
江蓠和四角蛤蜊对珍珠龙胆石斑鱼封闭养殖水体水质的净化作用
2021-05-27 09:18:42王晓艳李宝山王际英王成强黄炳山刘义豪
烟台大学学报(自然科学与工程版) 2021年2期
王晓艳,李宝山,王际英,王成强,黄炳山,刘义豪
(山东省海洋资源与环境研究院山东省海水渔用饲料工程技术研究中心,山东省海洋生态修复重点实验室,山东 烟台 264006)
在自然生态系统中,大型海藻在环境修复和生态调控方面具有显著的作用,能有效控制和减少环境中的氮、磷等营养盐负荷[1-2]。鱼虾类摄食、代谢产生食物残渣和粪便等有机碎屑悬浮在水中难以沉降,滤食性贝类通过滤水、摄食、吸收、排泄一系列生理活动,以粪便或假粪的形式使较难沉积的悬浮物快速沉积下来,以更好地供大型海藻利用[3-4],大型海藻与异养型动物组成的生态系统,可实现营养物质有机和无机间的自然循环[5-7]。通过模拟自然生态系统,将鱼贝藻混养,利用滤食性贝类和大型海藻净化养殖水体[8-9],是实现海洋环境保护、渔业高质量发展和“蓝色碳汇”的重要途径。
江蓠属(Gracilaria)隶属于红藻门、真红藻纲、杉藻目、江蓠科,具有分布广、适应性强、生长快、产量高、易栽培等特点[10-11]。研究表明,江蓠具有快速吸收营养物质的能力,是环境修复的良好材料[12]。利用江蓠处理养虾池的废水,可以显著改善虾池水质,降低氨氮、硝酸盐、磷酸盐质量浓度,并提高虾和江蓠的产量[13]。四角蛤蜊(Mactraveneriformis)属于软体动物门、双壳纲、异齿亚纲、帘蛤目、马珂蛤科,是中国沿海分布极其广泛的一种双壳贝类,在过滤水中悬浮物方面具有很大潜力[1]。赖龙玉等[5]用38.13 g/m3的真江蓠与鲍混养,具有较好的营养盐吸收效果;赵素芬等[14]用500 g/m3的江蓠与鲍混养能保持水中较低的氨氮、亚硝态氮、磷酸盐的质量浓度;蔡泽平等[15]将石莼与真鲷(Pagrosomusmajor)复合养殖,在未充氧条件下,石莼能有效清除水体的营养盐,改良水质,改善真鲷栖息环境,提高真鲷的存活率和生长率。
本研究试验1使用江蓠(777.78 g/m3)与珍珠龙胆石斑鱼混养,探讨江蓠对养殖水体的原位修复效果,在确定利用江蓠修复养殖水体行之有效后,在试验1基础上设计试验2,探讨江蓠(1 555.56 g/m3)和四角蛤蜊(555.56 g/m3)联合使用对不同养殖密度的水体原位修复效果,并估算适宜养殖密度,以期为水产绿色生态养殖提供参考。
1 材料与方法
1.1 试验材料
养殖试验在山东省海洋资源与环境研究院东营实验基地海水养殖车间进行。试验用珍珠龙胆石斑鱼取自实验基地;江蓠购自福建省漳州市东山县海域,选取生长旺盛、大小均匀的藻体去除表面杂质后,4~8 ℃运送至实验基地;四角蛤蜊购自山东省东营市渔港码头,挑选无损伤、规格统一的个体(壳长23.53±1.33 mm, 壳宽26.33±1.83 mm,壳高16.00±0.95 mm,均重46.46±7.48 mg),并清除体表污物及附着生物,所有试验材料均置于同正式试验相同的条件下暂养1周。
1.2 试验设计与养殖管理
1.2.1 江蓠对珍珠龙胆石斑鱼封闭养殖水体水质的影响 试验1以珍珠龙胆石斑鱼单养为对照组,以珍珠龙胆石斑鱼与江蓠混养为试验组。试验开始前,用采水器采集养殖用水(取样量为500 mL),测定水体的初始硝酸盐(NO3--N)、亚硝酸盐(NO2--N)、氨氮(NH4+-N)、总氮(TN)、总磷(TP)质量浓度。选取规格统一、体质健壮、平均体质量为(714.00±7.07)g的珍珠龙胆石斑鱼50尾(养殖密度1 983.33 g/m3)分别放于对照组和试验组的工厂化水泥池(长6.0 m、宽6.0 m、水深0.5 m)中,次日试验组投放生长良好的江蓠14.00 kg(密度为777.78 g/m3),并于放藻前采集水样。对照组和试验组均每天定时(16:00)定量投喂一次,固定投喂量为该组鱼体初始总重的1%。试验期间每2 d于投喂前采集水样,控制水温26~28 ℃,溶氧>6 mg/L,水面光照1000 Lx,盐度27~29,试验1持续34 d。
1.2.2 江蓠和四角蛤蜊对珍珠龙胆石斑鱼封闭养殖水体水质的影响 试验2在试验1确定了江蓠净水效果的基础之上,进一步将不同密度的珍珠龙胆石斑鱼同生长旺盛的江蓠(28.00 kg,生物量为1 555.56 g/m3)混养,并在每个水泥池均混养长势良好的四角蛤蜊10.00 kg(555.56 g/m3),以期摸索生态养殖模式下珍珠龙胆石斑鱼的最适养殖密度。在相同溶解氧条件下,工厂化养殖每天排换水,珍珠龙胆石斑鱼的养殖密度在5000~8000 g/m3(数据参考山东省海洋资源与环境研究院东营实验基地珍珠龙胆石斑鱼养殖车间)。因此本研究选取1200尾规格统一,体质健壮、均重为475 g的珍珠龙胆石斑鱼,随机分为3个试验组,每组2个平行,每个平行分别投放100、200、300尾鱼(命名为D100、D200、D300,养殖密度分别为2 638.89 g/m3、5 277.78 g/m3、7 916.67 g/m3),每天定时(16:00)定量投喂一次,固定投喂量为该组初始鱼体总重的1%。试验开始前,用采水器采集养殖用水,测定水体的初始NO3--N、NO2--N、NH4+-N、TN、TP质量浓度。试验期间每7 d取一次水样,水样处理及养殖管理同“1.2.1”,试验2持续21 d。
1.3 测定方法
水体NO3--N、NO2--N、NH4+-N、TN、TP的测定及样品采集方法参照《海洋监测规范》(GB 17378—2007)[16],采集的水样立即用0.45 μm的滤膜过滤除去杂质,采用锌-镉还原法测定NO3--N,采用盐酸萘乙二胺分光光度法测定NO2--N,采用次溴酸盐氧化法测定NH4+-N,采用过硫酸钾氧化法测定TN和TP。
1.4 统计学方法

2 结 果
2.1 江蓠对珍珠龙胆石斑鱼封闭养殖水体水质的影响
2.1.1 江蓠的生物量变化和珍珠龙胆石斑鱼的存活率 本研究中,对照组珍珠龙胆石斑鱼在试验的第12天存活率降低至16%,而试验组在试验结束时(34 d)存活率为88%,江蓠生物量为12.91 kg,较试验初始值14.00 kg减少了7.79%。
如图1(b)所示,试验组水体ρ(NO2--N)的初始值为0.06 mg/L,放入珍珠龙胆石斑鱼24 h后,ρ(NO2--N)降低(0.03 mg/L),投入江蓠24 h后,ρ(NO2--N)进一步降低(0.02 mg/L),10 d以内均维持在较低水平(0.30 mg/L以内),第16~34天,ρ(NO2--N)在0.91 mg/L ~1.40 mg/L之间上下波动,而对照组在12 d以内呈直线上升趋势,到试验第12天,ρ(NO2--N)达到1.37 mg/L,而试验组在第24天时(1.40 mg/L)达到对照组第12天的水平。
如图1(c),对照组和试验组养殖水体ρ(NH4+-N)的初始值均为0.05 mg/L,随着饲料投喂及鱼体代谢,水体ρ(NH4+-N)呈逐渐升高趋势,第4天开始对照组超过试验组,到第12天,对照组达到1.29 mg/L,试验组为0.89 mg/L,16d后,试验组ρ(NH4+-N)变化趋于平稳,在第32天时(1.35 mg/L)达到对照组第12天的水平(1.29 mg/L)。
如图1(d)所示,试验组水体ρ(TN)的初始值为1.29 mg/L,放鱼之后,ρ(TN)略有升高(1.37 mg/L),投入江蓠后,ρ(TN)值出现短暂的降低(1.18 mg/L),之后整体呈现逐渐升高趋势,16~34 d在3.58 ~5.03 mg/L之间上下波动,而对照组ρ(TN)呈直线上升趋势,到试验第12天,达到5.01mg/L,试验组在第28天时(5.03 mg/L)达到对照组第12天水平。
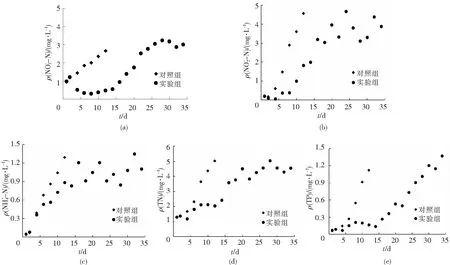
图1 江蓠对珍珠龙胆石斑鱼封闭养殖水体水质的影响
如图1(e)所示,试验组水体ρ(TP)的初始值为0.07 mg/L,放入珍珠龙胆石斑鱼后,略有升高(0.09 mg/L),放藻后,ρ(TP)水平下降(0.07 mg/L),之后到第14天,ρ(TP)水平较为稳定(0.14~0.21 mg/L),16天之后,ρ(TP)呈逐渐升高趋势,到试验结束(第34天)时,达到1.37 mg/L,对照组水体ρ(TP)呈直线上升趋势,到第12天,达到1.12 mg/L,与试验组第28~32天水平相当。
2.2 江蓠和四角蛤蜊对珍珠龙胆石斑鱼封闭养殖水体水质的影响
2.2.1 江蓠生物量和珍珠龙胆石斑鱼存活率 如表1所示,试验结束时,各组间江蓠末湿重的差异具有统计学意义(P<0.05),相比较初始湿重(28.00 kg),D100组江蓠生物量减少2.71%,D200组减少16.86%,D300组最低,生物量减少31.25%;D100组和D200组的四角蛤蜊末湿重之间的差异无统计学意义(P>0.05),但均显著高于D300组(P<0.05),D300组比初始湿重(10.00 kg)降低9.1%。各组间珍珠龙胆石斑鱼存活率的差异具有统计学意义(P<0.05),D100>D200>D300。

表1 江蓠生物量及珍珠龙胆石斑鱼存活率
2.2.2 江蓠和四角蛤蜊对珍珠龙胆石斑鱼封闭养殖水体NO3-N、NO2-N、NH4+-N、TN、TP质量浓度的影响 如图2(a)所示,D100、D200和D300组的水体的初始ρ(NO3--N)分别为0.98 mg/L、0.98 mg/L和0.97 mg/L,试验开始第7天,各组水体ρ(NO3--N)较初始值均有不同程度下降,之后均呈逐渐升高趋势,到试验第21天,D100、D200和D300组水体的ρ(NO3--N)分别达到第7天的5.91、3.54、4.40倍,D300组显著高于D100和D200组(P<0.05)。
如图2(b)所示,试验第7天,D100和D200组ρ(NO2--N)较初始值(0.064 mg/L, 0.062 mg/L)变化无统计学意义(P>0.05),D300组显著升高(P<0.05),随后3个处理组均呈不同程度的上升趋势,第21天时,D100、D200、D300组ρ(NO2--N)分别达到第7天的7.52、8.02、5.10倍,且D300组显著高于其他2组,D100组最低(P<0.05)。
如图2(c)所示,随着试验的进行,养殖水体的ρ(NH4+-N)浓度呈逐渐升高趋势直至试验结束,且养殖密度越大,水体ρ(NH4+-N)越高,D300>D200>D100(P<0.05)。
如图2(d),试验开始第7天,3个处理组水体的ρ(TN)较初始值均下降,且3组之间无统计学意义(P>0.05),7~21天3组均呈逐渐升高趋势,直至试验结束,且养殖密度越大,水体ρ(TN)越高,第21天时,各组ρ(TN)D300>D200>D100(P<0.05)。
如图2(e)所示,试验第7天,D100组ρ(TP)略有降低,D200和D300组的水体ρ(TP)略有升高,但3个组均与初始值(0天)之间差异无统计学意义(P>0.05),第14天,D100和D200组与第0和7天之间差异均无统计学意义(P>0.05),D300组显著高于D100和D200组(P<0.05),第7~21天,D100和D200组升高趋势较为平缓,D300组显著升高(P<0.05)。
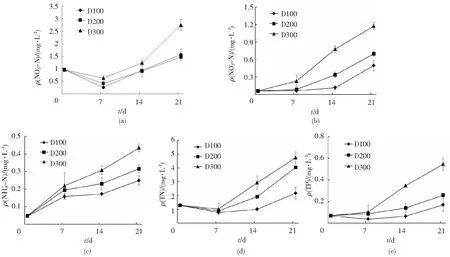
图2 江蓠和四角蛤蜊对珍珠龙胆石斑鱼养殖水体水质的影响
3 讨 论
3.1 江蓠对珍珠龙胆石斑鱼封闭养殖水体水质的影响
养殖鱼类代谢活动会产生大量的硝酸盐、氨氮、磷酸盐等代谢产物,残饲、粪便等也会造成水体中营养盐质量浓度增加,养殖废水若不经处理直接排放,会导致近岸海域有机质污染、水体富营养化[17]。大型海藻对水体富营养化的修复是一种安全可靠且具良好经济效益的原位生物修复方法,大量研究表明,江蓠属、海带属(Laminaria)、紫菜属(Porphyra)、石莼属(Ulva)、裙带菜属(Undaria)等大型海藻生长可有效吸收水体中过剩的N、P 等营养盐,改善水体富营养化状况[11]。温珊珊等[18]对真江蓠进行了分析,藻体N、P含量分别为4.645%和0.548%,表明每吨干重真江蓠可以从海区中除去46.5 kg N和5.5 kg P,可见大型海藻对于海区的生态环境具有非常显著的修复效果,且对N的去除能力强于P[19]。胡海燕[20]将大型海藻龙须菜与海水鱼类黑鲪(Sebastodesfuscescens)复合养殖,试验期间不换水,龙须菜自身生长状态良好,黑鲪的存活率与生长率均提高,该研究表明,龙须菜能有效降低黑鲪养殖水体溶解性N、P的含量。李志凌等[21]用500 g/m3的真江蓠与珍珠龙胆石斑鱼混养,各水质指标均优于珍珠龙胆石斑鱼单养组。在本研究试验1中,试验组将珍珠龙胆石斑鱼和江蓠混养,投入江蓠后,水体NO3--N、NO2--N、TN、TP质量浓度均有下降趋势,且能在10 d左右将各项指标维持在较低水平,表明江蓠具有短时间内储备营养物质的特点[22],之后各项指标升高,一方面可能是因为残饵、粪便等有机物不断积累,超过了一定量(14.00 kg,777.78 g/m3)的江蓠对水体N、P的吸收能力,另一方面可能是因为试验鱼对江蓠的摄食速度大于江蓠的生长速度,导致试验末江蓠重量(12.91 kg)低于初始重量(14.00 kg),从而使得江蓠对营养盐的吸收量低于营养盐积累量,在试验末期(20 d以后),试验组各项水质指标达到高水平,试验鱼开始出现死亡,试验结束时死亡率为12%,而未添加江蓠的对照组,在试验的第12天,鱼死亡率即达到84%,表明江蓠可以有效清除养殖水体的营养盐,改良水质,延长换水时间。
有研究报道,当介质中NH4+-N存在时会抑制大型海藻对NO3--N的吸收,有学者认为是由于藻类体内缺乏硝酸还原酶[23-24],也有学者认为是由于较NO3--N而言NH4+-N的结构简单,使得NH4+-N能迅速被转化成藻类氨基酸[13];另有研究报道,某些种类的海藻,即使在NH4+-N存在时也可以吸收NO3--N并且不受到抑制,某些海藻会出现NH4+-N和NO3--N同时吸收的现象[25]。而在本研究试验1中,水体NO3--N初始质量浓度较高(0.97 mg/L),投入江蓠后,水体ρ(NO3--N)显著降低,而ρ(NH4+-N)质量浓度没有降低趋势,反而逐渐升高,表明在本试验条件下,江蓠优先吸收NO3--N。
3.2 江蓠和四角蛤蜊对珍珠龙胆石斑鱼封闭养殖水体水质的影响
本研究试验2将江蓠和四角蛤蜊与不同密度的珍珠龙胆石斑鱼混养,利用江蓠和四角蛤蜊控制水体中的营养盐,水体各指标浓度与养殖密度呈正相关,养殖密度越大,营养盐质量浓度升高越快,且3个处理组的上升趋势均持续到试验结束(21 d)未出现降低,试验末江蓠生物量及试验鱼的存活率与养殖密度呈负相关,可能因为珍珠龙胆石斑鱼对江蓠摄食量大于江蓠的生长量,江蓠生物量的减少,使得江蓠对水体营养盐的吸收能力降低,而水体中累积过高的营养盐浓度,导致试验鱼存活率降低。
在氮素转化过程中,水体中残饵、粪便等有机物在短时间内被多种腐生细菌分解产生NH4+-N,NH4+-N在亚硝化细菌的作用下转化成NO2--N,亚硝化细菌的生长繁殖速度为18 min一个世代,NO2--N在硝化细菌的作用下转化成NO3--N,而硝化细菌的生长速度相对较慢,18 h一个世代,因此,由NO2--N转化到NO3--N的时间长很多[26]。在高密度养殖水体中,养殖动物生长依靠大量投饵,残饵、排泄物等有机物不断积累,水体产生大量的NH4+-N,大量积累的氮素硝化过程受阻,造成养殖水体中NH4+-N和NO2--N质量浓度高。相对于氨毒性,NO2--N对鱼类的毒性较小,但由于NH4+-N的转化速度较快,使得NO2--N的问题最为突出。养殖水体中离子氨允许的最高浓度不超过5 mg/L[27],本研究试验2中三个处理组在第21天时NH4+-N的质量浓度(分别为0.249 mg/L、0.316 mg/L、0.435 mg/L)均未达到最高限值5 mg/L。NO2--N的作用机理主要是通过呼吸作用,由鳃丝进入血液,与血红蛋白结合成高铁血红蛋白,血红蛋白的主要功能是运输氧气,而高铁血红蛋白不具备这一功能,从而导致养殖生物缺氧甚至窒息死亡,一般情况下,当水体中的NO2--N浓度达到0.1 mg/L,就会对养殖生物产生危害[28]。在本研究试验2中,D100和D200组的NO2--N浓度在第7天时均在安全范围内(分别为0.066 mg/L和0.087 mg/L),而在试验第14天,D100组(0.12 mg/L)达到安全限值,D200(0.34 mg/L)超出安全限值,D300组由于养殖密度过大,NO2--N积累过快,第7天(0.23 mg/L)即超过安全限值。比较试验1与试验2的D100组,试验1水泥池内鱼的生物量为35 700 g(每尾714 g,50尾),而试验2中D100组的生物量为47 500 g(每尾475g,100尾),在生物量大于试验1的情况下,试验2的D100组在第14天达到安全限值,晚于试验1的第6天,表明江蓠和四角蛤蜊搭配使用净化水质效果优于单独使用江蓠。
当环境中磷浓度较高时,某些藻类能大量吸收环境中的磷,将其储存在体内以维持低磷浓度时继续增殖,且藻类对磷的吸收速率与细胞内磷积累量成反比[29]。黄鹤忠等[30]研究表明,随着培养介质 N、P 浓度的不断提高,菊花江蓠对 N、P 的短期和长期吸收效率随之逐渐降低,且长期处于过高的 N、P 浓度环境中,还会破坏叶绿体等细胞器的结构。本研究试验1中,前14 d水体TP质量浓度均保持在较低水平(0.14~0.21 mg/L),14 d之后逐渐升高,可能因为江蓠在前期吸收了足够的磷维持自身生长,因此对磷的吸收率降低,也可能因为环境中的磷积累量超出一定量江蓠的吸收能力。
4 结 论
(1)在本试验条件下,使用江蓠(777.78 g/m3)修复珍珠龙胆石斑鱼(1 983.33 g/m3)封闭养殖水体,可以有效清除养殖水体的营养盐,改良水质;
(2)江蓠(1 555.56 g/m3)和四角蛤蜊(555.56 g/m3)联合使用可以有效净化养殖水体,珍珠龙胆石斑鱼的最适养殖密度为2 638.89 g/m3,最大不宜超过5 277.78 g/m3。江蓠、四角蛤蜊与珍珠龙胆石斑鱼建立生态循环系统,可以实现营养物质有机和无机间的自然循环,有效延长换水时间,降低工厂化养殖的生产成本,实现经济效益与生态效益双赢。在生产中,应该科学进行养殖管理,合理控制养殖密度,并根据鱼类对江蓠的摄食量,及时补充江蓠,保持江蓠和四角蛤蜊最佳净水能力。
猜你喜欢
当代水产(2023年2期)2023-05-19 08:23:02
连云港文学(2023年2期)2023-04-21 03:35:46
基层中医药(2022年7期)2022-11-17 08:25:02
作文小学高年级(2021年10期)2021-11-08 09:13:28
当代水产(2020年3期)2020-06-15 12:02:22
当代水产(2019年7期)2019-09-03 01:01:56
当代水产(2019年5期)2019-07-25 07:51:06
益寿宝典(2018年28期)2018-01-27 04:16:50
中成药(2017年12期)2018-01-19 02:06:43
中成药(2016年4期)2016-05-17 06:07:40
