
高寒冻土区生物结皮对土壤理化属性的影响
2021-05-24 02:31金会军杜玉霞
冰川冻土 2021年2期
明 姣, 盛 煜, 金会军, 张 泽, 杜玉霞
(中国科学院西北生态环境资源研究院冻土工程国家重点实验室,甘肃 兰州 730000)
0 引言
生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1]。其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3]。近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4]。目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10]。国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16]。我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17]。黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一。源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18]。然而,20 世纪70 年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20 世纪末,植被退化面积占源区草地总面积1/3[19]。2005 年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育。笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布。黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知。生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22]。而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10 月初至次年4 月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态。在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰。
为此,本文以青藏高原黄河源区玛多县附近的生物结皮为研究对象,借助野外调查采样和室内试验方法的结合,来进行定量研究,以探知高寒冻土区内存在的生物结皮对土壤理化属性的影响,为深入研究高寒地区生物结皮生态功能提供理论基础。
1 研究区概况与研究方法
1.1 研究区概况
黄河源区玛多县隶属果洛藏族自治州,地理位置为95°30′~103°30′E,32°30′~30°30′N,位于青海省南部,巴颜喀拉山北麓,土地面积12.2×104km2。该地属于平原地区,平均海拔4 200 m。植被类型以草地为主,约占88%。区域气候类型属于青藏高原亚寒带半干旱半湿润型气候[24-25],昼夜温差大,年平均气温-1.6 ℃;年内降水多集中在5—9 月,年内累积降水量407~582 mm;年累积蒸发量为484~584 mm[25]。该地区特有的地理位置和地形、地貌、水文、干寒的气候条件决定了该区为季节冻土区,并镶嵌着不连续多年冻土、零星高温多年冻土和季节冻土[26-27]。我们的采样观测场地位于玛多县野马滩,位于38°34′N,98°00′E,海拔4 320 m,属于季节冻土区,研究场地景观如图1所示。

图1 研究区地表生物结皮状况(样带A为浅色藻结皮,B为深色藻结皮,C为裸地)Fig.1 Biocrusts in the study region:light cyanobacteria biocrust(A);dark cyanobacteria biocrust(B);bare soil(C)
1.2 研究方法
1.2.1 野外调查及采样
在玛多县野马滩禁牧场地内每隔2 km 分别选取分别选取人为干扰较少、地形地貌呈现较高一致性、生物结皮发育稳定的5~6 块(每块400 m2)的禁牧地作为调查样地。每个样地中随机设定4 个5 m×5 m 的调查样方(大样方),每个大样方中随机选择10 个25 cm×25 cm 的小样方,调查生物结皮物种组成、盖度及样地植被等。然后在每个样地中,分别选择两种(浅色藻结皮和深色藻结皮)不同类型生物结皮斑块与裸土斑块(5 m×5 m),随机选取4个点,用铲子剥离结皮层和下层的分层处土壤,作为结皮层土壤样品。再依次采集下层分层土壤,即0~5 cm、5~10 cm、10~20 cm、20~30 cm 的土壤样品,4点同层混合作为1个分析样品,即每个斑块类型采集5~6 份土壤样品。采集到的样品经过风干后,剔除下层土壤中肉眼可见的植物残渣,磨细过筛备用。样点的具体情况见表1。
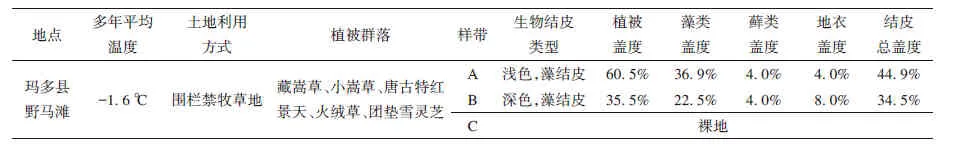
表1 土壤采样点基本情况Table 1 Characters of sampling sites
1.2.2 测定项目及方法
(1)生物结皮覆盖度
采用25 点样点法(小样方的长宽均为25 cm),在设置的每个5 m×5 m 的样方内,以之字状沿对角线设定10 个小样方,调查生物结皮种类和盖度,同时记录样方中苔藓、地衣、藻、裸土、小石砾、高等植物等出现的频率,以各物种与调查总点数的百分比表示其相应覆盖度。同时,调查记录下采样点的海拔、经纬度及植被状况等。
(2)土壤物理性质
土壤颗粒组成:由马尔文MS2000 激光粒度分析仪测定。由于生物结皮层土壤有机质呈现出较大含量,为了达到分散土壤颗粒的目的,进行样品先期处理,处理方式如下:用不同浓度(先用10%浓度,后用30%浓度)的过氧化氢(H2O2)溶液浸泡土壤,达到去除有机质的目的。然后剔除土壤中携带的碳酸盐,剔除过程用到了0.20 mol·L-1的HCl 溶液进行土壤的浸泡,最后用0.05 mol·L-1的HCl 溶液浸泡,直到没有气泡出现才结束土壤的浸泡,再进行上机测试。
田间持水量:将原装生物结皮放在具孔小筛上(孔径为1 mm),然后把结皮连同小筛一起,放在盛薄层的托盘上,当结皮充分饱和时,将结皮和小筛取出来,放置在定性滤纸(5层),吸水5 min 后,进行结皮湿重的称量,后再将生物结皮的烘干,并称量得到其干重,计算质量含水量,以此作为相对田间持水量[28]。
容重:结皮层容重与生物结皮层下土壤容重的测定采取的方法分别是涂膜法[29]和环刀法。
(3)土壤化学性质
全氮(TN)用开氏法测定[30],有机质(SOC)使用重铬酸钾氧化法测定[31],pH值用电极法测定[32-33]。
1.2.3 数据处理
借助于Origin 9.0 和SPSS 17.0 完成数据处理和统计分析,针对各种类型的生物结皮及裸地土壤导热系数及其理化属性分别进行单因素方差分析,用LSD 法进行多重比较,处理间差异均达5%显著水平。
2 结果与分析
2.1 高寒地区生物结皮发育特征
随着三江源地区禁牧工程实施,生物结皮在黄河源区大面积发育,在黄河源区玛多县调查结果表明,该地主要以藻类结皮为主,生物结皮总覆盖度可达37.3%~51.7%,厚度为6~22 mm,平均厚度为12.6 mm。生物结皮中各组分所在百分比如表1 所示。根据其鲜明的颜色特征,分为浅色藻类结皮和深色藻类结皮,分别如图1(a)和1(b)所示。生物结皮物种分布与植被密切相关,在未退化的草地斑块中,植被主要有藏嵩草(Kobresia tibetica)和团垫雪灵芝(Arenaria pulvinata),主要以深色藻结皮为主,其厚度较厚为6~22 mm(平均13.7 mm);在半退化的草地斑块中,主要植被为垂头菊(Cremanthodium reniforme),该区域则生物结皮以浅色藻结皮为主,其厚度为6~19 mm(平均11.5 mm)。
2.2 生物结皮对土壤物理性质的影响
研究区内两种类型生物结皮与对照组裸地及下层(0~30 cm)土壤粒级分布如图2所示,按照美国制颗粒分级标准,将整个粒径分为黏粒(<0.002 mm)、粉粒(0.002~0.050 mm)和砂粒(0.050~1.000 mm)。由图2 可见,两种生物结皮及对照组裸地各层土壤的颗粒组成均以粉粒(0.002~0.050 mm)和砂粒(0.05~1.00 mm)为主,占到所有颗粒组分的70.0%~80.0%左右,而黏粒(<0.002 mm)仅占25%左右。两种类型生物结皮结皮层黏粒、粉粒及砂粒均没有显著差异。两种类型结皮层下0~5 cm 土层土壤黏粒、粉粒和砂粒含量与裸地相比也没有显著性差异。5~10 cm、10~20 cm 土层中粉粒含量有所不同,表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著。而深色藻结皮覆盖下20~30 cm 土层中粉粒含量高于浅色藻结皮,同时均显著高于裸地。生物结皮对土壤颗粒含量的影响作用显著表现在20~30 cm 土层,这可能与青藏高原强烈的冻融作用及水热迁移有关。
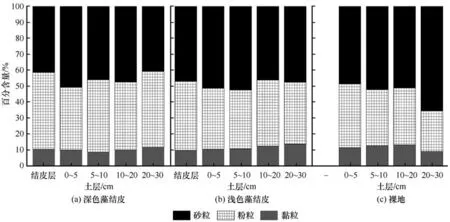
图2 不同类型地表覆盖下土壤颗粒组成Fig.2 Soil particle composition of two types of cyanobacteria biocrusts and bare soil
土壤容重是反映土壤结构、透水性能以及保水能力的一个指标,其与土壤密度、孔隙以及有机质含量等因素有关。因此生物结皮的形成可能影响了土壤孔隙度以及土壤有机质含量等,从而可能影响土壤容重的大小。不同土层土壤在生物覆盖和无生物结皮覆盖情况下各土层土壤容重变化情况如图3所示。由图3 可见,研究区域深色藻结皮和浅色藻结皮层容重分别为1.15 g·cm-3、1.08 g·cm-3,均显著低于下层0~30 cm 土层土壤容重(1.65 g·cm-3)。深色藻结皮覆盖下0~20 cm 土层土壤容重显著低于浅色藻结皮和裸地,浅色藻结皮覆盖下0~20 cm 土层土壤容重与裸地没有显著差异。20 cm 土层两种类型生物结皮和裸地土壤容重均没有显著差异。
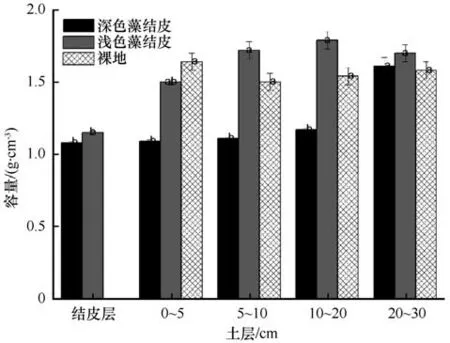
图3 不同土层土壤容重随地表覆盖变化Fig.3 The soil bulk density of different soil layers with different land cover
土壤田间持水量可反映土壤保水能力。由于生物结皮的发育,其结皮层孔隙度、容重及其他指标较下层土壤有显著变化。因此生物结皮层的保水能力显著影响下层土壤水分传输。故测量结皮层这一特殊结构与裸地表土(2 cm)土壤田间持水量变化情况见图4。由图可知,深色藻结皮结皮层田间持水量为60.3%,显著高于浅色藻结皮层(45.0%)和裸地表层(41.2%)土壤田间持水量,浅色藻结皮层土壤田间持水量略高于裸地,但两者之间差异不显著(P<0.05)。

图4 不同藻类生物结皮层及裸地表层土壤田间持水量Fig.4 The field water holding capacity of two kinds of cyanobacteria biocrusts and bare surface soil
2.3 生物结皮对土壤养分含量的影响
土壤有机质、全氮及酸碱度是评价土壤质量的重要化学指标。表2为不同类型生物结皮及裸地各层土壤有机质、全氮含量及pH 值。由表中可见,深色藻结皮结皮层土壤有机质和全氮含量与浅色藻结皮没有显著差异。两种类型生物结皮结皮层土壤有机质和全氮含量均显著高于0~20 cm 土壤层。0~20 cm 土层土壤有机质和全氮含量为深色藻结皮覆盖下含量最高,浅色藻结皮次之,裸地最低,且三者之间含量差异显著。生物结皮的形成显著影响了土壤pH大小。两种类型生物结皮结皮层土壤pH值均显著低于0~20 cm 土层土壤pH 值,而在0~20 cm 土层中,两种类型生物结皮之间土壤pH 没有显著性差异。结皮层pH 略低于裸地,但与裸地之间相比,差异性也不显著。

表2 不同类型生物结皮各土层土壤养分含量Table 2 Soil total nitrogen content in different soil layers of different types of biocrusts
3 讨论
生物结皮可以在各种气候条件下广泛发育[1],国内外关于生物结皮的研究主要集中在荒漠地区及干旱半干旱的黄土高原等地区[4-16],关于青藏高原高寒冻土区生物结皮的发育特点鲜有报道。本研究结果表明冻土区生物结皮主要是以藻为主的生物结皮,结皮层厚度大约为6~22 mm。其在厚度上大于干旱的荒漠地区藻类结皮厚度(1~5 mm)[16]。这与该地区特殊的生态环境条件密切相关,由于冻土区常年温度低,生物结皮活性较弱,呼吸等生理活性缓慢[4,21],其中积累的物质不易分解,因此,生物结皮层厚度较大。同时,生物结皮的厚度与生物结皮的发育年限密切相关[29]。因此,该区域植被退化区域生物结皮(浅色藻结皮)较未退化区生物结皮(深色藻结皮)发育年限短,故其深色藻结皮厚度大于浅色藻结生物结皮小。
生物结皮对土壤理化属性具有显著影响[1,3,14-15,28]。关于生物结皮对土壤颗粒组成的影响,在不同的研究区域所得的研究结论存在一定分歧。有些研究结果表明,生物结皮的存在,能够细化土壤颗粒[28,34-35]。也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36]。本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm 土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著。来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势。在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28]。这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm 左右,细颗粒呈现明显的表聚现象[28]。而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm 土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关。由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm 土层。这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化。与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm 土层土壤容重。而浅色藻结皮则对下层土壤容重影响不明显。本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重。生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地。
生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21]。生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38]。同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力。虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40]。本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm 土层土壤有机质含量。藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41]。同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42]。该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异。同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm 土层土壤有机质含量。然而,两种生物结皮对土壤全氮含量的影响则有所不同。深色藻结皮的形成同时也提高了结皮层及其下0~20 cm 土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm 土层土壤全氮含量没有显著影响。生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6]。生物结皮在湿润后,其固氮活性强烈受制于水分[43]。以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多。氮素在水热交换作用下,向下层输入。而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层。同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH 并没有显著影响。由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH 降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小。高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究。
4 结论
本文初步研究了青藏高原黄河源区生物结皮对土壤理化属性的影响,主要结论如下:
(1)生物结皮是青藏高原黄河源区植被退化的一个关键环节,其盖度在该区可达37.3%~51.7%,该区生物结皮主要以藻结皮为主,且厚度较干旱荒漠地区(1~5 mm)厚,其厚度平均可达12.6 mm。
(2)该区生物结皮的发育对土壤颗粒组成具有一定的影响,可增加其下覆土层土壤粉粒(0.002~0.050 mm)含量,但粉粒增加主要集中在5~20 cm土层;生物结皮的发育显著降低了结皮层土壤容重,增加了该层土壤田间持水量。
(3)高寒地区藻类型生物结皮均显著增加了结皮层及其下0~20 cm 土层土壤有机质含量;而深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,其下0~20 cm 土层土壤全氮含量有所增加,但差异不显著。研究区生物结皮的形成降低了结皮层土壤pH,对其下0~20 cm 土壤pH没有显著性影响。
猜你喜欢
新疆大学学报(自然科学版)(中英文)(2022年3期)2022-06-04
中老年保健(2021年3期)2021-12-03
小资CHIC!ELEGANCE(2021年25期)2021-07-29
草业学报(2021年6期)2021-06-28
小天使·四年级语数英综合(2021年4期)2021-05-10
阅读与作文(小学高年级版)(2020年6期)2020-07-04
水土保持通报(2020年6期)2020-03-10
新疆大学学报(自然科学版)(中英文)(2018年4期)2018-02-10
延河(2017年7期)2017-07-19
时尚北京(2017年4期)2017-05-02
