
甘白种间杂交后代的形态学特征及细胞学分析
2021-05-25 05:20余青兰赵志刚
西南农业学报 2021年3期
殷 婷,赵 彤,余青兰,赵志刚
(青海省农林科学院/青海大学/青海省春油菜遗传改良重点实验室/国家油菜改良中心青海分中心/农业农村部春油菜科学观测实验站/青海省春油菜研究开发中心/青海省春油菜工程技术研究中心,青海 西宁 810016)
【研究意义】油菜作为重要的经济作物和油料作物,隶属十字花科芸薹属,主要包括了二倍体的白菜型油菜(Brassicarapa,AA,2n=2x=20)、异源四倍体的芥菜型油菜(B.juncea,AABB,2n=4x=36)和甘蓝型油菜(B.napus,AACC,2n=4x=38)。甘蓝型油菜具有抗病、抗倒、抗裂角、产量高、品质好等突出特性[1],在新中国成立后逐渐取代了之前的主栽种白菜型油菜和芥菜型油菜[2]。甘蓝型油菜遗传资源相对匮乏且自然界中遗传变异有限,在很大程度上限制了甘蓝型油菜的遗传改良。近年来,随着科学技术的进步与发展,育种家们想通过远缘杂交来拓宽甘蓝型油菜遗传基础,丰富其遗传多样性。【前人研究进展】远缘杂交虽然丰富了遗传资源,但也存在不亲和问题,从而导致了远缘杂交难以获得杂交种。远缘杂交的亲和性受环境、亲本基因型等的影响[3]。随着甘白种间杂交的广泛应用,其亲和性的研究也受到许多关注。赵彩霞等[4]对青藏高原白菜型油菜和甘蓝型油菜杂交亲和性研究结果表明:甘白杂交F1植株表型偏母本且自交亲和,不同杂交方式杂亲和性从大到小依次为白菜型油菜品种间杂交、(甘蓝型油菜×白菜型油菜)、(白菜型油菜×甘蓝型油菜),自交亲和性从大到小依次为甘蓝型油菜、F1(甘蓝型油菜×白菜型油菜)、F1(白菜型油菜×甘蓝型油菜)、白菜型油菜,之后何丽[1]和赵艳宁等[5]对甘蓝型与白菜型冬油菜杂交亲和性分析时也表现相同的规律。因此,甘白杂交时以甘蓝型为母本可提高杂交亲和性,同时通过甘白杂交对改良甘蓝型油菜和白菜型自交亲和性研究具有积极的意义。由于远缘杂交种集合了双亲的染色体,一般其表型介于双亲之间,个别性状偏向于某一亲本且雄性不育。何凤仙等[6]发现甘蓝型油菜与荠菜属间杂交后代外部形态偏向母本,部分有利性状偏向父本,少数有超亲性状。在孙龙涛[7]的研究中发现,黑芥与白菜杂种性状整体介于双亲之间,但个别性状偏向于母本。远缘杂交种中出现一些异常现象如父本染色体的消除、染色体渗入及不减数配子的结合等会产生偏母本或其他类型的植株。如牛妍[8]在进行芥菜型与白菜种间杂交时发现,芥菜型油菜×白菜型油菜的F1表型偏芥菜型,植株生长旺盛,分枝少,花粉完全不育,而白菜型油菜×芥菜型油菜的F1植株形态介于中间型,分枝基部丛生且较多。甘蓝型油菜由甘蓝和白菜杂交并加倍进化而来,其A基因组由二倍体祖先中白菜提供,在长期的遗传进化过程中发生了很大变化,同时甘蓝型油菜的A亚基因组相比C亚基因组遗传变异更丰富,二者杂交较易获得甘蓝型品系[9-11]。因此,白菜型油菜的A基因组遗传成分的利用对丰富甘蓝型油菜遗传变异极为有利。目前,通过甘蓝型油菜和白菜型油菜杂交获得不少优良的新型甘蓝型油菜,但缺乏对甘白杂交早期世代的细胞遗传学分析,而且在后代处理上,传统育种是采用与甘蓝型油菜多次回交,这样白菜型油菜只有少量的遗传成分导入甘蓝型油菜。【本研究切入点】本研究通过甘蓝型油菜与特早熟白菜型油菜种间杂交,利用形态学、细胞学手段对F1及自交或姊妹交F2、F3进行分析,揭示甘白杂种后代染色体数目变化趋势。【拟解决的关键问题】此结果为通过甘白杂交快速获得新型特早熟甘蓝型油菜提供细胞学支撑。
1 材料与方法
1.1 供试材料
甘蓝型油菜青油14号、AG 14和特早熟白菜型油菜浩油11号、门源小油菜分别于2017年9月5日和15日分批播种于青海省农林科学院春油菜研究所温室内。开花后选择青油14号、AG 14中生长健壮且无明显病虫害的单株,分别与特早熟白菜型油菜门源小油菜、浩油11号进行正反交,严格隔离和去除雄蕊,杂交获得F1。套纱帐获得F2,选择偏甘蓝型油菜的F2植株自交获得F3。
1.2 试验方法
1.2.1 杂交结角率及亲和指数 杂交时剥除母本的雄蕊,柱头上涂抹父本花粉后严格套袋,防止串粉污染。杂交后分枝挂上记有杂交方式、授粉花朵及杂交日期的纸牌。授粉后在成熟裂角前分别收获统计各分枝结角数、饱满种子数,计算结角率和亲和指数。
1.2.2 花粉可染率观察 晴朗天气,选取每株当天新开放的2朵花,将花粉涂抹在载玻片上,用醋酸洋红染色,盖上盖玻片,滤纸吸取多余染液,显微镜10倍镜下观察每朵花的3个视野,统计可染花粉粒占视野花粉粒总数的比率即花粉可染率。
1.2.3 有丝分裂观察 幼嫩花蕾置于离心管内,用冰盒取回实验室,解剖针挑取子房,置于8-羟基喹啉溶液中处理3~4 h,更换溶液为卡诺固定液(V无水乙醇︰V冰乙酸=3︰1)。镜检观察时,固定好的子房取出,置于1 mol/L 的HCl溶液,在60 ℃恒温水浴锅中水解6~8 min,蒸馏水冲洗后在载玻片上捣碎,滴上卡宝品红后盖上盖玻片,显微镜下观察并统计染色体数目。
1.2.4 减数分裂观察 幼嫩花蕾置于装好卡诺固定液的玻璃瓶中,更换固定液至花蕾褪色。镜检时,固定好的花蕾中剥取花药,置于1 mol/L HCl溶液,在60 ℃恒温水浴锅中水解3 min,蒸馏水冲洗后在载玻片上捣碎,滴上卡宝品红后盖上盖玻片,显微镜下观察染色体行为。
2 结果与分析
2.1 甘白杂交亲和性分析
以甘蓝型油菜×特早熟白菜型油菜为正交,进行正反交结实比较(表1)。正交试验中,青油14号×门源小油菜结角率为85.71 %,亲和指数为11.36;AG 14×浩油11号结角率为67.44 %,亲和指数为10.23。反交试验中,门源小油菜×青油14号结角率为95.38 %,亲和指数为6.03;浩油11号×AG 14结角率为81.51 %,亲和指数为3.66。通过对结实情况的研究发现4个杂交组合方式结角率在65 %以上,且正交组合亲和指数皆高于反交,正反交亲和性表现为(甘蓝型油菜×特早熟白菜型油菜)>(特早熟白菜型油菜×甘蓝型油菜)。

表1 甘白种间杂交亲和性Table 1 Crossing-compatibility index between Brassica napus and B.rapa
2.2 甘白杂种F1形态分析
根据田间植株的形态,对甘白杂种进行初步鉴定。田间形态调查发现青油14号×门源小油菜和AG 14×浩油11号的F1植株均大部分生长旺盛,分枝较多,表现出明显的营养生长优势,个别植株苗弱、死蕾、不育等,表现为劣势。杂种植株株高和花期介于双亲之间,但整体形态上偏向于母本甘蓝型油菜(图1)且花粉育性不佳。
2.3 甘白杂种F1细胞学观察
体细胞检测2个甘白杂交组合杂种F1植株染色体数均为2n=29,推测其基因组成为AAC(图2-A)。青油14号×门源小油菜的F1减数分裂终变期PMCs共观察了118个,染色体配对以二价体为主,二价体数目变化于10~14之间,出现多价体的PMCs为15.25 %(三价体15个,四价体2个,五价体1个);中期I的PMCs共观察了119个,96.64 %出现落后染色体,数目在1~5之间(图2-B);后期I/II的PMCs共109个,存在落后染色体的PMCs为20.18 %(图2-C),出现染色体桥的为1.83 %,后期I 有42.20 %的PMCs以15/14方式分离,其他的PMCs以16/13、17/12和18/11等方式分离;四分体时期的PMCs共观察了104个,以四分孢子为主,14.40 %的PMCs出现三分的异常分裂,还出现了3个五分孢子。
AG 14×浩油11号的F1减数分裂终变期PMCs共观察了97个,染色体配对以二价体为主,二价体数目变化介于10~14之间,也有23.71 %的PMCs出现多价体,其中三价体21个,四价体1个,五价体1个;中期I的PMCs共103个,97.09 %出现落后染色体,数目在1~6之间,分别分布在赤道板两侧;后期I/II的PMCs共观察了121个,14.88 %的PMCs出现落后染色体(图2-D),5.79 %出现染色体桥(图2-E),后期I主要呈15/14分离,占观察细胞总数的35.54 %,其余以16/13、17/12和18/11等方式分离;四分体时期的PMCs共观察了106个,以四分孢子为主(图2-F),4.72 %的PMCs出现三分的异常分裂,还出现了1个五分孢子。甘白杂种三倍体AAC减数分裂过程中,终变期染色体以单价体、二价体和多价体等形式存在及中期I出现染色体落后情况,导致染色体在后期I、后期II向两极移动时速度缓慢且方向不定,从而在后期I、后期II及四分体时期出现落后染色体、染色体桥等现象,这些染色体行为表明了A、C染色体组间具有一定的同源性,同时减数分裂的紊乱导致了F1花粉育性不佳。
2.4 F2细胞学观察
青油14号×门源小油菜杂交F2群体体细胞观察如表2所示:共观察97株,染色体数目介于20~58之间。2n=20的植株有2株,形态上偏白菜型,占总数的2.06 %,2n=23~27的植株15株,形态上介于双亲之间,占总数的15.46 %,2n=29~48的植株80株,形态上偏甘蓝型,占总数的82.47 %,其中5株2n=38,花粉可染率均大于80 %。
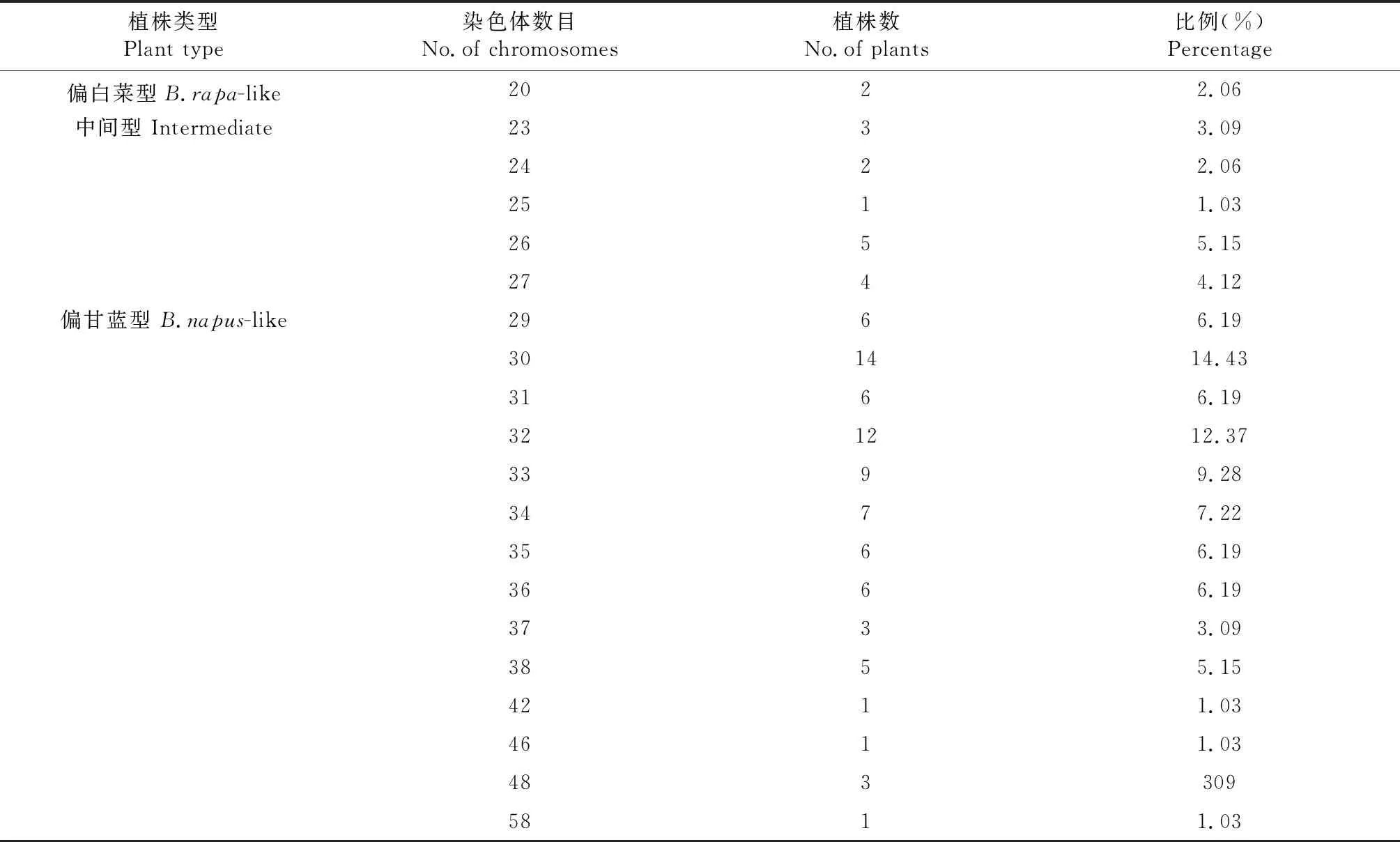
表2 青油14号×门源小油菜杂交F2群体染色体数目Table 2 No.of chromosomes with F2 between Qingyou 14×Menyuanxiaoyoucai 11
AG 14×浩油11号杂交F2群体体细胞观察如表3所示:共观察108株,染色体数目介于20~58之间。2n=20的植株有1株,形态上偏白菜型,占总数的0.93 %,2n=24~28的植株19株,形态上介于双亲之间,占总数的17.59 %,2n=29~58的植株88株,形态上偏甘蓝型,占总数的81.48 %,其中9株2n=38,花粉可染率在64.94 %~90.27 %。

表3 AG 14×浩油11号杂交F2群体染色体数目Table 3 No.of chromosomes with F2 between spring AG×Haoyou 11
青油14号×门源小油菜和AG 14×浩油11号杂交F2群体体细胞观察发现:染色体数目介于20~58之间。2n=20的植株形态上偏白菜型,2n=23~28的植株形态上介于双亲之间,2n=29~58的植株形态上偏甘蓝型,其中2n=38的植株较少。F2植株形态变异广泛,经细胞学观察发现偏白菜型和中间型植株2n=20~28,在选育甘蓝型油菜时可根据表型对其淘汰。
2.5 F3细胞学观察
在青油14号×门源小油菜和AG 14×浩油11号杂交的F2群体中,分别选取偏甘蓝型油菜,自交种子于2019年5月播种于试验田。根据叶型、株高等调查发现F3群体形态上均偏甘蓝型。在两个正交组合后代F3群体中,选择到具有38条染色体的植株20和6株,分别占27.40 %和12.77 %。如表4所示,2个组合中,2n=29~30的植株F3中2n=38的比例较低甚至没有,2n=31~34的植株后代中2n=38的比例约为0~26.67 %。总体上可以看出,F2植株染色体数目越高,其后代获得2n=38的植株比例越大。2n=38的F2植株F3中2n=38的比例约为46.67 %,仍有一半以上的F3植株没有达到38条染色体,这可能与F2中2n=38条的植株减数分裂终变期出现单价体和多价体、中期I及后期出现落后染色体和染色体桥有关。

表4 F3中染色体为38条植株Table 4 Somatic cell with 38 chromosomes of F3 plants
2.6 甘白杂交后代花期
高等植物开花这一现象是植株由营养生长转为生殖生长的重大转变过程。植株开花时间的早晚对其成熟期、产量、品质等有着一定的影响。油菜适宜的开花时间是保证高产、稳产的前提条件[14]。对各杂交后代的开花时间情况统计分析(表5),发现两个正交组合F1花期均介于双亲之间。青油14号×门源小油菜的F2偏白菜型植株中80.00 %的开花期比甘蓝型亲本早7 d,中间型的为30.00 %,偏甘蓝型的仅28.85 %,F3群体均为偏甘蓝型,开花期比亲本青油14号早7 d的占19.77 %;AG 14×浩油11号的F2偏白菜型植株中66.67 %的开花期比甘蓝型亲本早7 d,中间型的为31.43 %,偏甘蓝型的仅28.13 %,F3群体均为偏甘蓝型,开花期比亲本青油14号早7d的占8.16 %。由此可知:在甘蓝型油菜与特早熟白菜型油菜杂交后代中,F1植株花期介于双亲之间,F2群体中偏白菜型植株大都开花较早,而中间型、偏甘蓝型植株中早花植株的概率在依次降低,且F2偏甘蓝型油菜后代F3中早花植株更少,不足20.00 %。
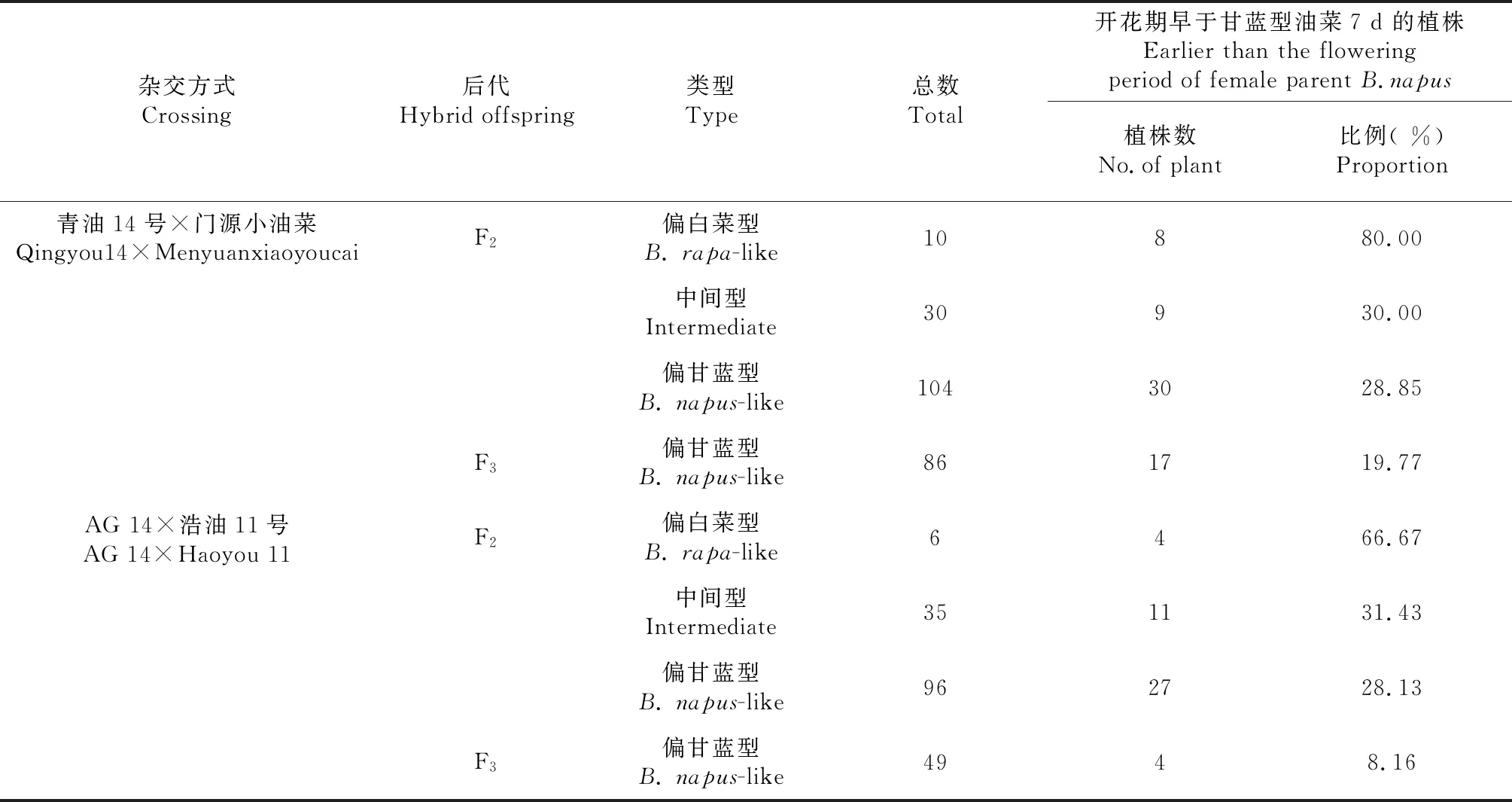
表5 甘白杂交后代开花期Table 5 Flowering period of hybridization pogenies between B.napus and B.rapa
3 讨 论
3.1 甘白杂交正反交组合的差异
远缘杂交由于亲本间基因型差异较大,普遍存在生殖隔离的障碍,具体分为受精不亲和、受精后胚胎发育障碍、杂种夭亡或不育等情况。芸薹属6个种间均可相互杂交,实现遗传物质的相互转移,但不同的物种之间杂交,其交配能力存在一定差异。前人研究表明甘蓝型油菜与白菜型种间杂交容易,通过常规种间杂交即可得到杂交种,后代中也易得到优良的甘蓝型油菜[9]。本试验通过3个甘蓝型油菜与2个特早熟白菜型油菜正反交,对杂交亲和性进行分析。从结实情况看,不同的甘蓝型油菜与特早熟白菜型油菜通过有性杂交,结角率达65 %以上,但双亲相同的正反交亲和性存在一定的差异,这说明甘蓝型油菜与特早熟白菜型油菜种间杂交亲和性受细胞质背景的影响。以特早熟白菜型油菜为母本的反交组合角果脱粒发现存在许多瘪籽,饱满籽粒少,初步推断特早熟白菜型油菜与甘蓝型油菜杂交存在着受精后胚胎发育的障碍。以甘蓝型油菜为母本的正交组合角果中多为饱满籽粒,较反交组合更易获得杂交种,亲和指数高,这与前人[15-17]的研究结果中以染色体数目多的亲本为母本获得真杂种概率高的结论相一致。
3.2 甘白杂交后代细胞学分析
甘白杂种F1形态均偏向于母本。有研究结果表明外源基因在后代这一新的遗传背景不一定能在形态上得以体现,孙万仓等[18]]认为植株偏母本是异源父本花粉诱导孤雌生殖所致。因此,对杂种真假的鉴定还需结合细胞、分子等方法进一步鉴定。甘白杂种F1花粉育性差,自交难以结实,可能是杂种含有2个十分相似但又有分化的芸薹属A基因组以及甘蓝型油菜的C基因组,在减数分裂过程中遗传物质发生重组,产生多种类型的配子所致,与Choudhary[19]研究观察B.juneca×toria、B.juncea×yellow sarson杂种F1花粉母细胞减数分裂形成不减数配子,导致花粉活力分别为16.5 %和20.4 %,花粉育性不佳相似。本研究甘蓝型油菜与特早熟白菜型油菜杂种F1体细胞具有预期的染色体数(2n=29),这与陈树忠等[20]对羽衣甘蓝和甘蓝型油菜杂种体细胞染色体数为双亲配子染色体数之和相一致。杂种F1染色体来源于甘蓝型油菜和特早熟白菜型油菜两个不同的物种,由于一些染色体同源性较低,在减数分裂中期配对不正常,形成单价体,而另一些染色体具有部分同源性,在减数分裂中进行同源配对,形成多价体,减数分裂后期单价体随机移向细胞两极,部分由于移动缓慢且方向不定而落后在赤道板上,出现落后染色体,这与Olsson[21]、牛妍[8]细胞学观察杂种染色体配对时存在单价体、二价体、三价体等相一致。减数分裂过程中单价体、多价体、落后染色体、染色体桥等的出现,说明存在A基因组间的同源配对、A基因组和C基因组以及C基因组内部部分同源配对情况,也伴随染色体的交换、缺失、易位、插入等的发生,表明甘蓝型油菜与特早熟白菜型油菜种间杂交实现了物种间遗传物质的相互渗透和重新组合,是获得新变异后代的理论基础。
由于甘白杂种F1染色体数目为双亲染色体的一半,说明每一个亲本只提供了相当于单倍体的基因组,双亲基因组间具有较大的差异,导致基因组内部分同源配对及基因组间部分同源配对,减数分裂终变期形成多价体,同源性较低的染色体形成单价体,从而出现落后染色体、染色体桥等异常现象,形成多种类型的配子。后代F2通过体细胞观察发现染色体数目存在明显差异,将表型与细胞学结合,表型偏白菜型油菜植株2n=20,中间型植株2n=23~28,偏甘蓝型油菜植株2n=29~58。与牛妍[8]的研究中芥白杂交后代2n=20的植株偏白菜型,2n=21~25的植株中间型,2n=26~28的植株偏芥菜型,染色体数目越多,形态越偏染色体数目多的亲本这一结果一致。偏白菜型和中间型植株2n=20~28,在选育新型甘蓝型油菜时,可以此初步给予淘汰。F2中2n=38的植株较少,且自交难以结实。F2群体偏甘蓝型植株自交后代F3全为偏甘蓝型,细胞学观察发现,F2中2n=35~38的植株自交后代中2n=38的植株达25 %以上且减数分裂单价体、三价体明显减少,但F2中2n=38的植株自交后代2n=38的比例并非100 %,可能是由于杂交后代中异源染色体的存在,使减数分裂过程中染色体配对时存在单价体、三价体以及后期分离出现落后染色体、染色体桥等,形成的配子染色体数目及组成存在差异,导致后代广泛分离。为减少甘白杂交选育甘蓝型油菜的中因群体过大造成的繁重工作量,可根据表型在田间初步淘汰偏白菜型和中间型的F2植株,对F2群体偏甘蓝型植株细胞学检测,选出2n≥35的植株自交选育新型甘蓝型油菜。
3.3 甘白杂交后代早花材料的获得
我国白菜型油菜品种资源丰富,栽培驯化历史长,分布广,具有耐瘠、抗旱、早熟、分枝多且分枝低等优良特性,且遗传资源丰富,是青海高海拔地区唯一能正常成熟的油菜栽培类型[8]。然而,白菜型油菜相比甘蓝型油菜产量低、杂种优势利用困难。甘蓝型油菜原产于欧洲,仅有400多年的历史,具有抗病、产量高、品质好、适应性强等特点,在新中国成立后逐渐取代了之前的主栽种白菜型油菜和芥菜型油菜[1]。然而,甘蓝型油菜没有野生种质资源,种内分化程度低,遗传基础在人为选择过程中渐趋狭窄,遗传资源匮乏,且原有春性甘蓝型油菜资源生育期较长,且在青海高海拔地区无法正常成熟,严重阻碍了甘蓝型油菜产区的增产增收和甘蓝型油菜的育种进程。开花时间对油菜成熟期、产量等有着一定的影响。甘蓝型油菜在青海等高海拔地区难以正常成熟,而作为高海拔地区主栽类型的白菜型油菜具有早熟的特点,但产量低。本研究在资源创新的亲本选择上,选择了春性AG 14以及青海省农林科学院育成的春性双低的早熟油菜青油14号。作为供体亲本的特早熟白菜型油菜选择了青海高海拔地区种植较为广泛的浩油11号和门源小油菜。通过甘蓝型油菜与特早熟白菜型油菜种间杂交,其后代F1表型偏甘蓝型,但花期介于双亲之间,早于甘蓝型亲本,晚于特早熟白菜型亲本。F2植株形态发生变异,主要分为偏白菜型、中间型和偏甘蓝型,且以偏甘蓝型植株为主。其中早花植株多为偏白菜型,偏甘蓝型且自交结实的早花植株少,鉴于甘白杂交主要目的是获得早熟的新型甘蓝型油菜,结合上述试验结果,偏向选择F2中偏甘蓝型的油菜植株自交获得F3。F3群体中存在开花较早的偏甘蓝型植株,但比F2群体偏甘蓝型植株中早花植株占比少,这可能与特早熟白菜型油菜遗传物质在后代中渗入多少有关。远缘杂交后代性状受父母本影响的程度不对等,虽能获得父本的某些性状,但其后代整体更偏向于母本,且随着世代的增加,后代群体中父本的遗传力更加弱化[18]。本研究甘白杂交后代中,特早熟白菜型油菜早花这一性状在后代群体中的表现情况再一次证明了该观点。
4 结 论
本研究通过对甘白杂交早期世代细胞学观察,发现(甘蓝型油菜×特早熟白菜型油菜)较(特早熟白菜型油菜×甘蓝型油菜)更易获得杂交种,且所得杂种F1染色体为双亲配子染色体数之和,PMCs减数分裂时存在异常行为。后代F2形态变异广泛,偏白菜型的植株2n=20,中间型植株2n=23~28,偏甘蓝型植株2n=29~58,其中2n=38的植株较少。偏甘蓝型的F2植株自交后代全为偏甘蓝型,其中2n=35~38的植株后代中2n=38的植株占25.00 %以上。花期调查发现F1花期介于双亲之间,F2中早花植株的概率降低,且F3中早花植株更少,不足20.00 %。
猜你喜欢
作物学报(2022年12期)2022-10-14
中国饲料(2022年5期)2022-04-26
作物学报(2022年4期)2022-02-10
作物学报(2022年4期)2022-02-10
作物学报(2022年3期)2022-01-22
散文诗世界(2019年10期)2019-09-10
科学之谜(2019年3期)2019-03-28
科学之谜(2018年8期)2018-09-29
农业科技与装备(2017年9期)2018-03-02
中学生理科应试(2016年4期)2016-11-19
