
瑞士乳杆菌抗高温胁迫补偿应机制
2021-06-02 09:37徐礼云陈延儒董弦弦吴晓江邓梦菲巫小丹刘成梅付桂明
南昌大学学报(理科版) 2021年1期
徐礼云,陈延儒,董弦弦,吴晓江,邓梦菲,巫小丹,万 茵,刘成梅,b,付桂明,b*
(南昌大学a.食品科学与技术国家重点实验室;食品学院;b.国际食品创新研究院,江西 南昌 330047)
瑞士乳杆菌(Lactobaccillushelveticus)是一种常见的具有益生功能的乳酸菌,具有高蛋白水解活性,常被用于强化酸奶,能将乳制品中的蛋白质转化为生物活性肽[1-2]。添加瑞士乳杆菌的发酵乳制品具有减缓疲劳和肌肉损伤的作用,并且食用加入了瑞士乳杆菌的发酵牛奶可以提高老年人的睡眠效率[3-4]。此外,瑞士乳杆菌产生的胞外多糖,具有多种益生特性,如抗氧化和抗癌作用[5]。因此,瑞士乳杆菌可以被认为是一种多功能益生菌,在功能性食品和乳制品工业中有着广阔的应用前景。
工业生产活菌发酵剂时,喷雾干燥由于其价格低廉而得到广泛应用。然而乳酸菌耐热性能弱,在喷雾干燥过程中易遭受高温和脱水等恶劣环境影响,引起细胞膜结构损伤、蛋白质变性和酶活性的丧失等,导致获得的乳酸菌菌剂活力较低[6]。目前提高乳酸菌发酵剂的活力的研究主要集中于通过添加高效抗逆保护剂、特殊营养成分或优化干燥工艺参数等方法[7-8]。如El-Sayed等[6]研究发现,加入黄油可以提高喷雾干燥制得的酸奶粉中瑞士乳杆菌的存活率。
在受外界胁迫时,微生物可通过启动代谢补偿机制,产生一系列的生理和代谢变化,如调节膜的流动性和通透性、增加渗透调节物质(海藻糖,可溶性蛋白)产生和提高抗氧化酶活性等,从而使整个细胞进入对外界胁迫的防御状态[9-10]。但对高温胁迫下瑞士乳杆菌代谢补偿机制的研究较少。因此,本文以实验室保藏的瑞士乳杆菌NCUF 210.1为研究对象,对其高温胁迫后的细胞膜的流动性和通透性,渗透调节物质和抗氧化酶活性进行分析,并通过转录组测序,从分子层面探究高温胁迫下瑞士乳杆菌的补偿效应,以期为瑞士乳杆菌在喷雾干燥等食品热加工条件的选择和耐高温工程菌株的构建提供理论参考。
1 材料与方法
1.1 材料与试剂
1.1.1 供试菌株
瑞士乳杆菌(LactobaccillushelveticusNCUF 210.1),为南昌大学实验室保藏菌株。
1.1.2 培养基
MRS培养基:葡萄糖20 g,蛋白胨10 g,牛肉浸粉8 g,酵母浸粉4 g,柠檬酸氢二铵2 g,CH3COONa 5 g,K2HPO42 g,MnSO4·4H2O 0.04 g,MnSO4·7H2O 0.2 g,吐温-80 1 mL,水1 L。调节pH值为6.0,121 ℃高压灭菌15 min。
1.1.3 试剂
蛋白胨、牛肉浸粉、酵母浸粉:北京奥博星生物技术责任有限公司;其它试剂均为国产分析纯。
1.2 仪器与设备
DDS-307型电导率仪,上海仪电科学仪器股份有限公司;SP-756P可见分光亮度计,上海光谱仪器有限公司;F-700荧光分光光度计,日本日立高新技术公司;SPX-80S-Ⅱ恒温培养箱,上海新苗实验仪器有限公司。
1.3 方法
1.3.1 瑞士乳杆菌的活化
将实验室保藏的瑞士乳杆菌在MRS固体培养基平板上划线分离后,挑取单菌落在MRS液体培养基中进行活化,37 ℃恒温培养24~48 h。
1.3.2 不同培养温度条件下瑞士乳杆菌生长趋势的研究
将活化后的瑞士乳杆菌按1%的接种量接种于MRS液体培养基,分别于37 ℃,39 ℃,41 ℃,42 ℃,45 ℃,47 ℃和49 ℃下培养。每隔2 h取样一次,采用比浊法,测定波长600 nm的吸光值。绘制其在不同培养温度下的生长曲线[11]。
1.3.3 细胞膜流动性的测定
根据杨胜平等[12]的方法并略有修改。将瑞士乳杆菌种子液按1%的接种量接种于50 mL的MRS液体培养基中,置于42 ℃和47 ℃培养,分别于8,10和12 h时取样,4 ℃下7 000 r·min-1离心10 min,收集菌体进行细胞膜流动性的测定。
1.3.4 细胞膜透性的测定
参照史明科[10]的方法并略有修改。将瑞士乳杆菌种子液按1%的接种量接种于50 mL的MRS液体培养基中,置于42 ℃和47 ℃培养,分别于8,10和12 h时取样,4 ℃下7 000 r·min-1离心10 min,收集菌体沉淀进行电导率的测定。
1.3.5 可溶性蛋白含量测定
可溶性蛋白含量测定采用考马斯亮蓝染色法[10]。将瑞士乳杆菌种子液按1%的接种量接种于50 mL的MRS液体培养基中于42和47 ℃培养,分别于8,10和12 h时取样,4 ℃下7 000 r·min-1离心10 min,收集菌体。经磷酸缓冲盐溶液清洗2次,重悬于5 mL磷酸缓冲盐溶液中,4 ℃下10 000 r·min-1离心20 min,取1.0 mL上清液加入5.0 mL考马斯亮蓝G-250溶液后静置5 min后,测定波长595 nm的吸光值。对照标准曲线计算可溶性蛋白含量,单位为mg/g细胞干重。
1.3.6 海藻糖含量测定
海藻糖含量测定参照史明科[10]的方法并略有修改。将瑞士乳杆菌种子液按1%的接种量接种于50 mL的MRS液体培养基中于42和47 ℃培养,分别于8,10和12 h时取30 mL菌样,4 ℃下7 000 r·min-1离心10 min,收集菌体。经磷酸缓冲盐溶液清洗2次,加入5 mL 10%三氯乙酸,冰水浴1 h,4 ℃下7 000 r·min-1离心10 min。取稀释至合适倍数的上清液(1 mL)进行海藻糖的含量的测定。将菌体沉淀置于105 ℃干燥以评估菌体生物量。对照标准曲线计算海藻糖含量,单位为mg·g-1细胞干重。
1.3.7 超氧化物歧化酶(SOD)活性测定
SOD活性测定参照史明科[10]的方法并略有修改。将瑞士乳杆菌种子液按1%的接种量接种于50 mL的MRS液体培养基中于42和47 ℃培养,分别于8,10和12 h时取样,4 ℃下7 000 r·min-1离心10 min,收集菌体。经磷酸缓冲盐溶液清洗2次,重悬于5 mL磷酸缓冲盐溶液中,4 ℃下10 000 r·min-1离心20 min,弃沉淀,取上清液进行SOD活性测定。
1.3.8 转录组技术分析高温胁迫下瑞士乳杆菌差异表达基因
将高温胁迫(47 ℃,10 h)与最适生长温度(42 ℃,10 h)下的瑞士乳杆菌菌体沉淀保存于液氮中用于转录组测序,设置3个生物学重复用于转录组测序。由上海美吉生物医药科技有限公司完成RNA的提取、质控、建库及测序工作。通过生物信息学手段对差异表达基因进行分析,明确其相应生物学功能。
1.4 数据分析
采用SPSS Statistics 19软件进行单因素方差分析。采用Origin 2018和Excel 2010进行数据统计和图表制作。每组实验重复3次。
2 结果与分析
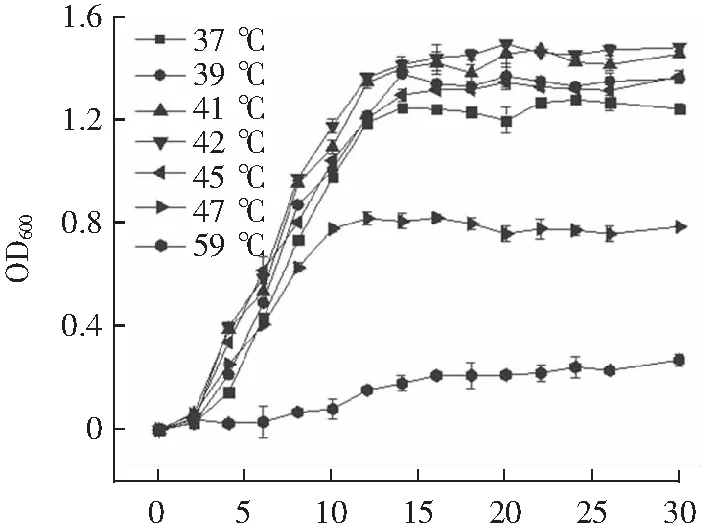
2.1 在不同培养温度条件下瑞士乳杆菌的生长趋势
由图1可知,不同生长温度下瑞士乳杆菌生长趋势相同。其中42 ℃是瑞士乳杆菌的最适生长温度,与岳莹雪等[5]结果一致。培养12 h后瑞士乳杆菌开始进入稳定期,OD值最大为1.387±0.03。当培养温度提升至47 ℃时,瑞士乳杆菌在培养10 h开始进入稳定期,OD值为0.794±0.02,与置于42 ℃培养相比,其生长受到明显的抑制。处于对数生长后期的瑞士乳杆菌对外界胁迫的耐性高于对数前期和中期[13],所以本实验选择42℃作为对照,探究高温胁迫(47 ℃)对培养8,10和12 h的瑞士乳杆菌细胞补偿效应的影响。
2.2 高温胁迫对瑞士乳杆菌的细胞膜流动性的影响
适宜的细胞膜流动性对维持细胞膜的正常生理功能有重要意义。当细胞受到外界不良环境的影响时,流动性会降低,以提高细胞的抗逆性[14]。采用ANS荧光探针法对细胞膜流动性进行测定[12]。ANS的荧光强度值与细胞膜的流动性呈反比,荧光强度越高,细胞膜的流动性越低。如图2所示,相比最适生长温度(42 ℃),瑞士乳杆菌在高温胁迫(47 ℃)下荧光强度值升高,其细胞膜流动性显著降低,这可能是由于高温胁迫引起膜脂肪酸组成的变化,导致膜流动性下降[15]。张爱静等[14]对大肠杆菌进行热胁迫,发现随着胁迫温度的提高,细胞膜流动性降低,与本研究结果趋势一致,且菌株细胞膜流动性的降低可能有利于保证其在高温环境下正常的流动性,使细胞更好地适应外界胁迫环境。在高温胁迫(47 ℃)下,瑞士乳杆菌在稳定期(12 h)时的细胞膜流动性高于对数期后期(10 h),这可能是由于瑞士乳杆菌经自身代谢补偿逐渐适应了高温环境。
t/h
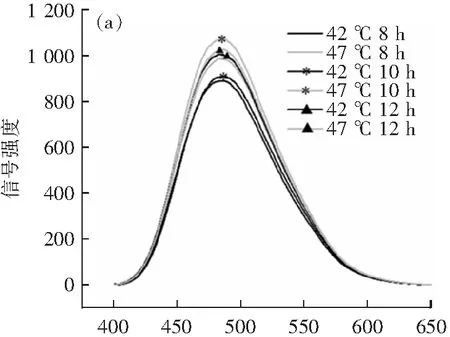
λ/nm
2.3 高温胁迫对瑞士乳杆菌的相对电导率的影响
细胞膜是保护和维持瑞士乳杆菌细胞稳定的重要组成部分,是细胞抵抗外界胁迫的重要屏障,也是反映细胞完整性的重要指标。微生物在遭受外界胁迫时菌体细胞膜通透性增加,导致一些钾盐、磷酸盐等小分子释放,使发酵液电导率升高[16]。由图3可知,与最适生长温度(42 ℃)相比,瑞士乳杆菌在高温胁迫(47 ℃)下相对电导率分别提高了11.43%(8 h),10.81%(10 h)和12.97%(12 h)。这可能是因为高温胁迫导致细胞膜蛋白质变性,渗透调节紊乱,同时细胞膜通透性增大,造成内部电解质的外渗。与最适生长温度(42 ℃)相比,瑞士乳杆菌在高温胁迫条件(47 ℃)下,随着培养时间的延长,发酵液的电导率显著升高(P<0.05),之后趋于平稳。这可能是由于瑞士乳杆菌对高温胁迫产生完全应答需要一定时间,在此期间细胞不能适应高温胁迫环境,可能导致细胞膜的屏障作用减弱,通透性增加,引发包括电解质在内的细胞内容物外泄,从而造成发酵液相对电导率逐渐增大。随着高温胁迫时间的延长,细胞可能逐渐完成细胞应答过程,适应了高温胁迫环境,胞内电解质外泄速度减缓,因此发酵液相对电导率趋于平稳。
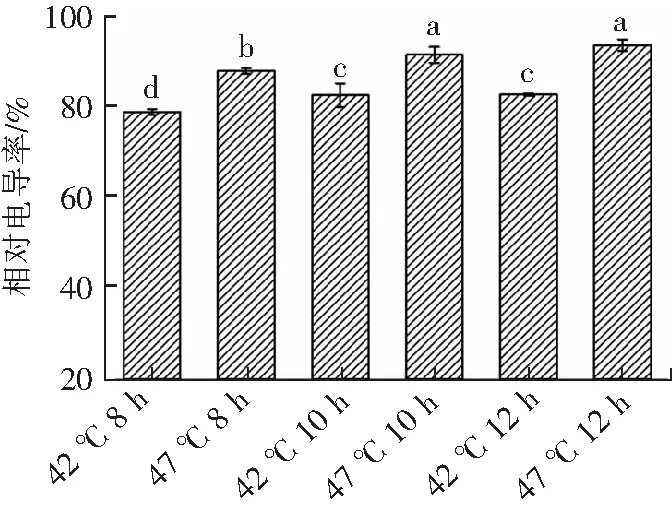
不同上标小写字母表示具有显著差异(P<0.05)。
2.4 高温胁迫对瑞士乳杆菌细胞可溶性蛋白含量的影响
研究发现微生物在遭受外界胁迫时,在其适应过程中,细胞会产生了一系列的应答反应,通过产生大量渗透调节物质(如可溶性蛋白,海藻糖等),提高生物对外界胁迫的适应性[10]。由图4可知,与最适生长温度(42 ℃)相比,瑞士乳杆菌在高温胁迫(47 ℃)下可溶性蛋白含量分别提高了77.46%(8 h),140.13%(10 h)和31.62%(12 h)。这可能是外界胁迫导致细胞渗透调节紊乱,合成酶活性下降,水解酶活性增加,诱导蛋白质转变成可溶性化合物,细胞内可溶性化合物增加,因而增强细胞的渗透势,提高其渗透调节的能力,保护细胞膜,从而增强其耐热性[10]。在高温胁迫(47 ℃)下,瑞士乳杆菌在稳定期(12 h)时的可溶性蛋白含量显著低于对数期后期(10 h)(P<0.05),这可能是由于瑞士乳杆菌在稳定期(12 h)时经自身代谢补偿逐渐适应了高温环境,水解酶活性降低,可溶性蛋白质的生成降低。
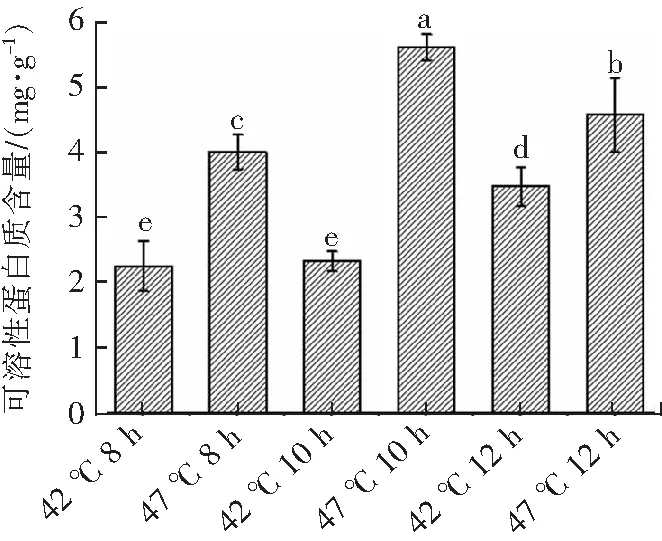
不同上标小写字母表示具有显著差异(P<0.05)。
2.5 高温胁迫对瑞士乳杆菌细胞海藻糖含量的影响
海藻糖是一种重要的渗透调节物质。外界胁迫会造成生物体内海藻糖的积累,从而保护细胞质膜,核酸等生物大分子空间结构和功能活性,维持细胞渗透压,帮助细胞抵抗外界不良环境[10,17]。如图5所示,瑞士乳杆菌在最适生长温度(42 ℃)培养下海藻糖含量最低;在高温胁迫(47 ℃)下海藻糖含量分享提高了30.12%(8 h),17.23%(10 h)和21.95%(12 h);这可能由于海藻糖是一种生物应激代谢产物,生长在不良环境中微生物细胞中海藻糖含量较高[18]。海藻糖对生物体的保护作用是由于它提高了细胞内渗透调节物质含量,使细胞渗透压降低,维持细胞膜的完整性,从而提高细胞的抗外界胁迫能力[10]。叶燕锐等[19]发现在42 ℃热胁迫下,酿酒酵母细胞内的海藻糖含量较胁迫前提高了4倍,表明海藻糖的积累能够增强细胞对外界胁迫的抗性。近年来的报道中还发现,在生物体处于胁迫条件时,海藻糖可通过诱导自噬的发生,从而保护细胞,使其免于凋亡,最终使生物体在极端环境中得以存活[20]。在高温胁迫(47 ℃)下,瑞士乳杆菌在稳定期(12 h)和对数期后期(10 h)的海藻糖含量变化不显著(P>0.05),这可能是由于瑞士乳杆菌经自身代谢补偿逐渐适应了高温环境,海藻糖合成减缓。
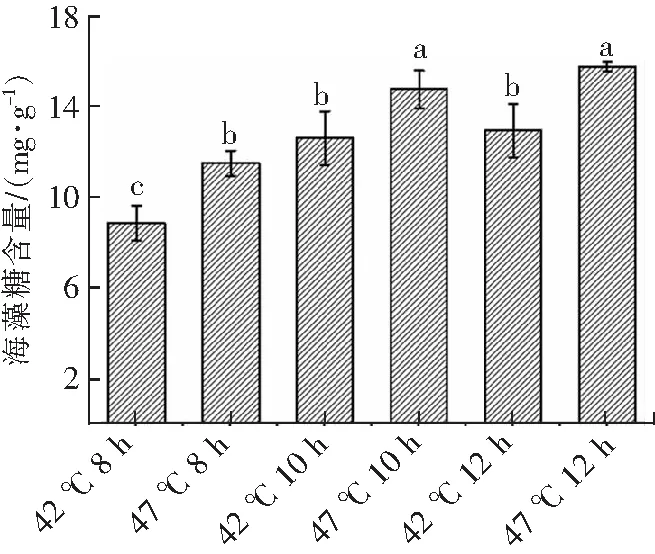
不同上标小写字母表示具有显著差异(P<0.05)。
2.6 高温胁迫对瑞士乳杆菌中SOD活性的影响
外界胁迫会增加生物体内活性氧自由基的产生,导致活性氧自由基的产生和清除间的平衡被破坏,造成膜脂过氧化,从而对细胞造成损伤[21]。SOD是生物体抗氧化酶系统重要组分,能够清除生物体产生的自由基[10]。在高温胁迫条件下,瑞士乳杆菌SOD活性有不同程度的变化(图6)。由图6可知,与最适生长温度(42 ℃)相比,瑞士乳杆菌在高温胁迫(47 ℃)时SOD活性分别提高了97.24%(8 h),159.28%(10 h)和126.96%(12 h),差异显著(P<0.05)。这可能是由于瑞士乳杆菌在高温胁迫下通过激发补偿效应,提高的SOD活性,增强清除活性氧的能力,减小膜脂过氧化作用,降低高温对细胞造成的伤害,从而达到保护细胞的作用[10]。
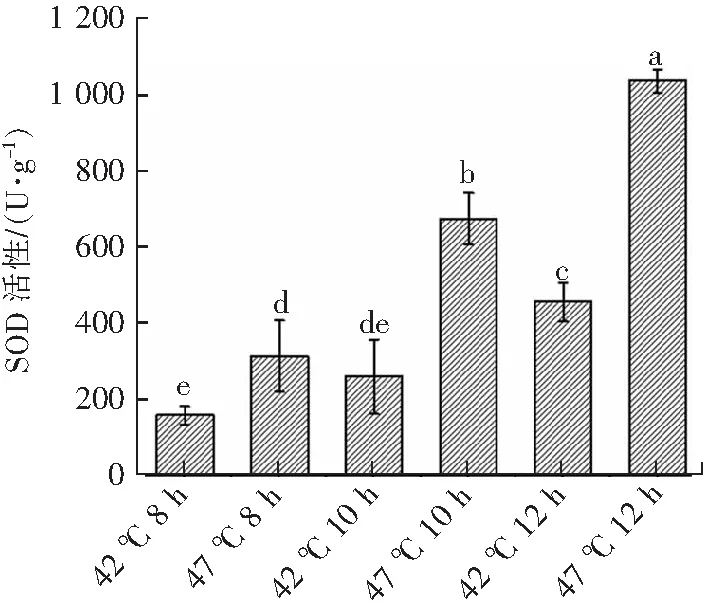
不同上标小写字母表示具有显著差异(P<0.05)。
2.7 高温胁迫下瑞士乳杆菌转录组差异表达基因分析
从高温胁迫对瑞士乳杆菌的生长情况、细胞膜通透性和流动性、可溶性蛋白含量、海藻糖含量和SOD活性多个指标的变化分析,选取高温胁迫(47 ℃,10 h)与最适生长温度(42 ℃,10 h)的瑞士乳杆菌送往上海美吉生物医药科技有限公司进行转录组测序,探究高温胁迫下瑞士乳杆菌的转录水平变化情况。选择差异倍数|log2(fold change)|>1和显著水平q value<0.05的差异基因进行分析。结果如图7所示,与最适生长温度(42 ℃)相比,高温胁迫(47 ℃)下的瑞士乳杆菌中有432个基因的调控模式发生变化。其中,有219个基因转录水平上调,213个基因转录水平下调。由此可见,高温胁迫(47 ℃)下瑞士乳杆菌基因转录水平发生一定改变。
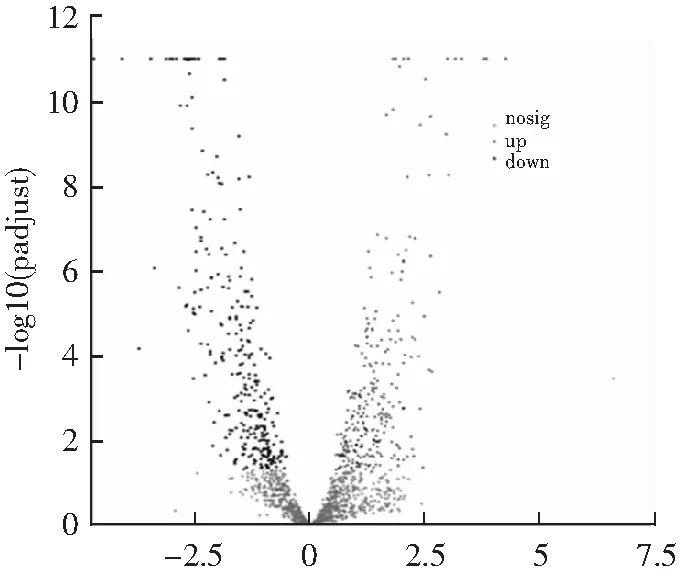
log2(fold change)
2.8 高温胁迫下瑞士乳杆菌差异表达基因GO和KEGG注释分类
将432个差异基因根据不同的功能进行GO(Gene Ontology)功能注释及分类,结果如图8所示。GO功能分为3大类:生物过程(biological process)、细胞组分(cellular component)及分子功能(molecular function)。在生物过程上的10个亚分类中,以代谢过程(metabolic process)和细胞过程(cellular process)居多,占该类别的比例分别为35.18%和33.10%,其次是生物调控(biological regulation)、应激反应(response to stimulus)和定位(localization),分别占比5.09%,3.47%和2.08%。其中,应激反应中有10个差异基因转录水平上调,5个差异基因转录水平下调。瑞士乳杆菌可能通过增加应激反应过程的差异基因表达,积累更多的热激蛋白,提高其抗热胁迫的能力。在细胞组分类别中,差异表达基因主要与细胞构造(cellular anatomical entity)和含蛋白质复合物(protein-containing complex)有关,这2类功能占该大类的比例分别为35.65%和2.3%。在分子功能的10个亚分类中,差异表达基因主要与催化活性(catalytic activity)和结合(binding)有关,分别占该类别的51.62%和40.27%。其次是结构分子活性(structural molecule activity)和转运活性(transporter activity),分别占比5.56%和5.09%。其中,催化活性中有125个差异基因转录水平上调,98个差异基因转录水平下调;抗氧化活性(antioxidant activity)中有2个差异基因转录水平上调。以上结果表明,高温胁迫影响了瑞士乳杆菌的催化活性和抗氧化活性等分子功能、细胞构造和含蛋白质复合物等细胞组分以及代谢过程和应激反应过程等生物过程。
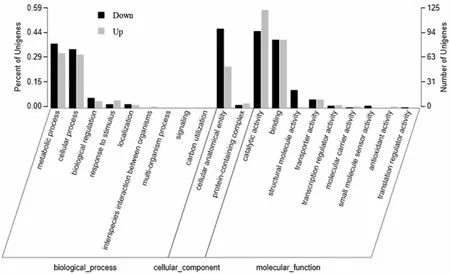
图8 差异表达基因的GO分类图
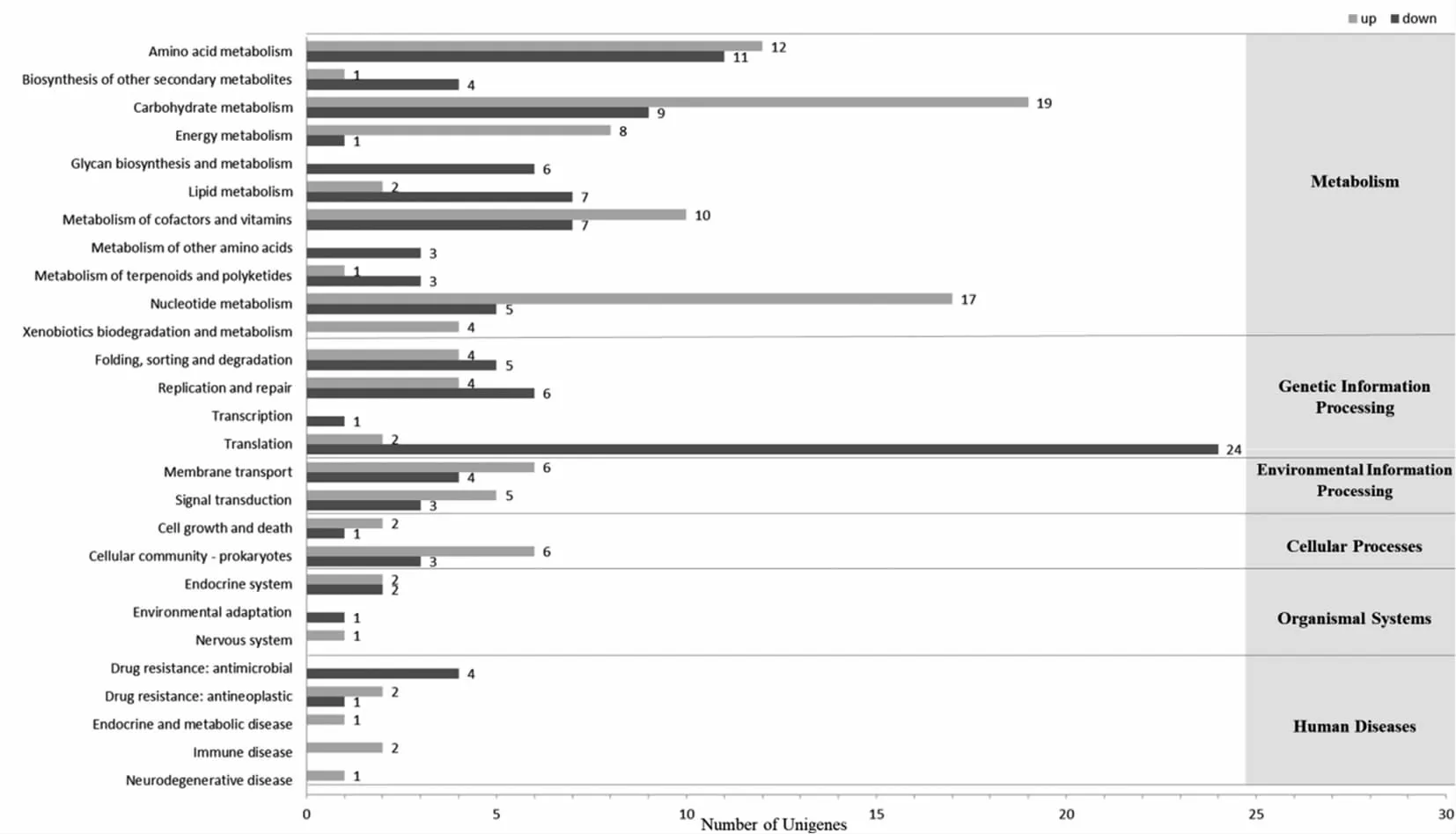
图9 差异表达基因的KEGG功能分类图
为了解差异基因的生物学功能,对差异表达基因进行KEGG(Kyoto Encyclopedia of Genes and Genomes)注释。结果如图9所示,高温胁迫下瑞士乳杆菌在KEGG数据库中共有223个差异表达基因分别在代谢(Metabolism),遗传信息处理(Genetic Information Processing),环境信息处理(Environmental Information Processing),细胞过程(Cellular Processes),生物体系统(Organismal Systems),人类疾病(Human Diseases)6大功能27个通路上得到功能注释。结果显示,130个差异基因在代谢通路上得到注释,其中56个差异基因转录水平上调,74个差异基因转录水平下调。代谢途径层面的差异基因主要集中在糖代谢(Carbohydrate metabolism)、氨基酸代谢(Amino acid metabolism)和核苷酸代谢(Nucleotide metabolism)等代谢。上调差异基因中,糖代谢途径中有19个差异基因转录水平上调;氨基酸代谢途径中有12个差异基因转录水平上调;核苷酸代谢途径中有17个差异基因转录水平上调。表明氨基酸代谢、糖代谢和核苷酸代谢水平提高,可促进相关功能蛋白和酶的合成,对高温胁迫的瑞士乳杆菌细胞损伤具有重要修复作用。Gao等[22]研究表明植物乳杆菌可通过提高核苷酸代谢水平,为ATP合成提供更多的腺嘌呤,从而为抵抗恶劣环境提供能量。遗传信息处理途径中有10个差异基因转录水平上调,36个差异基因转录水平下调。瑞士乳杆菌处于47 ℃高温胁迫时,其生长受到明显的抑制。环境信息处理途径中有11个差异基因转录水平上调,其中与膜运输(Membrane transport)有关的上调差异基因为6个,与信号传导(Signal transduction)有关的上调差异基因为5个。高温胁迫下可能通过调节膜运输和信号传导等代谢途径,增加细胞膜通透性,维持细胞膜内外渗透压平衡,保持细胞膜的完整性,提高瑞士乳杆菌抗高温胁迫的能力。
高温胁迫会对瑞士乳杆菌的生长发育、生理过程产生多种不利的影响。细胞膜是分隔细胞与外界环境的一道屏障,也是细胞与外界进行物质运输和信息交流的重要通道[10]。在高温胁迫下瑞士乳杆菌的细胞膜流动性降低,膜通透性增加。细胞膜流动性和膜通透性主要与细胞膜的组成有关[23]。GO功能分类分析显示,细胞组分类别中,差异表达基因集中在细胞构造和含蛋白质复合物。KEGG通路分析显示,与膜运输有关的上调差异基因为6个。高温胁迫可能导致瑞士乳杆菌细胞膜成分发生变化,细胞膜流动性和细胞膜通透性发生改变,从而使瑞士乳杆菌对高温环境具有更好的适应性和保护性。
此外,在高温胁迫下瑞士乳杆菌的可溶性蛋白含量、海藻糖含量和SOD活性升高。GO功能分类分析显示,催化活性中有125个差异基因转录水平上调。这可能导致水解酶和合成酶相关基因转录水平上调,增加渗透调节物质(可溶性蛋白、海藻糖)的产生,提高其渗透调节的能力,维护细胞膜的正常生理功能,帮助细胞抵抗外界不良环境[10]。GO功能分类分析显示,与抗氧化活性有关的差异表达基因增加,可能增强瑞士乳杆菌清除活性氧的能力,防止膜脂过氧化,降低高温对细胞膜造成损伤。KEGG通路分析显示,高温胁迫会影响瑞士乳杆菌多条代谢途径,促进了氨基酸代谢、糖代谢和核苷酸代谢,为瑞士乳杆菌抵抗高温胁迫提供能量和组织细胞合成,从而增强细胞的代谢补偿效应,进而使瑞士乳杆菌对高温环境具有抗逆性。
3 结论
本文研究发现瑞士乳杆菌处于47 ℃高温胁迫时,其生长受到明显的抑制,细胞膜流动性降低,膜通透性增加。此外,在高温胁迫条件下瑞士乳杆菌的可溶性蛋白含量、海藻糖含量和SOD活性升高,维护细胞膜的正常生理功能,保护细胞抵抗外界不良环境。高温胁迫会影响瑞士乳杆菌多条代谢途径,主要促进了氨基酸代谢、糖代谢和核苷酸代谢,为瑞士乳杆菌抵抗高温胁迫提供能量和组织细胞合成,同时提高了调节膜运输和信号传导等相关基因代谢水平,从而增强细胞的代谢补偿效应,进而提高瑞士乳杆菌对高温胁迫的抗性。通过本研究对瑞士乳杆菌高温胁迫补偿效应机制的探讨,为进一步研究提高瑞士乳杆菌高温耐受性提供理论指导。
猜你喜欢
作文周刊·小学四年级版(2022年8期)2022-03-11
心肺血管病杂志(2020年5期)2021-01-14
心电与循环(2020年1期)2020-02-27
分析化学(2018年11期)2018-01-16
食品与生活(2017年12期)2018-01-09
江苏农业科学(2017年5期)2017-04-15
科学中国人(2015年13期)2015-02-28
世界热带农业信息(2014年6期)2014-09-12
湖北农业科学(2014年3期)2014-07-21
食品工业科技(2014年23期)2014-03-11
