
Biomarkers in autoimmune pancreatitis and immunoglobulin G4-related disease
2021-05-25 10:01AkaneHaraTomohiroWatanabeKosukeMinagaTomoeYoshikawaKenKamataMasatoshiKudo
World Journal of Gastroenterology 2021年19期
Akane Hara, Tomohiro Watanabe, Kosuke Minaga, Tomoe Yoshikawa, Ken Kamata, Masatoshi Kudo
Abstract
Key Words: Biomarker; Autoimmune pancreatitis; Immunoglobulin G4 -related disease;Plasmacytoid dendritic cells; Cytokine; Chemokine
INTRODUCTION
Autoimmune pancreatitis (AIP) is a unique form of the chronic fibroinflammatory disorder of the pancreas, which is driven by autoimmune responses[1]. AIP is classified into type 1 and type 2 , and more than 95 % of AIP cases represent the former,which is a pancreatic manifestation of systemic immunoglobulin G4-related disease(IgG4 -RD)[2 -4 ]. In this article, type 1 AIP is hereafter referred to as AIP. AIP and IgG4 -RD are recently established disease entities proposed by rheumatologists and gastroenterologists[2 -4 ]. As awareness and recognition of these disorders by physicians increase, the number of patients diagnosed with AIP and IgG4 -RD is growing. Thus, the clinical manifestations and immunopathogenesis of AIP and IgG4-RD are attracting much attention from physicians and researchers.
IgG4 -RD occurs most commonly in elderly men; it is characterized by a marked elevation of serum IgG4 antibody (Ab) and accumulation of plasma cells secreting IgG4 Ab into injured organs[2 -4 ]. Another important feature of IgG4 -RD is multiple organ involvement: this disorder preferentially affects the pancreas, bile duct, lung,salivary glands, and kidney. AIP is a pancreatic manifestation of IgG4 -RD. The elevated concentration of serum IgG4 Ab is widely used as a diagnostic marker for AIP and IgG4 -RD[5 ,6 ]. In addition, patients with IgG4 -RD exhibiting multiple organ involvement display higher concentrations of serum IgG4 Ab[7 ,8 ], suggesting that measurement of serum IgG4 concentration is useful not only for the diagnosis but also for the evaluation of disease activity. It should be noted, however, that the concentration of this IgG subtype is not always regarded as a perfect biomarker for the diagnosis or evaluation of disease activity in AIP and IgG4-RD. In fact, serum concentration of IgG4 Ab is elevated in a significant fraction of patients with pancreatic cancer[9 ] and about 20 % of patients with AIP display normal serum concentration of IgG4 Ab[10 ]. Furthermore, patients with AIP sometimes relapse even if they have normal serum concentration of IgG4 Ab[11 ]. Therefore, it is necessary to identify other biomarkers that could be useful for the diagnosis and evaluation of disease activity in AIP and IgG4-RD.
Remarkable progress has been made in understanding the immunopathogenesis of AIP and IgG4-RD. Elucidation of immune networks associated with the development of these autoimmune disorders has led us to identify candidate biomarkers other than IgG4 Ab. In this Frontier article, we summarize recent progress in the biomarkers of AIP and IgG4-RD based on the knowledge of abnormal immune microenvironments.
IMMUNOPATHOGENESIS OF AUTOIMMUNE PANCREATITIS AND IGG4 -RD
Adaptive immunity
AIP and IgG4 -RD are characterized by enhanced IgG4 Ab responses; thus, immune microenvironments leading to IgG4 Ab production are likely to be involved in the development of these disorders[2 -4 ]. Various types of differentiated T cell subpopulations are involved in the enhanced IgG4 Ab response (Figure 1 ). These effector T cells include T helper type 2 (Th2 ) cells, regulatory T cells (Tregs), follicular helper T (Tfh)cells, and cytotoxic CD4 + T cells (CD4 + CTLs)[4 ]. Cytokines produced by effector T cell subpopulations promote IgG4 Ab production by B cells.
Interleukin-4 (IL-4 ), IL-10 , and IL-13 secreted by Th2 cells and/or Tregs promoted IgG4 Ab production by healthy control B cells in vitro[12 ]. In fact, expression of IL-10 ,IL-13 , and transforming growth factor-β1 (TGF-β1 ), also produced by Th2 and/or Tregs, was found to be higher in the livers of patients with IgG4 -RD than in the livers of patients with other autoimmune biliary diseases[13 ]. Moreover, the cytokine responses seen in IgG4-RD were accompanied by the enhanced expression of forkhead box P3 (FOXP3 ), a critical transcription factor for Tregs[13 ]. Koyabu et al[14 ] reported that the number of Tregs correlated with that of IgG4 + cells in the livers of IgG4 -RD patients. In line with the enhanced expression of Th2 and Treg-associated cytokines in the liver, patients with IgG4 -RD displayed higher expression of Th2 cytokines and chemokines such as IL-4 , IL-5 , IL-10 , C-C motif chemokine ligand 17 (CCL17 ), and CCL22 in the salivary glands as compared with the levels of these molecules in healthy controls and individuals with the Sjogren syndrome[15 ]. Such enhanced Th2 responses in the salivary glands were accompanied by TGF-β1 production and accumulation of FOXP3 + Tregs[15 ]. These data support the idea that Th2 cells and Tregs are involved in the development of AIP and IgG4-RD. Given that Tregs are potent negative regulators of autoimmune reactions, it might be possible that activation of Tregs is an epiphenomenon of persistent strong inflammation rather than a component of inflammation in AIP and IgG4-RD.
Ectopic germinal center formation is observed in the salivary glands of IgG4 -RD patients[15 ]. Tfh cells, which express B cell lymphoma 6 (BCL6 ) and C-X-C chemokine receptor type 5 (CXCR5 ), and produce IL-21 , play critical roles in germinal center reactions[4 ]. Expression levels of BCL6 , CXCR5 , and IL-21 in the ectopic germinal centers in the salivary glands were significantly higher in patients with IgG4-RD than in those with Sjogren syndrome[16 ]. Tfh cells isolated from the salivary glands and peripheral blood of patients with IgG4 -RD had a greater capacity to stimulate IgG4 Ab production by B cells than tonsillar Tfh cells[17 ,18 ]. The percentage of circulating Tfh2 cells, defined as CXCR5 + CXCR3 − C-C chemokine receptor type 6−cells, positively correlated with serum IgG4 concentration in patients with IgG4 -RD[19 ]. Thus, these data support the idea that Tfh cells are involved in the immunopathogenesis of IgG4-RD through the induction of germinal center reaction. However, the roles played by Tfh cells in AIP have not been clarified.
CD4+CTLs are a unique population of effector T cells that are often seen in patients with chronic viral infections. CD4 + CTLs are localized in the salivary glands of IgG4 -RD patients[20 ,21 ]. These cells express T-box-expressed-in-T-cells (T-bet) and produce interferon-g (IFN-γ)[4 ]. Although both Th1 cells and CD4+CTLs express T-bet and IFN-γ, they differ in expression levels of myeloid cell markers (CD11 b) and in the ability to produce CCL4 and IL-1 β[20 ,21 ]. In addition, these cells produce several cytotoxic proteins, including perforin and granzymes[20 ,21 ]. More importantly, these cells secrete TGF-β1 , one of the prototypical pro-fibrogenic factors. Thus, CD4+CTLs are involved in chronic fibroinflammatory responses associated with IgG4 -RD.However, accumulation of CD4+CTLs has not been verified in the pancreas of patients with AIP.
Innate immunity
Innate immunity is one of the major host defense mechanisms against microbial infections[22 ,23 ]. Recognition of microbial components by Toll-like receptors (TLRs)and nucleotide-binding oligomerization domain-like receptors (NLRs) induces proinflammatory cytokine responses to eradicate microbial infections[22 ,23 ]. It is well established that excessive innate immune responses underlie various types of autoimmune disorders[4 ,24 ]. Recent studies have highlighted the importance of innate immunity in AIP and IgG4 -RD (Figure 1 )[4 ].
We were the first to address the role of innate immunity in the development of AIP and IgG4 -RD. We initially examined whether peripheral blood mononuclear cells(PBMCs) isolated from IgG4-RD patients produced pro-inflammatory cytokines upon exposure to TLR ligands and found that they secreted more IgG4 Ab and Th2 cytokines than PBMCs from healthy controls[25 ]. In the subsequent studies, we utilized a co-culture system composed of peripheral blood CD14 + monocytes, CD19+B cells, and CD3 + T cells isolated from patients with AIP and IgG4 -RD. This co-culture system allowed us to show that B cells produced a large amount of IgG4 Ab in the presence of NLR and TLR ligands upon co-culture with monocytes isolated from patients with AIP and IgG4 -RD, but not with monocytes from healthy controls[26 ].Interestingly, peripheral blood monocytes isolated from patients with AIP and IgG4-RD efficiently induced IgG4 Ab production by B cells from healthy controls in a T cellindependent manner. Stimulation of TLRs and NLRs led to the production of B cellactivating factor (BAFF), which induced IgG4 Ab responses[26 ]. In addition to monocytes, peripheral blood basophils isolated from patients with AIP and IgG4 -RD also promoted IgG4 Ab production by B cells from healthy controls in a T cellindependent and BAFF-dependent manner[27 ]. These pioneering studies fully support the concept that excessive innate immune responses are involved in the development of AIP and IgG4 -RD. Indeed, the expression of TLRs was verified in the salivary glands and pancreas of patients with AIP and IgG4 -RD[28 -30 ].

Figure 1 Immunopathogenesis of autoimmune pancreatitis and immunoglobulin G4 -related disease. Plasmacytoid dendritic cells produce interferon-α and interleukin-33 and thereby mediate chronic fibroinflammatory responses in the pancreas. T helper type 2 cells expressing GATA-binding protein 3 ,regulatory T cells expressing forkhead box P3 , follicular helper T cells expressing B cell lymphoma 6 , and CD4 + cytotoxic T cells expressing T-box-expressed-in-Tcells are involved in the development of autoimmune pancreatitis and immunoglobulin G4 -related disease. IgG4 : Immunoglobulin G4 ; pDCs: Plasmacytoid dendritic cells; IFN: Interferon; IL: Interleukin; Th2 : T helper type 2 cells; Tregs: Regulatory T cells; GATA3 : GATA-binding protein 3 ; FOXP3 : Forkhead box P3 ; Tfh: Follicular helper T; BCL6 : B cell lymphoma 6 ; T-bet: T-box-expressed-in-T-cells.
It remains uncertain whether innate immune responses are shared by peripheral blood and affected organs in AIP and IgG4 -RD. To identify innate immune cells responsible for the development of AIP and IgG4 -RD, we utilized a murine experimental model of AIP and IgG4-RD. Repeated intraperitoneal injections of polyinosinicpolycytidylic acid [poly (I:C)] into MRL/MpJ mice led to the development of AIP characterized by the destruction of the pancreatic acinar architecture, immune cell infiltration, and fibrosis[31 ,32 ]. Thus, this murine experimental AIP model recapitulated pathological findings observed in human AIP. Extensive flow cytometry analyses revealed massive accumulation of plasmacytoid dendritic cells (pDCs),defined as PDCA-1 +B220 low, in the pancreas[31 ]. pDCs are a specialized DC population that produce type I IFNs (IFN-α) upon recognition of TLR7 and TLR9 ligands[33 ].Indeed, activation and accumulation of pDCs in the pancreas mediated experimental AIP through the production of IFN-α, because the depletion of pDCs or neutralization of type I IFN by Abs efficiently prevented the development of AIP[31 ]. Furthermore,pDCs expressing IFN-α and BAFF were found in the pancreas of patients with AIP and IgG4 -RD, and peripheral blood pDCs isolated from these patients promoted IgG4 Ab production by healthy control B cells in a type I IFN dependent manner[31 ]. Thus,these results strongly suggest that activation of pDCs and type I IFN production are prominent features of murine experimental and human AIP.
Although a unique form of fibrosis, called storiform fibrosis, is one of the characteristic findings in human AIP[2 -4 ], molecular mechanisms accounting for the induction and generation of this fibrogenic response have been poorly understood. We recently discovered that the type I IFN-IL-33 axis plays a pro-inflammatory and profibrogenic role in chronic alcoholic pancreatitis[34 ]. Type I IFN production by pancreatic acinar cells acts in concert with TNF-a produced by pancreatic macrophages to induce a robust production of IL-33 by the former cells[34 ]. Given that type I IFN produced by pDCs mediates experimental AIP, we hypothesized that IL-33 is involved in the generation of chronic fibroinflammatory responses in the pancreas.pDCs, which accumulate in the pancreas after repeated injections of poly (I:C),produced IL-33 in a type I IFN-dependent manner[32 ]. Importantly, the blockade of IL-33 -mediated signaling pathways by an Ab against the IL-33 receptor attenuated chronic fibroinflammatory responses of the pancreas, which was accompanied by a marked reduction in pro-fibrogenic cytokines such as IL-13 and TGF-β1 [32 ]. Immunofluorescence studies of pancreatic specimens from patients with AIP and IgG4 -RD confirmed pancreatic localization of pDCs expressing IL-33 [32 ]. Taken together, these results support the idea that activation of pDCs followed by the production of IFN-α and IL-33 mediates both experimental and human AIP. However, it should be noted that pDCs are not the only cellular source of IL-33 . For example, M2 macrophages have been shown to co-localize with IL-33 in the salivary glands of IgG4 -RD patients[30 ,35 ].
IMMUNOGLOBULINS AS BIOMARKERS IN AUTOIMMUNE PANCREATITIS AND IGG4-RD
The diagnosis of AIP and IgG4-RD relies on the detection of elevated serum concentration of IgG4 Ab as well as on the characteristic pathological findings, including abundant infiltration of IgG4 -expressing plasma cells, storiform fibrosis, and obliterative phlebitis[2 -4 ]. Thus, serum level of IgG4 Ab is widely used as an established biomarker for the diagnosis of AIP and IgG4-RD. Moreover, serum concentration of IgG4 declines rapidly after the induction of remission by prednisolone[11 ]. Indeed,serum concentration of IgG4 Ab was much higher in patients with AIP and IgG4 -RD than in individuals with chronic alcoholic pancreatitis and in healthy controls in our previous study[36 ]. Therefore, there is no doubt that measurement of serum IgG4 Ab concentration in clinical practice is necessary not only for the diagnosis of AIP and IgG4 -RD but also for the assessment of disease activity. However, IgG4 Ab level is not always informative for the diagnosis or assessment of disease activity in such patients.Around 20 % of patients with AIP have normal serum concentration of IgG4 Ab[10 ].Furthermore, a significant fraction of patients with pancreatic cancer also exhibit elevated IgG4 Ab concentration[9 ]. Moreover, disease flare-up is sometimes seen in patients with AIP and IgG4-RD, even on the background of normalized serum concentrations of IgG4 Ab[11 ].
IgG4 Ab is unique in that it has a limited ability to activate Fcγ receptors and complements[37 ]. Thus, IgG4 Ab is considered to play non-pathogenic rather than pathogenic roles in the development of AIP and IgG4 -RD. Shiokawa et al[38 ] directly addressed this issue by utilizing a passive transfer of patient IgG subtypes into neonatal mice. They found that pancreatic injury was successfully induced by a passive transfer of total IgG isolated from patients with AIP, but not by total IgG from healthy controls. The degree of pancreatic injury was much greater in neonatal mice treated with the IgG1 Ab from AIP patients than in those that received the IgG4 Ab from the same patients. Moreover, pancreatic injury induced by a passive transfer of IgG1 Ab was efficiently inhibited by a co-transfer of IgG4 Ab[38 ]. These studies conducted by Shiokawaet al[38 ] strongly suggest that IgG1 Ab rather than IgG4 Ab contributes to the immunopathogenesis of AIP and IgG4 -RD. Consistent with this idea, serum concentrations of both IgG1 and IgG4 Abs were significantly higher in patients with AIP and IgG4 -RD than in individuals with chronic alcoholic pancreatitis or in healthy controls (Table 1 )[36 ].
As for the other IgG subtypes, no significant differences were observed in serum concentrations of IgG3 Ab in patients with AIP and IgG4 -RD in comparison with those in patients with chronic alcoholic pancreatitis or in healthy controls[36 ]. Serum concentration of IgG2 Ab was significantly lower in patients with AIP and IgG4 -RD than in patients with chronic alcoholic pancreatitis (Table 1 )[36 ]. In contrast, another report showed that serum concentration of IgG2 Ab was elevated in patients suffering from IgG4 -RD[39 ]. This discrepancy can be explained by the difference in the organ distribution of IgG4 -RD. Serum concentration of this IgG subtype is preferentially elevated in patients with orbital IgG4 -RD, but not in those with pancreatic IgG4 -RD[36 ,39 ].

Table 1 Possible biomarkers in autoimmune pancreatitis and immunoglobulin G4 -related disease
Serum total IgG concentration is also elevated in patients with AIP and IgG4 -RD[2 -4 ]. Thus, elevations in serum concentrations of both total IgG and IgG4 are prominent features of patients with AIP and IgG4-RD. In contrast to augmented IgG and IgG4 levels, serum concentrations of IgM and IgA are decreased in patients with AIP and IgG4 -RD[40 ]. Furthermore, serum concentration of IgM inversely correlated with those of IgG and IgG4 (Table 1 )[40 ]. The diagnostic value of reduced serum concentrations of IgA and IgM needs to be determined in future studies.
Co-occurrence of AIP/IgG4 -RD and allergic disorders is often observed[2 -4 ]. In fact,serum concentration of IgE is significantly higher in patients with AIP and IgG4-RD than in those with chronic alcoholic pancreatitis and in healthy controls[36 ]. This raises the possibility that serum concentration of IgE can be used as a biomarker for AIP and IgG4 -RD. In line with this idea, approximately 50 % of patients with AIP and IgG4 -RD exhibit elevated serum concentration of IgE[41 ]. Moreover, changes in serum concentration of IgE are associated with the relapse of these disorders[41 ]. Therefore, serum concentration of IgE can be used as a biomarker for the diagnosis and prediction of relapse in AIP and IgG4 -RD[41 ].
AUTOANTIBODIES AS BIOMARKERS IN AUTOIMMUNE PANCREATITIS AND IGG4-RD
Although AIP and IgG4 -RD are considered to be caused by autoimmune reactions,autoAbs responsible for the development of autoimmunity have not been identified.Recently, three different types of autoAbs have been identified[42 -44 ]. These autoAbs recognize laminin 511 , annexin A11 , and galectin-3 (Table 1 ). Fifty-one percent of AIP patients were positive for autoAb against laminin 511 -E8 , a truncated variant of the extracellular matrix protein laminin 511 . Furthermore, serum IgG1 purified from AIP patients co-localized with laminin 511 in the pancreas of neonatal mice upon passive transfer[42 ]. Huber et al[43 ] identified annexin A11 , a calcium-dependent phospholipid-binding protein, as a candidate autoantigen in AIP. Interestingly, annexin A11 -specific IgG4 and IgG1 Abs purified from patients with AIP shared antigenic epitopes and IgG4 autoAbs inhibited pathogenic binding of IgG1 Ab to the shared epitopes[43 ]. These data suggest that IgG1 autoAbs rather than IgG4 autoAbs play pathogenic roles in the development of AIP and IgG4 -RD. Confirmation of these results awaits future studies that should address the diagnostic utility of these autoAbs in a large number of patients with AIP and IgG4 -RD. However, identification of autoAbs associated with AIP and IgG4-RD strongly supports the idea that these disorders arise from autoimmune reactions.
ADAPTIVE IMMUNITY CYTOKINES AS BIOMARKERS IN AUTOIMMUNE PANCREATITIS AND IGG4-RD
Effector CD4 + T cell subpopulations, including Th2 cells, Tregs, Tfh cells, and CD4+CTLs, are involved in the immunopathogenesis of AIP and IgG4-RD, as shown by the localization of these T cells in the affected organs. Th2 cells, Tregs, Tfh cells, and CD4+CTLs cells were detected in the peripheral blood of patients with AIP or IgG4 -RD[17 ,19 ,20 ,45 -48 ] and found to be markedly decreased after induction of remission[20 ,46 ], raising the possibility that serum concentrations of cytokines derived from effector T cells can be useful biomarkers (Table 1 and Figure 2 ). Yamamotoet al[49 ] found that serum concentration of IL-5 was elevated in patients with IgG4 -RD,whereas serum concentrations of IL-10 , IL-13 , IL-21 , and TGF-β1 were comparable in patients and healthy controls. Given that the major cellular source of IL-5 is Th2 cells,these data support the idea that the activation status of Th2 cells might be a surrogate marker for IgG4 -RD and AIP. In line with this notion, bile concentrations of Th2 cytokines such as IL-4 , IL-5 , and IL-13 were significantly higher in the patients with IgG4 -related sclerosing cholangitis than in those with primary sclerosing cholangitis[50 ]. Mechanistically, these Th2 cytokines induce bile leakage due to the impairment of the tight junction-associated biliary epithelial cell barrier, thereby causing chronic biliary inflammation[50 ]. Moreover, two cases of IgG4 -RD successfully treated with dupilumab that neutralizes IL-4 receptor α have been reported[51 ,52 ].Thus, Th2 responses may underlie the immunopathogenesis of AIP and IgG4 -RD.Therefore, Th2 cytokines, especially IL-5 , can be used as biomarkers of AIP and IgG4 -RD.
A positive correlation between serum concentration of IgG4 Ab and circulating numbers of Tfh cells has been demonstrated in patients with IgG4 -RD[17 ,19 ,46 ].However, the utility of serum concentration of IL-21 , a prototypical cytokine produced by Tfh cells, for IgG4-RD diagnosis has not been verified. Similarly, the usefulness of serum concentrations of IFN-γ and TGF-β1 produced by CD4+CTLs as biomarkers of AIP and IgG4-RD has not been reported either.
Therefore, at present, the utility of adaptive immunity cytokines as biomarkers for AIP and IgG4 -RD is limited. The reason why previous studies did not successfully identify adaptive immunity cytokines as biomarkers might be partially explained by a broad range of affected organs in AIP and IgG4 -RD or by complex effector T cell responses. Thus, the classification of IgG4 -RD into subtypes by affected organ distribution might lead to the identification of biomarkers specific for each such subtype.
INNATE IMMUNITY CYTOKINES AS BIOMARKERS IN AUTOIMMUNE PANCREATITIS AND IGG4-RD
Activation of pDCs and the subsequent robust production of IFN-α and IL-33 are characteristic pathogenic immune responses in experimental AIP and human IgG4-RD[31 ,32 ]. These findings led us to examine whether serum concentrations of IFN-α and IL-33 could be useful biomarkers for AIP and IgG4 -RD (Table 1 and Figure 2 ). For this purpose, we measured serum concentrations of these cytokines in patients with AIP and IgG4 -RD who met the well-established diagnostic criteria[5 ,6 ,36 ]. In comparison with the patients with chronic alcoholic pancreatitis and healthy controls,the patients with AIP and IgG4-RD displayed markedly elevated serum concentrations of IFN-α and IL-33 [36 ]. In contrast, serum levels of the prototypical proinflammatory cytokines IL-1 β and IL-6 were comparable in the patients with AIP/IgG4 -RD and those with chronic alcoholic pancreatitis[36 ]. Serum concentrations of IFN-α and IL-33 positively correlated with those of IgG4 Ab[36 ]. Thus, measurements of serum concentrations of IFN-α and IL-33 may be very useful for the diagnosis of AIP and IgG4 -RD[36 ].
We then evaluated the diagnostic performance of serum IFN-α and IL-33 concentrations for AIP and IgG4-RD. Surprisingly, the diagnostic performance of serum IFNα and IL-33 concentrations as diagnostic markers for AIP and IgG4 -RD was comparable to that of serum IgG4 Ab, as calculated by the receiver operating characteristic curve analysis[36 ]. Moreover, the induction of remission by prednisolone markedly reduced serum concentrations of IFN-α and IL-33 . Thus, serum IFN-α and IL-33 concentrations can be biomarkers that are useful not only for the diagnosis but also for the assessment of disease activity. Taken together, our data strongly suggested that serum concentrations of IFN-α and IL-33 might serve as novel biomarkers in AIP and IgG4 -RD. This idea is fully supported by an observation of an AIP/IgG4 -RD case in which serum concentrations of IFN-α and IL-33 were markedly reduced soon after the induction of remission, whereas those of IgG4 remained unchanged even after the successful induction of remission[53 ]. Identification of the IFN-α-IL-33 axis as a crucial pathogenic pathway as well as a biomarker leads us to speculate that patients with AIP and IgG4 -RD can be treated with biologics targeting IFN-α as in the case of systemic lupus erythematosus[54 ,55 ].
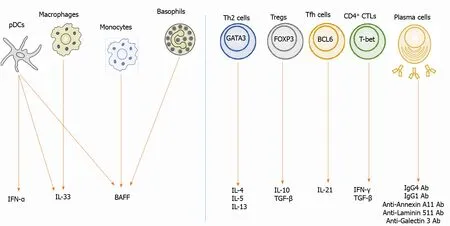
Figure 2 Biomarkers in autoimmune pancreatitis and immunoglobulin G4 -related disease. Left panel: Interferon-a (IFN-α) is produced by plasmacytoid dendritic cells (pDCs). Interleukin-33 (IL-33 ) is produced by pDCs and macrophages. B cell-activating factor (BAFF) is produced by pDCs, monocytes,and basophils; Right panel: IL-4 , IL-5 , and IL-13 are produced by T helper type 2 cells expressing GATA-binding protein 3 (GATA3 ). IL-10 and transforming growth factor-β1 (TGF-β) are produced by regulatory T cells (Tregs) expressing forkhead box P3 . IL-21 is produced by follicular helper T cells expressing B cell lymphoma 6 .TGF-β is produced by Tregs and CD4 + cytotoxic T cells (CTLs) expressing T-box-expressed-in-T-cells. IFN-γ is produced by CD4 + CTLs. Plasma cells produce immunoglobulin G1 (IgG1 ) and IgG4 Ab. These cytokines are possible biomarkers for autoimmune pancreatitis and IgG4 -related disease. IgG: Immunoglobulin G;pDCs:Plasmacytoiddendritic cells;IFN:Interferon;IL: Interleukin; BAFF:Bcell-activatingfactor;Th2 : T helpertype2 cells;Tregs:Regulatory T cells;GATA3 :GATAbindingprotein3 ;FOXP3 :Forkhead boxP3 ;Tfh:Follicularhelper T;BCL6 :Bcelllymphoma6 ;T-bet: T-box-expressed-in-T-cells;CTLs:CD4 + cytotoxicTcells.
BAFF and a proliferation-inducing ligand (APRIL) are cytokines produced by antigen-presenting cells[56 ]. BAFF and APRIL are crucial factors for B cell survival and thus, they promote Ig production[56 ]. Given that AIP and IgG4 -RD are characterized by elevated concentrations of serum total IgG and IgG4, in particular, it is likely that BAFF and APRIL are involved in the immunopathogenesis of these disorders. Indeed,pDCs producing BAFF have been demonstrated in the pancreas of patients with AIP and IgG4 -RD[31 ]. Moreover, T cell-independent class switch recombination of IgG4 Ab requires BAFF production by monocytes[26 ]. The involvement of B cell survival factors in the development of AIP and IgG4 -RD is further supported by the high probability of successful remission induction by rituximab in patients with IgG4 -RD[57 ]. As for the utility of BAFF and APRIL as biomarkers for AIP and IgG4 -RD,serum concentrations of BAFF and APRIL were significantly higher in patients with these disorders than in healthy controls[31 ,58 ]. In addition, induction of remission by prednisolone markedly reduced serum concentrations of BAFF[58 ]. Therefore,measurements of serum concentrations of BAFF and APRIL might be very useful not only for the diagnosis but also for monitoring disease activity.
IL-6 is a pleiotropic cytokine associated with autoimmune responses[59 ]. Although serum concentrations of this cytokine were comparable in patients with chronic pancreatitis, patients with AIP/IgG4-RD, and healthy controls in our study, elevated serum IL-6 level might help to discriminate a specific type of patients with AIP and IgG4 -RD. Tsukuda et al[60 ] compared clinical manifestations of patients with AIP and IgG4 -RD in relation to serum concentration of IL-6 . They found that hepatosplenomegaly and biliary tract involvement tended to be more prevalent in patients with high IL-6 serum level than in those with low IL-6 concentration[60 ]. However, we need to be cautious regarding the interpretation of these data, because hepatosplenomegaly is often seen in patients with multicentric Castleman disease, an IL-6-driven systemic autoimmune disorder[61 ]. Therefore, some patients with AIP and IgG4 -RD might exhibit clinical manifestations similar to those of Castleman disease.
These previous studies on innate immunity cytokines have opened up new research vistas that can facilitate identification of novel biomarkers in AIP and IgG4 -RD. In particular, serum concentrations of IFN-α and IL-33 , which faithfully reflect disease activity, may be informative diagnostic examinations in AIP and IgG4 -RD[36 ].
CHEMOKINES AS BIOMARKERS IN AUTOIMMUNE PANCREATITIS AND IGG4-RD
AIP and IgG4 -RD are characterized by Th2 responses[2 -4 ]. The prototypical Th2 chemokine, thymus and activation-regulated chemokine, also known as CCL17 , is a well-established biomarker for atopic dermatitis[62 ]. Given that atopic dermatitis and IgG4 -RD share Th2 responses, it is likely that serum concentration of CCL17 could be a useful biomarker for IgG4 -RD and AIP. Umeda et al[63 ] explored the utility of serum concentration of CCL17 as a possible biomarker for IgG4 -RD (Table 1 ). They found that serum concentration of CCL17 was significantly higher in patients with IgG4 -RD than in those with Sjogren syndrome or in healthy controls[63 ]. Although no association between serum concentrations of CCL17 and IgG4 Ab have been observed,those of CCL17 positively correlated with the number of affected organs. However, the utility of this chemokine as a biomarker for AIP has not been examined.
As mentioned above, production of IFN-α by pDCs is a prominent feature of AIP and IgG4 -RD. Excessive IFN-α responses result in the robust production of chemokines such as C-X-C motif chemokine ligand 9 (CXCL9 ) and CXCL10 [31 ]. In fact, the development of experimental AIP is accompanied by the enhanced expression of CXCL9 and CXCL10 in the pancreas[31 ]. The utility of CXCL9 and CXCL10 as biomarkers for AIP and IgG4-RD awaits future studies with the use of samples from patients with AIP and IgG4-RD.
CONCLUSION
AIP and IgG4 -RD are newly established disease entities[2 -4 ]. Both disorders are characterized by elevated serum concentration of IgG4 Ab and accumulation of IgG4 -expressing plasma cells in the affected organs[2 -4 ]. Moreover, the induction of remission by prednisolone is accompanied by a marked decrease in serum concentration of IgG4 Ab in patients with AIP and IgG4 -RD[2 -4 ]. Therefore, serum concentration of this IgG subtype is undoubtedly a useful biomarker for both the diagnosis and assessment of disease activity. However, a significant fraction of patients with AIP display active disease even at normal serum IgG4 Ab concentration. Recent elegant studies have shown that IgG1 rather than IgG4 plays the main pathogenic role in the development of AIP and IgG4 -RD[38 ]. Thus, elevated IgG4 Ab responses seen in AIP and IgG4 -RD are an epiphenomenon associated with chronic inflammatory reactions.Therefore, novel biomarkers based on the understanding of immunopathogenesis need to be established. We have recently found that pDCs producing IFN-α and IL-33 mediate experimental AIP and human IgG4 -RD[31 ,32 ]. Interestingly, serum concentrations of IFN-α and IL-33 have been identified as potent biomarkers for the diagnosis and assessment of disease activity in AIP and IgG4 -RD[36 ,53 ]. Based on recently identified biomarkers of these disorders, we propose diagnostic algorithm for patients with AIP and IgG4-RD exhibiting normal or slightly elevated concentrations of serum IgG4 Ab (Figure 3 ). As shown in Figure 3 , measurement of serum concentrations of cytokines and autoAbs in combination with serum IgG4 Ab might be useful for the diagnosis of AIP and IgG4-RD affluent in diversity.
As our knowledge of the immunopathogenesis of AIP and IgG4-RD increases, many candidate biomarkers will likely be identified in the future. The discovery of such biomarkers will contribute to the clinical practice and advance further our understanding of AIP and IgG4-RD immunopathogenesis.
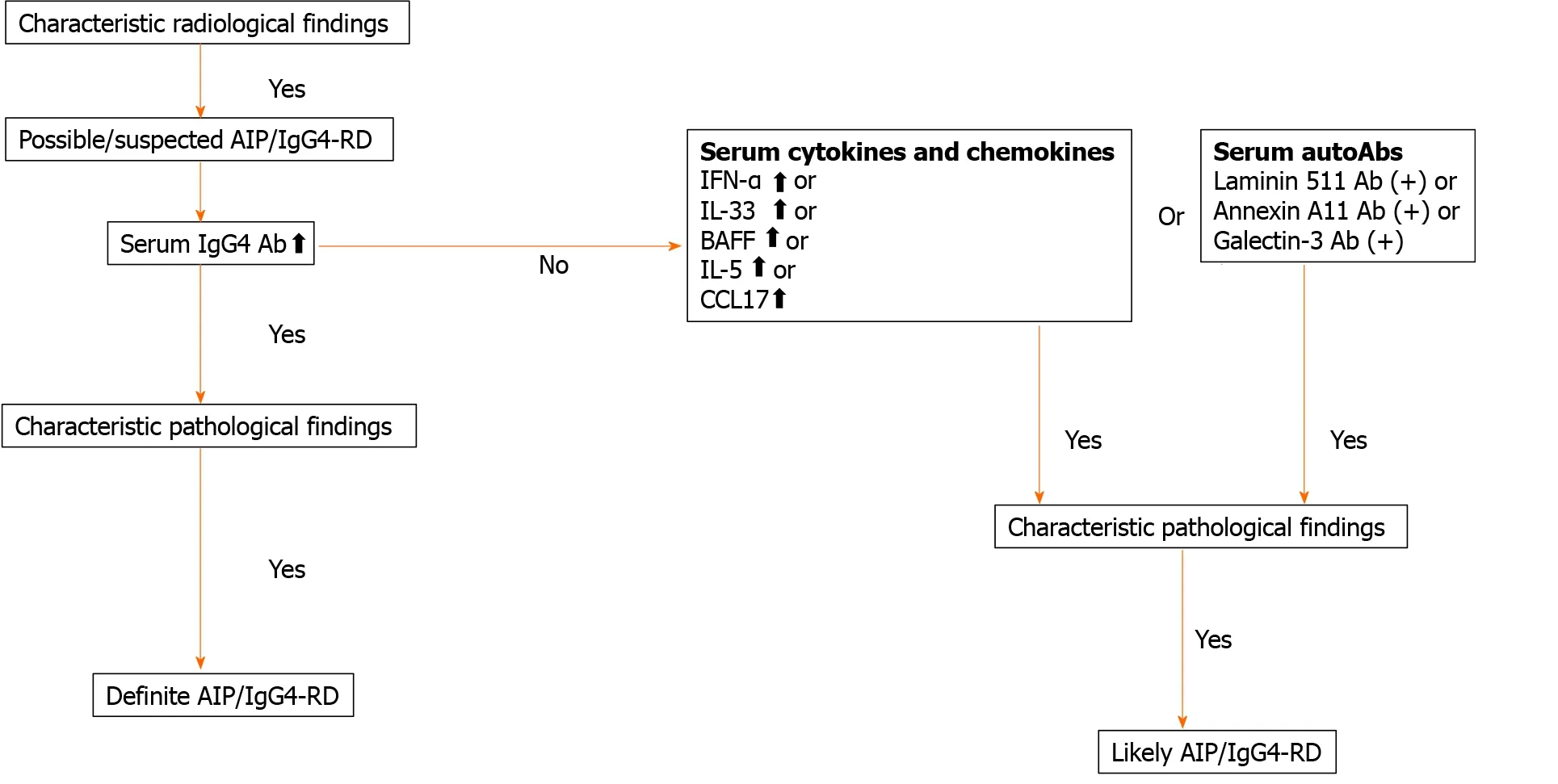
Figure 3 Diagnostic algorithm for autoimmune pancreatitis and immunoglobulin G4 -related disease. Serum concentration of cytokines and chemokines or the presence of serum auto-antibodies may be useful for diagnosis of autoimmune pancreatitis and immunoglobulin G4 (IgG4 )-related disease displaying normal or slightly elevated concentrations of IgG4 . AIP: Autoimmune pancreatitis; IgG4 -RD: Immunoglobulin G4 -related disease; IFN: Interferon; IL:Interleukin; BAFF: B cell-activating factor; CCL17 : C-C motif chemokine ligand 17 .
ACKNOWLEDGEMENTS
We would like to thank Ms. Yukiko Ueno for her secretarial assistance.
 World Journal of Gastroenterology2021年19期
World Journal of Gastroenterology2021年19期
- World Journal of Gastroenterology的其它文章
- Celiac Disease in Asia beyond the Middle East and Indian subcontinent: Epidemiological burden and diagnostic barriers
- Risk of hepatitis B virus reactivation in patients with autoimmune diseases undergoing non-tumor necrosis factor-targeted biologics
- Risk factors and prognostic value of acute severe lower gastrointestinal bleeding in Crohn’s disease
- Changes in the nutritional status of nine vitamins in patients with esophageal cancer during chemotherapy
- Effects of sepsis and its treatment measures on intestinal flora structure in critical care patients
- Gut microbiota dysbiosis in Chinese children with type 1 diabetes mellitus: An observational study