
OsWD40过表达水稻在盐胁迫下的生理响应
2021-05-27 07:59柏华美黄梓轩郭敏鲍聆然沈波
浙江农业科学 2021年6期
柏华美,黄梓轩,郭敏,鲍聆然,沈波
(杭州师范大学 生命与环境科学学院,浙江 杭州 311121)
文献著录格式:柏华美,黄梓轩,郭敏,等.OsWD40过表达水稻在盐胁迫下的生理响应[J].浙江农业科学,2021,62(6):1076-1079.
DOI:10.16178/j.issn.0528-9017.20210607
土壤盐碱化是一个日益严重的全球性问题,其所造成的作物产量的损失对全球农业的威胁越来越大[1]。土壤中盐分过多阻碍了作物的生长,引起发育变化和代谢的适应,表现为细胞膜损伤、Na+和Cl-积累导致的离子失衡、脂质过氧化增强,活性氧如超氧自由基、过氧化氢和羟基自由基的产生增加[2]。水稻是盐敏感作物,盐胁迫也是制约水稻生产的主要逆境胁迫因子之一。多年以来,学者围绕水稻耐盐品种的评价及选育、农艺栽培措施、细胞信号响应机制、胁迫生理及相关基因的挖掘进行了广泛的研究,解析了水稻耐盐的内在机理[2-6]。随着转基因技术的不断成熟及新的耐盐基因的不断发现,通过转基因技术获得耐盐植株日趋成熟,加快了水稻耐盐种质改良的进程。Liu等[7]利用100 mmol NaCl处理转OsVP1和OsNHX1基因水稻幼苗3周后,发现转OsVP1和OsNHX1基因明显促进了水稻根系生长、水分吸收和叶水势,存活率分别为85.3%和100%,远高于野生型水稻的38.7%;而且,过量表达OsNHX1的转基因水稻可以在200 mmol NaCl胁迫下存活[8]。研究显示,转AtNHX1水稻植株在300 mmol高浓度NaCl处理下,能够正常存活,而野生型水稻5 d内几乎全部死亡,去除300 mmol NaCl胁迫条件的转AtNHX1水稻的恢复生长实验显示,转基因植株10 d内可以恢复正常生长,而野生型水稻则无法恢复,表明过表达AtNHXS1显著提高了水稻的耐盐性[9]。目前,利用基因工程技术培育耐盐性转基因水稻已经取得了较好的进展[10-12]。
盐胁迫制约了水稻的生长、发育,培育耐盐水稻品种已成为当务之急。本实验利用本实验室构建的转OsWD40水稻株系,进行盐胁迫处理,考察转基因水稻对盐胁迫的生理响应。
1 材料与方法
1.1 供试材料
OsWD40是本实验室通过生物信息学和同源克隆的方法,找到的一个响应NaCl的基因。在本课题研究中,以转OsWD40过表达T6代水稻株系(B3-5和B4-3)和受体日本晴(OryzasativaL. japonica.cv.Nipponbare)为实验材料。
1.2 处理设计
水稻种植采用国际水稻研究所(IRRI)水稻营养液配方配制水培溶液[13],经消毒、浸种、催芽、沙培后移入黑色小桶中水培,每3 d换一次全营养液。水稻在温室大棚中长至四叶一心时,用含200 mmol NaCl的水稻营养液处理水稻,在0、12、24、48 h对叶片取样,冷冻保存,用于生理生化指标测定。
1.3 指标测定
叶片相对含水量采用郭龙彪等[14]方法;叶片电导率使用DDS-307电导率仪测定;叶片叶绿素含量测定采用丙酮浸提法[15];叶片可溶性糖含量测定采用苏州科铭生物技术有限公司试剂盒;叶片总蛋白含量和过氧化氢酶(CAT)测定采用南京建成生物工程研究所试剂盒。所有数据测定均重复3次。
1.4 数据分析
所用数据为同一处理3次重复的平均值,采用SPSS软件的ANOVA过程处理,显著性检验采用邓肯式新复极差法。
2 结果与分析
2.1 对叶片相对含水量的影响
转基因水稻株系和日本晴的叶片相对含水量在处理开始时基本相同。随着胁迫时间的延长,水稻叶片相对含水量呈降低趋势,在处理后24、48 h时,转基因水稻B3-5和B4-3的相对含水量分别是64%、62%和39%、47%,均高于日本晴的50%和33%。其中24 h时B3-5的相对含水量、48 h的B4-3的相对含水量与日本晴相比呈显著差异,表明转基因水稻株系B3-5和B4-3叶片具有较好的保水作用(图1)。
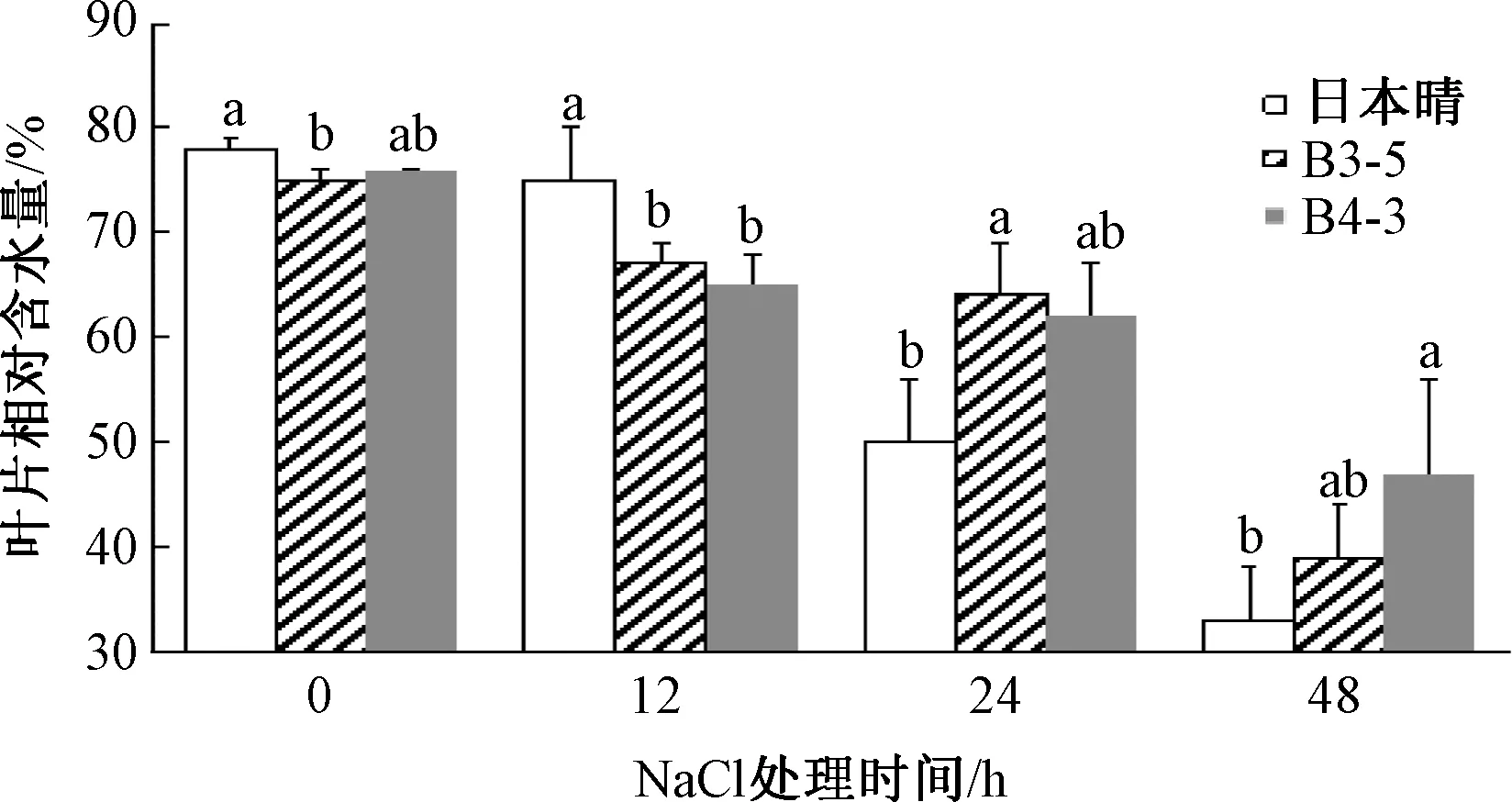
同一时间柱上,无相同字母表示处理间差异显著(P<0.05)。图2~7同。
2.2 对叶片相对电导率的影响
随着盐胁迫时间的增加,转基因水稻株系和日本晴的叶片电导率总体呈上升趋势。胁迫处理12 h时,B3-5、B4-3的叶片相对电导率分别是30.6%和34.5%,而日本晴为42.0%;24 h时,B3-5、B4-3的叶片相对电导率分别是45.0%和52.2%,而日本晴为60.3%;至48 h时,叶片细胞质膜严重受损,日本晴和2个转基因株系相对电导率均达最高值(图2)。说明在200 mmol NaCl盐胁迫处理下,在24 h时间范围内,转基因水稻株系B3-5和B4-3比日本晴具有较好的耐盐性,能够保持一定的细胞质膜功能。
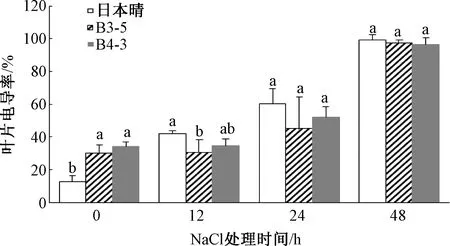
图2 NaCl处理对水稻叶片电导率的影响
2.3 对叶片叶绿素含量的影响
转基因株系B3-5、B4-3和日本晴在12 h时叶绿素a的含量都比未处理时高,说明此时在盐胁迫下水稻有一个适应过程,尚未影响叶绿素a的合成和积累。12 h之后3个品种的叶绿素a含量都呈快速下降的趋势。48 h时,转基因水稻株系的的叶绿素a含量显著高于日本晴,特别是B4-3品种(图3)。

图3 NaCl处理对水稻叶片叶绿素a含量的影响
叶绿素b含量的差异在盐胁迫24、48 h比较明显,特别是48 h时,转基因水稻株系的叶绿素b含量大于日本晴,其中B3-5的叶绿素b含量是日本晴的1.2倍,B4-3的叶绿素b含量是日本晴的1.6倍(图4)。实验结果表明,盐胁迫24、48 h时,转基因水稻株系与日本晴叶绿素a和叶绿素b的含量有较显著的差异。

图4 NaCl处理对水稻叶片叶绿素b含量的影响
2.4 对叶片总蛋白含量的影响
与胁迫开始时相比,盐处理12 h时的叶片总蛋白含量呈整体上升的现象。盐处理24 h时,日本晴水稻叶片总蛋白含量急剧下降,转基因水稻株系B3-5和B4-3的叶片总蛋白含量均显著高于日本晴,其中B3-5、B4-3叶片总蛋白含量分别是日本晴的1.9倍和2.2倍(图5)。盐处理48 h时,日本晴、B3-5、B4-3叶片总蛋白含量均有显著下降,叶片总蛋白含量分别为0.46、0.53和0.93 g·L-1,表明转基因水稻株系在盐胁迫的24~48 h仍然具有较好的蛋白质合成能力。

图5 NaCl处理对水稻叶片蛋白质含量的影响
2.5 对叶片可溶性糖含量的影响
随着盐处理时间的增加,日本晴和转基因株系B3-5、B4-3叶片的可溶性糖含量有增加,在12、24 h时,日本晴的可溶性糖含量均高于B3-5和B4-3。但盐胁迫48 h时,与日本晴相比,B3-5和B4-3的可溶性糖含量明显上升,分别增加了66.6%和37.1%(图6)。说明转基因水稻株系具有较持久的可溶性糖合成能力。

图6 NaCl处理对水稻叶片可溶性糖含量的影响
2.6 对叶片CAT活力的影响
在盐处理的不同时间,转基因株系B3-5和B4-3的CAT活性均高于日本晴,而日本晴随着盐胁迫时间进程,其CAT活性逐渐下降。从胁迫12 h到48 h,B3-5的CAT活性稳步增加,至48 h时,其CAT活性是日本晴的3.6倍;而B4-3的24 h CAT活性增加不明显,但48 h时,其CAT活性是日本晴的2.1倍(图7)。说明,转基因水稻株系B3-5和B4-3在盐胁迫条件下,可以保持较长时间的CAT活力来保持细胞机制的相对稳定的运行。

图7 NaCl处理对水稻叶片CAT活力的影响
3 小结
根据世界粮农组织估计,全世界超过6%的土地受到盐分的影响。如何在环境胁迫下提高作物产量,以应对日益增长的粮食需求,是研究人员急需解决的重要科学课题[16]。随着植物分子生物学的迅猛发展,耐盐基因的不断发现,通过转基因技术来获得抗盐植株已成为常规耐盐育种之外的一种有效手段。
目前,与盐分胁迫有关的生理指标如叶片相对含水量、电导率、抗氧化酶活性等,常被用于评价转基因植物对盐胁迫的抗性,这些生理指标主要反映了盐胁迫条件下细胞膜的损伤程度、氧化代谢水平及渗透物质的调节能力。实验表明,在盐胁迫条件下,ZFP179过表达水稻具有较高的相对含水量和存活率、较低的电导率和丙二醛(MDA)含量,并可积累比野生型更多的可溶性糖[17]。与野生型相比,在150 mmol NaCl处理后3 d,过表达OsCaM1-1的转基因水稻株系的相对含水量、相对生长率、光合色素含量以及过氧化氢酶和谷胱甘肽还原酶等活性均较高[18]。而转OsMYB6水稻150 mmol NaCl胁迫6 d后的恢复生长实验显示,野生型水稻全部死亡,转基因水稻仍有43.9%的存活率;生理指标显示,转基因水稻具有较低的电导率和MDA含量,较高的CAT和超氧化物歧化酶(SOD)活性[12]。通过这些生理指标的测定,可知转基因水稻的盐胁迫耐受能力,证实转基因水稻可以通过提高适应性的生理活动来增强水稻的耐盐能力[19-23]。
本研究表明,盐胁迫会影响转基因水稻的生长发育生理。在200 mmol NaCl处理条件下,与非转基因水稻相比,转基因水稻株系叶片细胞质膜损伤程度较轻,细胞锁水能力较强,表现为叶片相对含水量较高而电导率增加较缓;同时,转基因水稻株系通过自身对盐胁迫的响应,合成蛋白质、渗透调节物质及提高抗氧化酶代谢水平来改善对盐胁迫的耐受性,表现为总蛋白含量、可溶性糖含量及CAT活力的提高。
猜你喜欢
亚热带植物科学(2022年1期)2022-05-17
今日农业(2021年21期)2021-11-26
纺织科学研究(2020年1期)2020-02-25
四川农业科技(2019年5期)2019-07-01
浙江工业大学学报(2017年5期)2018-01-22
现代园艺(2017年23期)2018-01-18
中国塑料(2016年11期)2016-04-16
浙江柑橘(2016年1期)2016-03-11
化工进展(2015年6期)2015-11-13
食品工业科技(2014年15期)2014-03-11
