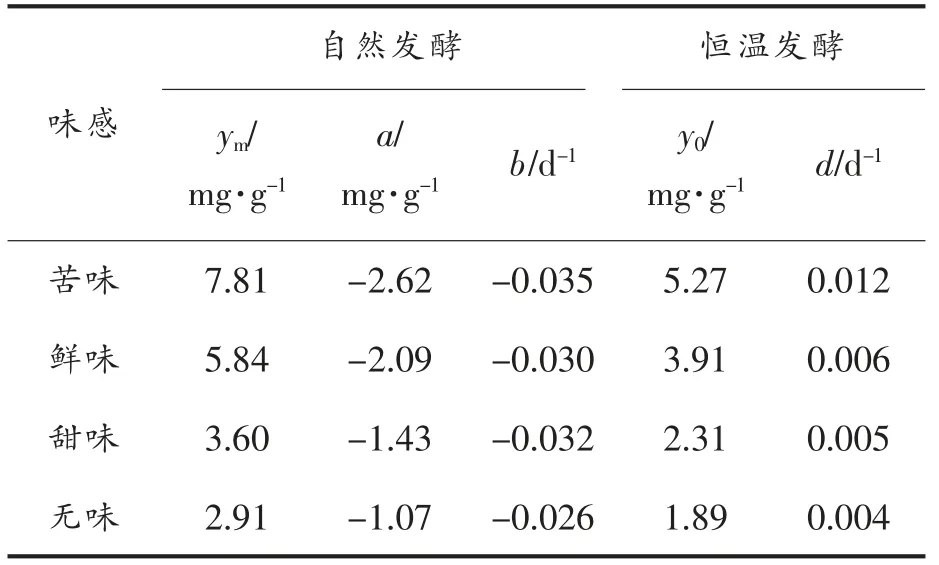
表6 氨基酸动力学参数Table 6 Kinetic parameters of free amino acids
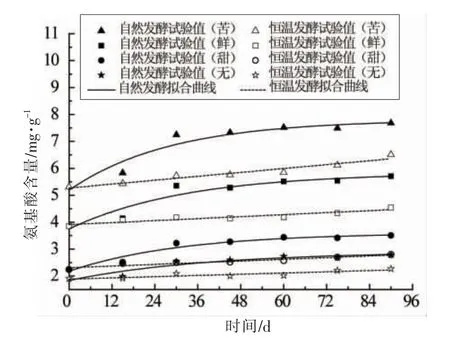
图7 不同发酵模式下氨基酸含量随时间变化及拟合曲线Fig.7 Variations of free amino acids content with time and fitting curves in different fermentation modes
由式(7)可知,自然发酵的氨基酸含量呈指数型增长,发酵初期快速增加,发酵后期基本不变,这是因为发酵初期,自然发酵温度较高,酶活性较好,加速了蛋白质分解生成氨基酸的过程,随着温度的降低,酶活受到抑制,氨基酸累积量减慢直至不变[35];由式(8)可知,恒温发酵的氨基酸含量保持较小的速率呈线性增长,这是因为恒温发酵温度保持不变,发酵温度并不完全适宜蛋白质的分解,所以氨基酸累积量保持一个较小的速度缓慢上升。
由表6可知,自然发酵下的各种味感氨基酸增长速率b 值均大于恒温发酵d 值,表明自然发酵更有利于氨基酸的生成,提示恒温发酵可以参考自然发酵的工艺条件进行改进,增大生成速率,从而更快增加氨基酸含量。自然发酵下,4 种呈味氨基酸中苦味氨基酸的增长速率最高,为0.035 d-1,是其它3 种呈味氨基酸增长速率的1.09~1.35倍;恒温发酵下,4 种呈味氨基酸中苦味氨基酸的增长速率也最高,为0.012 d-1,是其它3 种呈味氨基酸增长速率的2~3 倍,由此可以看出,自然发酵下的4 种呈味氨基酸的增长速率相对均衡。由于豆瓣的特殊滋味是由各种氨基酸的相互作用产生的[36],各氨基酸所占比例一旦发生改变,郫县豆瓣特有的风味也可能随之消失,因此如何改变恒温工艺的发酵条件,缩小各种味型氨基酸之间增长速率的差距具有重要的研究意义。
3 结论
对郫县豆瓣半封闭恒温与传统自然后熟发酵工艺试验数据采用了指数、线性、Logistic 以及GaussAmp 模型进行了拟合与分析,结果表明所采用的动力学模型均能较好的描述各个指标的变化规律。
拟合结果表明,自然发酵模式下,氨基酸态氮生成曲线延滞期短于恒温发酵,还原糖含量达到最大值所用时间长于恒温发酵,4 种呈味氨基酸增长速率均衡性高于恒温发酵,由于这些指标对产品质量的提升有较大帮助,因此恒温发酵的工艺条件可以参考自然发酵进行改进,通过调控各个发酵阶段的持续时间使这些指标朝有利方向发展;半封闭式恒温发酵更容易实现在实际生产中对水分和总酸的控制以及色价的保持,在后续罐式发酵中采用封闭式发酵可能会提高豆瓣品质。
猜你喜欢
桂林理工大学学报(2021年3期)2021-12-06
福建农业科技(2019年8期)2019-09-10
中国民族民间医药·上半月(2019年4期)2019-06-24
中国民族民间医药·上半月(2019年11期)2019-01-16
天然产物研究与开发(2018年9期)2018-10-08
当代县域经济(2017年9期)2017-09-03
西北民族大学学报(自然科学版)(2016年3期)2016-12-20
中国品牌(2016年1期)2016-03-19
摄影世界(2015年9期)2015-09-15
