
不同种源石斛表型性状多样性及其与地理因子的相关性
2021-05-22 03:38樊小莉黄田钫
植物资源与环境学报 2021年2期
何 涛,樊小莉,鲁 璐,黄田钫
(中国科学院成都生物研究所,四川 成都 610041)
石斛(DendrobiumnobileLindl.)又名金钗石斛,为多年生草本植物,生于林中树干或山谷岩石上;在中国主要分布于四川南部、贵州西南部至北部、云南东南部至西北部、湖北、广西、海南和台湾等地[1]。石斛具有极高的药用、生态和科研价值,目前,有关石斛的研究主要集中于化学成分[2-4]、药理作用[5-6]、分子标记[7-11]、基因克隆[12-13]、组织培养[14]、遗传转化[15-16]以及人工栽培[17-18]等方面。对石斛分子遗传多样性的研究结果[19-23]表明:在分子水平上,不同种源石斛存在高度变异,种源间的变异大于种源内,且种源间存在较高水平的遗传分化。而在形态特征上,不同产地石斛的茎直径存在显著差异[24],表明石斛的遗传和表型均存在不同程度的变异。
表型多样性是遗传多样性的主要研究内容之一,在适应和进化上具有重要意义,有助于了解其遗传稳定性和选择潜力。表型性状具有直观、测量简单、经济方便的特点,可以快捷、有效地检测群体的变异和分化。目前,虽然已有研究者对石斛表型性状进行了初步的观察和统计[24],但不同产地石斛表型性状多样性及其与地理因子的关系尚不清晰,不利于石斛资源的可持续利用和保护。
鉴于此,作者从四川、贵州和云南的不同产地收集了10个石斛种源,对其营养器官和花器官的18个表型性状进行了测定,采用变异系数、Shannon-Wiener多样性指数和表型分化系数以及方差分析、相关性分析和聚类分析等指标和方法,对不同种源间各表型性状的表型分化程度和变异规律及其影响因子进行分析,以期为石斛的生物多样性保护和种质资源的可持续利用提供基础数据。
1 材料和方法
1.1 材料
在实地调查的基础上,在四川、贵州和云南选取10个有代表性的石斛种源地,包括贵州省赤水市的旺隆镇和复兴镇,四川省合江县的五通镇、凤鸣镇和福宝镇,四川省宜宾市的思坡镇和宋家镇,四川省洪雅县柳江镇,四川省夹江县歇马乡以及云南省勐腊县勐腊镇,各种源产地的基本概况见表1。对种源地中心位置进行GPS定位,各种源地间隔大于或等于15 km。在生境为林下岩石上的5个种源地分别设置1个面积约2 000 m2的样地,随机选择30丛石斛样本;在生境为花盆中或树干上的种源地随机选择零散种植的石斛样本4至20丛。
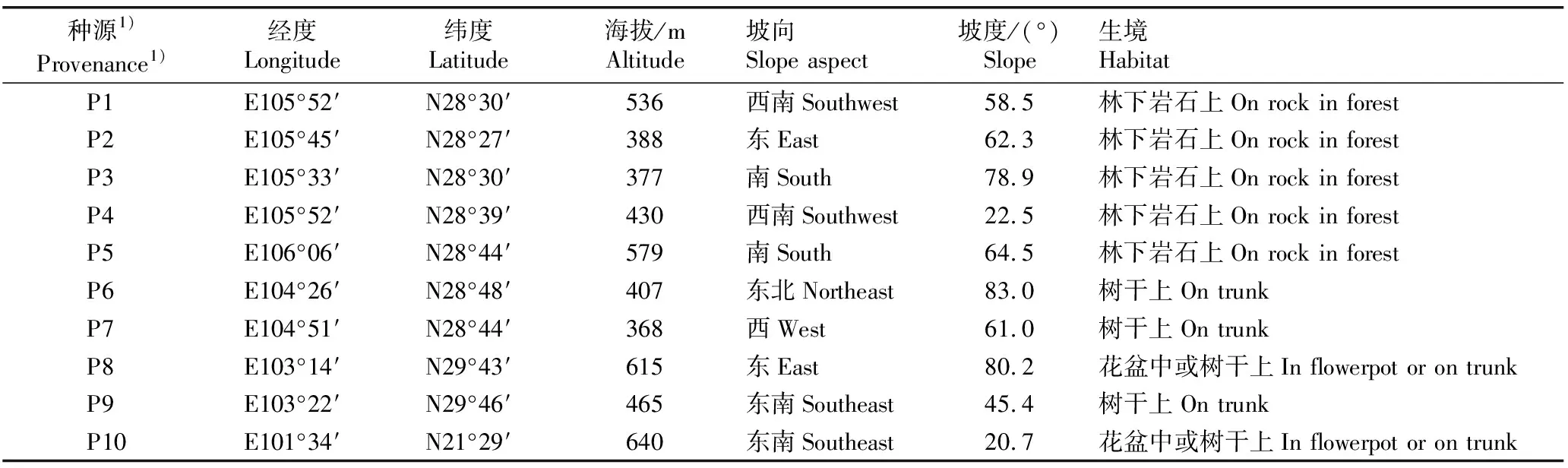
表1 供试10个石斛种源产地的基本概况
供试石斛样株为野生转人工育苗的栽培植株,栽种年限在5 a以上,已稳定生长和繁衍,且无明显病虫害。所有样株由作者进行种类鉴定。在4月至5月的盛花期采集50朵带花梗的完整花以及50条2年生带叶的完整鲜茎作为测定样本。
1.2 方法
1.2.1 表型性状的选择 依据文献[25-27]确定表型性状的选择标准,选取易于获取的18个表型性状,包括8个营养器官性状(茎节数、茎长、茎基部直径、茎中部直径、茎上部直径、叶片长、叶片宽和叶片长宽比)以及10个花器官性状(花梗长、中萼片长、中萼片宽、中萼片长宽比、侧萼片长、侧萼片宽、侧萼片长宽比、花瓣长、花瓣宽和花瓣长宽比)。
1.2.2 测定方法 单茎的茎节数直接目测计数;采用直尺(精度0.1 cm)分别测量茎长(茎基部至茎尖的长度)、叶片长(伸直状态下叶片基部到叶尖的长度)、叶片宽(伸直状态下叶片中部最宽处的宽度)、花梗长(连接花朵和茎的短柄长度)、中萼片长(自然状态下中萼片基部至尖端的长度)、中萼片宽(自然状态下中萼片中部的宽度)、侧萼片长(自然状态下侧萼片基部至尖端的长度)、侧萼片宽(自然状态下侧萼片中部的宽度)、花瓣长(自然状态下花瓣基部至尖端的长度)和花瓣宽(自然状态下花瓣中部的宽度);采用游标卡尺(精度0.01 mm)分别测量茎基部直径(茎基部第1节间的最大直径)、茎中部直径(茎中部节间的最大直径)和茎上部直径(茎最上部节间的最大直径)。根据测量结果,分别计算叶片、中萼片、侧萼片和花瓣的长宽比。
1.3 数据处理和分析
采用EXCEL 2010软件整理数据。采用DPS 18.10软件计算变异系数、Shannon-Wiener多样性指数,并进行Duncan’s多重比较、方差分析、相关性分析和聚类分析(UPGMA法),分析前对数据进行标准化处理和转换。参照文献[28]计算表型分化系数。
2 结果和分析
2.1 不同种源石斛表型性状的比较和变异分析
不同种源石斛营养器官和花器官性状的测定结果及变异系数分别见表2和表3。
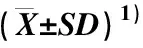
表2 不同种源石斛营养器官和花器官18个表型性状的比较
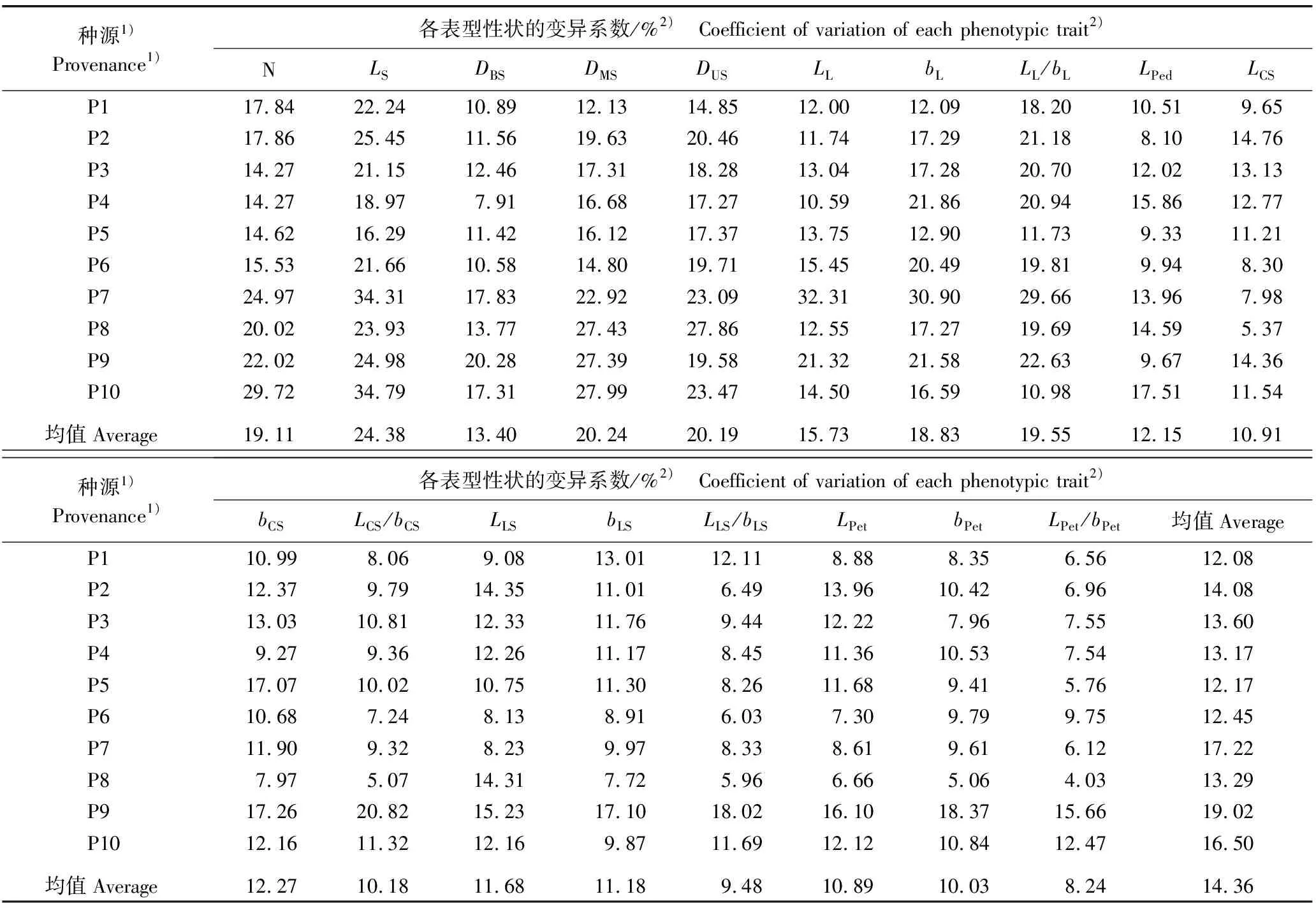
表3 不同种源石斛营养器官和花器官18个表型性状的变异系数
2.1.1 营养器官性状比较 由表2可见:供试10个石斛种源的茎节数、茎长、茎基部直径、茎中部直径、茎上部直径、叶片长、叶片宽和叶片长宽比的均值分别为9.6、30.08 cm、3.67 mm、9.80 mm、5.06 mm、8.98 cm、2.02 cm和4.59。其中,云南省勐腊县勐腊镇(P10)种源的茎节数、茎基部直径、茎中部直径、茎上部直径、叶片长和叶片宽均最大,且总体上与其他种源差异显著(P<0.05);贵州省赤水市旺隆镇(P1)种源的茎长最大,且与其他种源差异显著;四川省宜宾市宋家镇(P7)种源的叶片长宽比最大,但其茎基部直径和茎上部直径均最小,且总体上与其他种源差异显著;四川省合江县福宝镇(P5)种源的叶片长和叶片宽均最小,且与多数种源差异显著;四川省洪雅县柳江镇(P8)种源的茎节数和茎中部直径均最小,且与多数种源差异显著;四川省夹江县歇马乡(P9)种源的茎长和叶片长宽比均最小,且与多数种源差异显著。总体上看,8个营养器官性状在多数种源间存在显著差异。
2.1.2 花器官性状比较 由表2还可见:供试10个石斛种源的花梗长、中萼片长、中萼片宽、中萼片长宽比、侧萼片长、侧萼片宽、侧萼片长宽比、花瓣长、花瓣宽和花瓣长宽比的均值分别为4.58 cm、3.66 cm、1.29 cm、2.87、3.53 cm、1.21 cm、2.93、3.75 cm、2.19 cm和1.71。其中,四川省宜宾市思坡镇(P6)种源的中萼片长、中萼片长宽比、侧萼片长、侧萼片长宽比、花瓣长和花瓣长宽比均最大,且与其他种源差异显著;P10种源的花梗长、中萼片宽、侧萼片宽和花瓣宽均最大,且与多数种源差异显著;而P8种源的花梗长、中萼片长、中萼片宽、侧萼片长、侧萼片宽和花瓣长宽比均最小,且除花瓣长宽比外,其他表型性状总体上与另9个种源差异显著;P9种源的花瓣长和花瓣宽均最小,且总体上与其他种源差异显著;P1种源的中萼片长宽比最小,且与多数种源差异显著;P5种源的侧萼片长宽比最小,且与多数种源差异显著。总体上看,10个花器官性状在多数种源间存在显著差异。
2.1.3 表型性状变异分析 由表3可见:石斛营养器官和花器官性状在种源间和种源内均存在不同程度的变异。在8个营养器官性状中,茎长的变异系数最大,为16.29%~34.79%,均值为24.38%;茎基部直径的变异系数最小,为7.91%~20.28%,均值为13.40%,表明在种源内个体间茎长的差异较大,而茎基部直径的差异较小。在10个花器官性状中,中萼片宽的变异系数最大,为7.97%~17.26%,均值为12.27%;花瓣长宽比的变异系数最小,为4.03%~15.66%,均值为8.24%,说明在种源内个体间中萼片宽的差异较大,而花瓣长宽比的差异较小。总体上看,8个营养器官性状的变异系数均值均大于13%,而10个花器官性状的变异系数均值均小于13%,表明石斛营养器官性状的变异水平高于花器官性状。
在10个种源中,P9种源的变异系数均值最大(19.02%),P1种源的变异系数均值最小(12.08%)。变异系数总均值为14.36%,表明石斛各种源间表型性状的变异较丰富。
2.2 不同种源石斛表型性状的多样性和表型分化分析
不同种源石斛营养器官和花器官性状的Shannon-Wiener多样性指数和表型分化系数分别见表4和表5。
2.2.1 多样性分析 由表4可见:供试10个石斛种源18个表型性状的Shannon-Wiener多样性指数均在3.8以上,均值为3.881~3.908,变幅较小。在18个表型性状中,茎长的Shannon-Wiener多样性指数最小,而花瓣长宽比的Shannon-Wiener多样性指数最大。总体上看,在不同种源中,石斛的18个表型性状均存在丰富的多样性,且多样性水平相近。
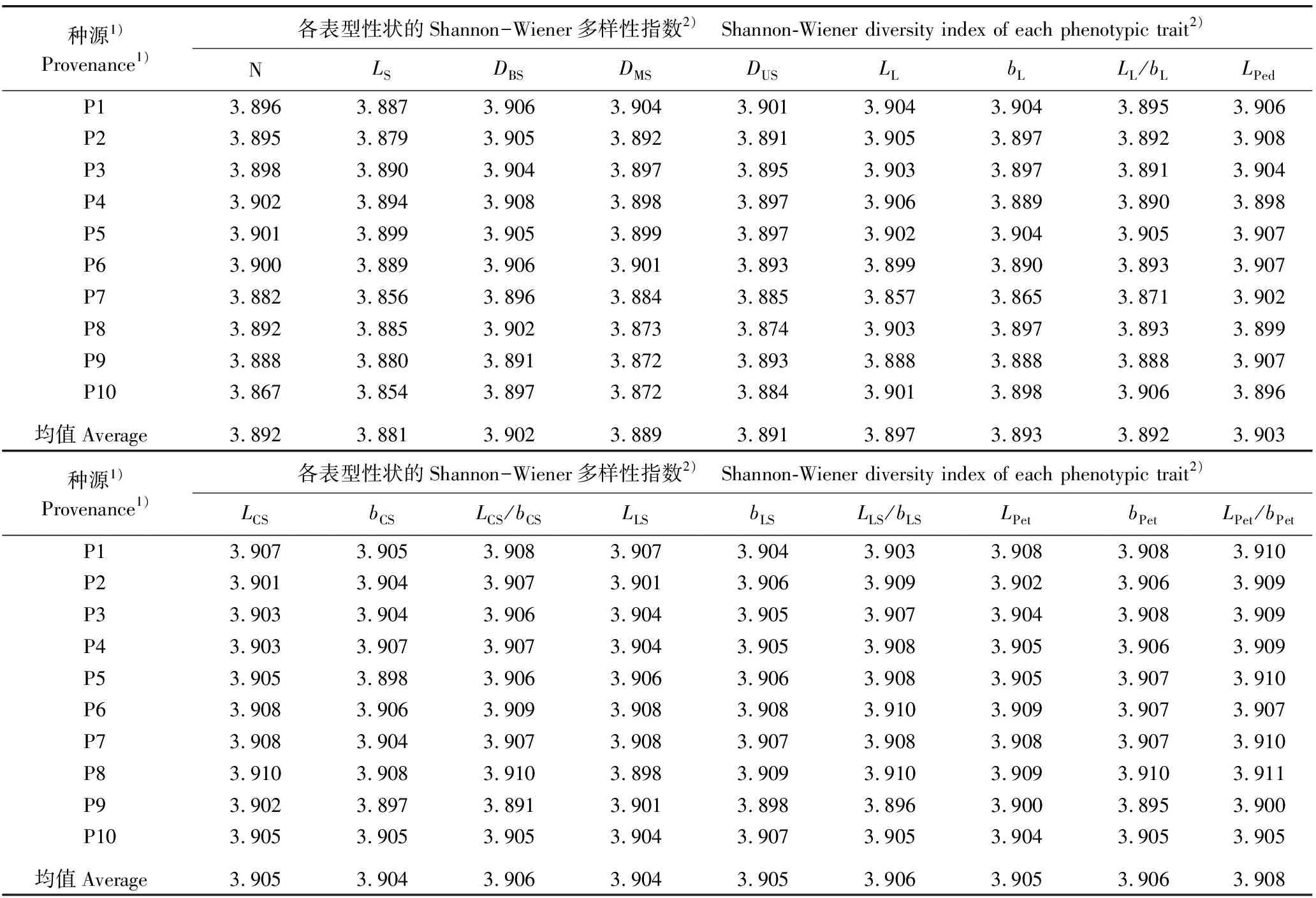
表4 不同种源石斛营养器官和花器官18个表型性状的Shannon-Wiener多样性指数
2.2.2 表型分化分析 由表5可见:石斛18个表型性状在种源间均存在极显著(P<0.01)差异;而在种源内只有茎节数、茎长、花瓣长和花瓣宽4个表型性状存在极显著差异。种源间各表型性状的方差分量均大于或等于种源内,种源间的方差分量为0.000 1~0.036 2,种源内方差分量为0.000 1~0.015 1。
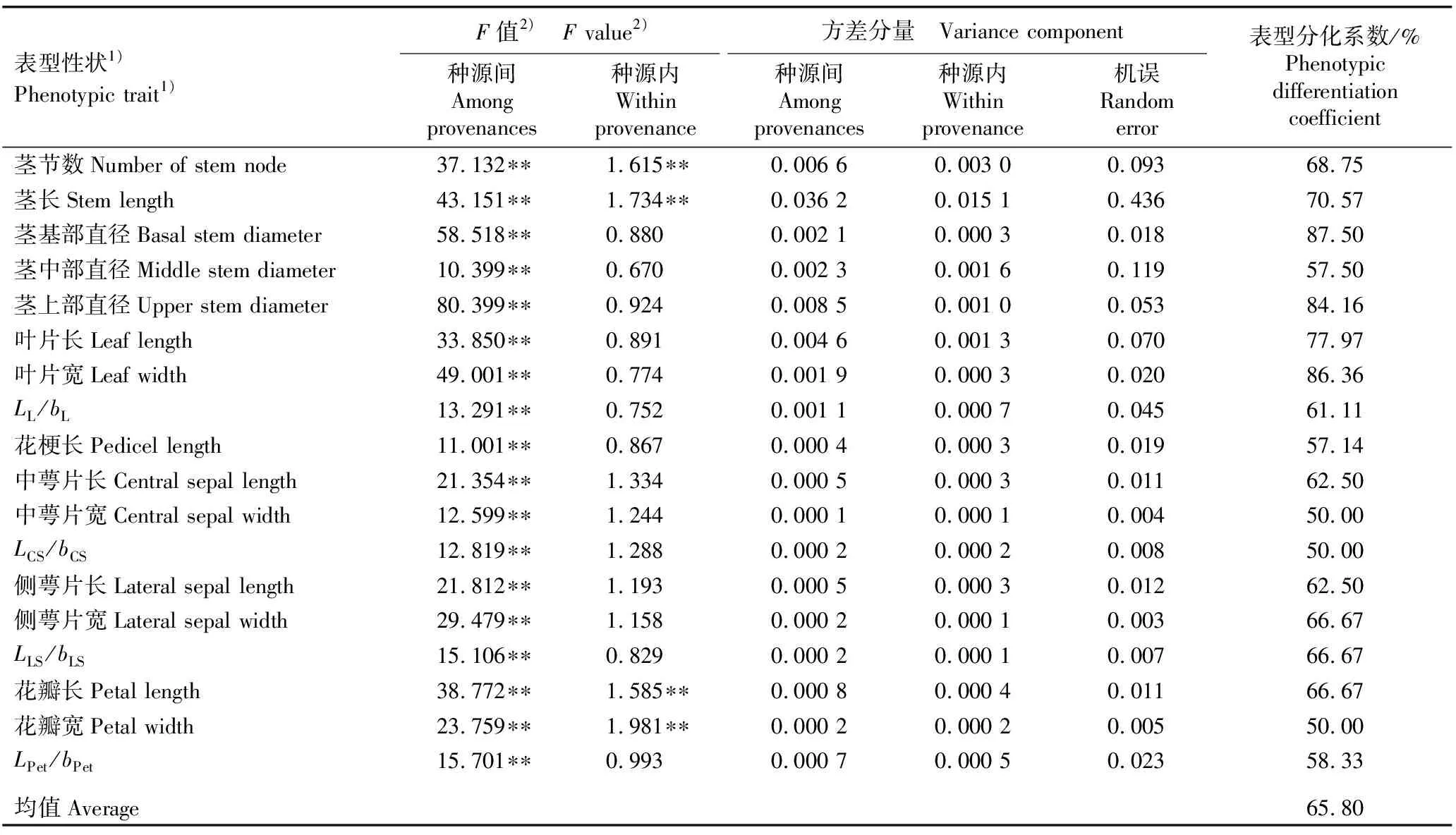
表5 不同种源石斛营养器官和花器官18个表型性状的方差分析结果和表型分化系数
各表型性状的表型分化系数为50.00%~87.50%,其中,仅中萼片宽、中萼片长宽比和花瓣宽3个表型性状的表型分化系数为50.00%,其他表型性状的表型分化系数均在57%以上;18个表型性状的表型分化系数均值为65.80%,表明种源间的变异是石斛表型性状变异的主要来源。
2.3 不同种源石斛表型性状与地理因子的相关性分析
对不同种源石斛的18个表型性状与经度、纬度和海拔的相关性进行分析,结果见表6。
由表6可见:在18个表型性状中,仅叶片长和叶片宽与经度呈显著(P<0.05)负相关,相关系数分别为-0.705和-0.697,其他表型性状与经度均无显著相关性,表明石斛叶片生长呈现以经度为主导的变异模式,即随经度升高,叶片变短、变窄。茎节数、茎基部直径、茎中部直径、茎上部直径和花梗长与纬度呈显著或极显著(P<0.01)负相关,相关系数分别为-0.712、-0.905、-0.786、-0.750和-0.660,其他表型性状与纬度均无显著相关性,表明这5个指标均呈现以纬度为主导的变异模式,即随纬度升高,茎节数减少、茎直径变小、花梗变短。18个表型性状与海拔均无显著相关性,表明海拔变化对石斛表型性状变异无明显的影响效应。
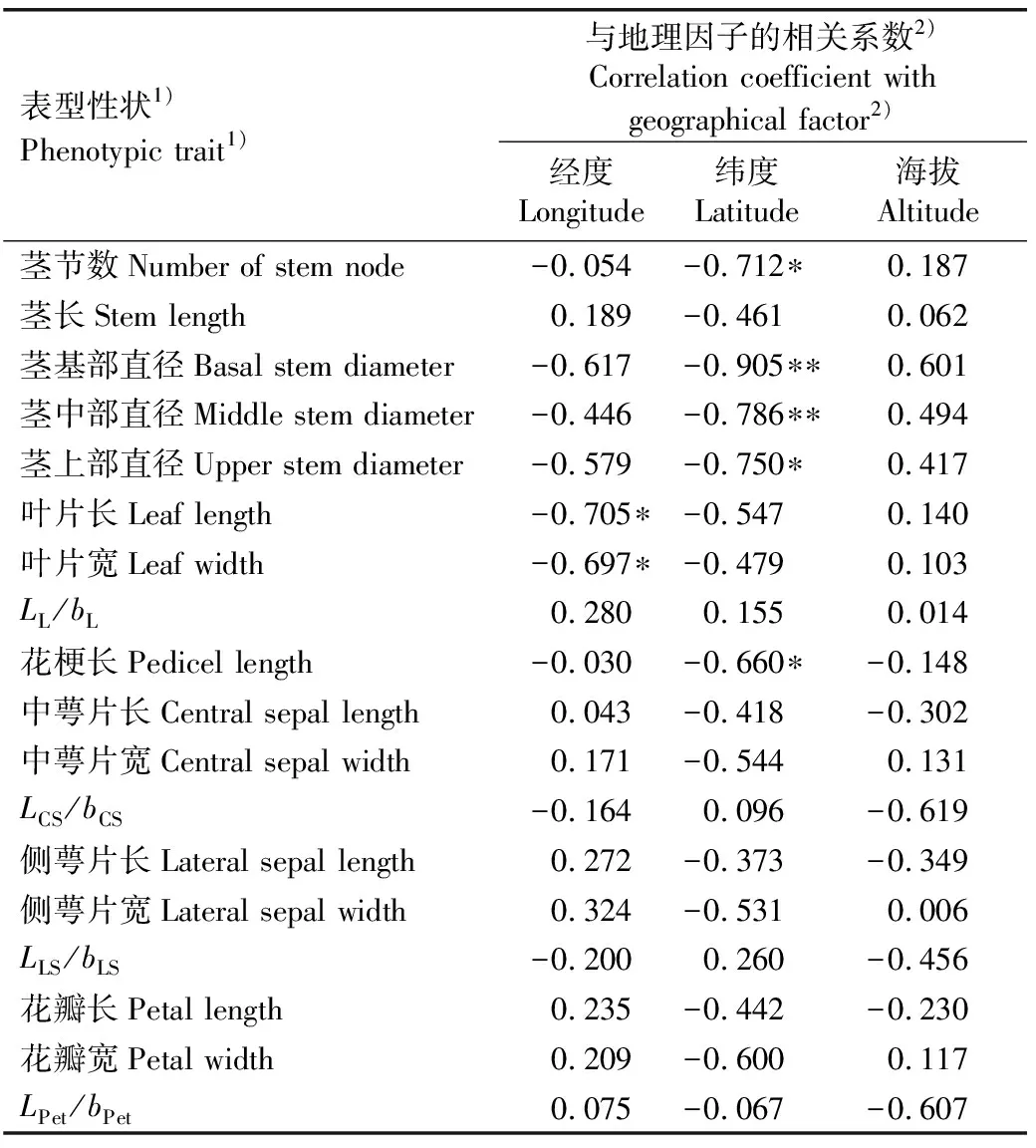
表6 不同种源石斛营养器官和花器官18个表型性状与地理因子的相关系数
2.4 不同种源石斛的聚类分析
基于营养器官和花器官的18个表型性状,采用欧氏距离对不同种源石斛进行聚类分析,结果见图1。
由图1可见:在欧氏距离6.5处,可将10个种源分为3组:组Ⅰ包含贵州省赤水市的旺隆镇和复兴镇,四川省合江县的五通镇、凤鸣镇和福宝镇,四川省宜宾市的宋家镇和思坡镇的7个种源,纬度均在N28°范围内,且这7个种源的地理位置相近,均位于四川南部和与四川南部交界的贵州西北部;组Ⅱ包含四川省洪雅县柳江镇和夹江县歇马乡的2个种源,纬度均在N29°范围内,且这2个种源均位于四川西南部;组Ⅲ仅包含云南省勐腊县勐腊镇的1个种源,纬度在N21°范围内,该种源位于云南最南端,与其他9个种源的地理位置最远。
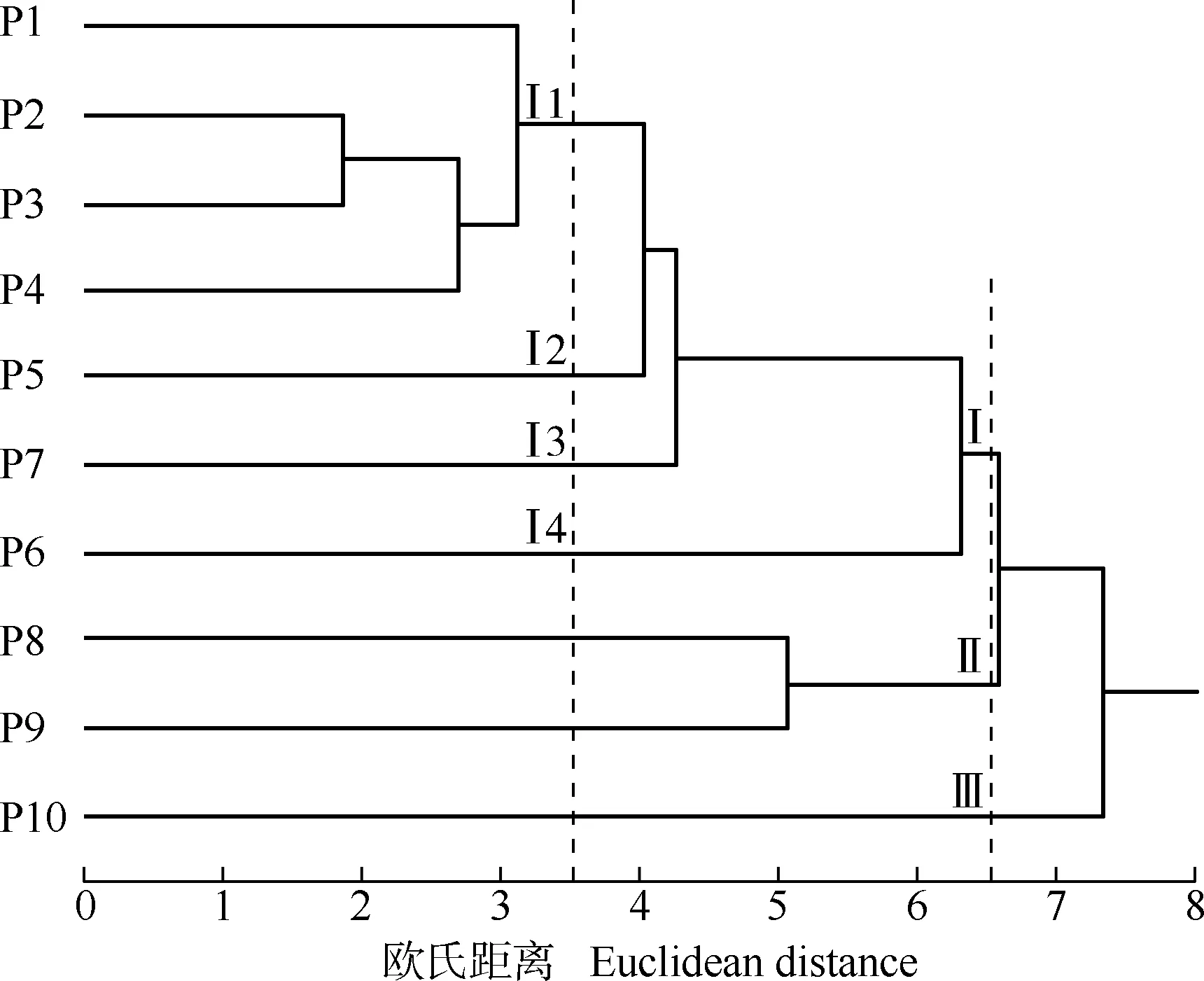
P1,P2:分别来源于贵州省赤水市的旺隆镇和复兴镇Collected from Wanglong Town and Fuxing Town in Chishui City of Guizhou Province,respectively;P3,P4,P5:分别来源于四川省合江县的五通镇、凤鸣镇和福宝镇Collected from Wutong Town,Fengming Town and Fubao Town in Hejiang County of Sichuan Province,respectively;P6,P7:分别来源于四川省宜宾市的思坡镇和宋家镇Collected from Sipo Town and Songjia Town in Yibin City of Sichuan Province,respectively;P8:来源于四川省洪雅县柳江镇Collected from Liujiang Town in Hongya County of Sichuan Province;P9:来源于四川省夹江县歇马乡Collected from Xiema Township in Jiajiang County of Sichuan Province;P10:来源于云南省勐腊县勐腊镇Collected from Mengla Town in Mengla County of Yunnan Province.
在欧氏距离3.5处,组Ⅰ又可分为4个亚组:贵州省赤水市的旺隆镇和复兴镇2个种源以及四川省合江县的五通镇和凤鸣镇的2个种源聚为亚组Ⅰ1,经度均在E105°范围内;四川省合江县福宝镇的1个种源单独组成亚组Ⅰ2,经度在E106°范围内;四川省宜宾市的宋家镇和思坡镇的2个种源各自单独成组,分别为亚组Ⅰ3和Ⅰ4,经度均在E104°范围内。
总体上看,供试10个种源的分组与其地理位置和纬度有关,而亚组内种源的聚类则与经度相关,聚类结果表现出与经度和纬度相关的连续变异模式。
3 讨论和结论
上述研究结果表明:在10个石斛种源的18个表型性状中,有16个表型性状的变异系数均值大于10%,占表型性状总数的88.9%,仅侧萼片长宽比和花瓣长宽比2个表型性状的变异系数均值小于10%,仅占表型性状总数的11.1%;10个种源的变异系数均值为12.08%~19.02%,均大于10%。变异系数越大,表型性状差异越明显,遗传变异可能性越大,表明不同种源石斛的营养器官和花器官存在丰富的变异,可为石斛的遗传改良和生物多样性保护提供物质基础。变异系数的差异还反映了不同表型性状对环境的不同适应能力[27],同一种源内也可能存在多种变异类型[29]。在本研究中,石斛的8个营养器官性状的变异系数均值均大于13%,而10个花器官性状的变异系数均值均小于13%,表明石斛茎和叶性状的变异水平高于花器官性状,花器官受环境的影响较小,更趋于稳定[30]。
10个石斛种源18个表型性状的Shannon-Wiener多样性指数均在3.8以上,说明不同种源石斛的表型性状变异较大,多样性丰富。但Shannon-Wiener多样性指数与变异系数的变化规律并不一致,例如:茎长的变异系数均值最大(24.38%),花瓣长宽比的变异系数均值最小(8.24%),而茎长的Shannon-Wiener多样性指数均值最小(3.881),花瓣长宽比的Shannon-Wiener多样性指数均值却最大(3.908)。虽然变异系数和Shannon-Wiener多样性指数都是反映生物多样性的重要指标,但这2个指标变化规律的不一致揭示了其内涵的不同。表型性状的变异系数反映的是表型性状的离散程度,数值越大,说明表型性状的变异程度越大;Shannon-Wiener多样性指数反映的是表型性状的多样性,数值越大,说明表型性状多样性越丰富。但二者之间不存在关联性[31]。
种源间变异是种内生物多样性的重要组成部分[32-33]。方差分析结果表明:石斛18个表型性状种源间的F值为10.399~80.399,种源间差异均极显著(P<0.01),而种源内的F值仅为0.670~1.981,显示各表型性状在种源间的F值明显大于种源内。石斛所有表型性状的表型分化系数均大于或等于50.00%,表型分化系数均值为65.80%,显示种源间表型性状的分化程度明显大于种源内。表明不同种源石斛的表型性状变异丰富,在种源水平上未表现出表型多样性水平下降,且种源间变异是石斛种源表型性状变异的主要来源。
已有的相关研究结果[33-35]显示:环境条件(如经度、纬度和海拔等主要地理因子)变化是种源间变异形成的主要因子,并因植物种类的不同而异。本研究结果显示:石斛表型性状变异丰富的原因与所选择的种源产地范围较广有关,10个种源的产地分别位于四川、贵州和云南,跨越了3个省份,经纬度的跨度较大(分别达到6°和9°),气候类型变化多样,为了适应不同区域环境的异质性,石斛的表型性状随之发生了变化。
在不同的环境中,石斛通过形态变化和表型性状分化调整自身的生长发育及其与环境之间的关系,从而适应环境的变化。相关性分析结果表明:石斛的茎节数、茎基部直径、茎中部直径、茎上部直径和花梗长与纬度均呈显著(P<0.05)或极显著负相关;叶片长和叶片宽与经度呈显著负相关;其他表型性状与经度和纬度均无显著相关性。表明随纬度的升高,茎节数减少、茎直径变小、花梗变短;随地理位置的西移,叶片变得更为宽大。纬度的变化,意味着气候的变化,纬度越低,气温越高,而较高的气温有利于石斛的营养生长,使植株变得高大。这也说明营养器官性状更容易受到环境因子中水分、养分和光照的影响,如云南省勐腊县勐腊镇种源的产地纬度只有N21°,但该种源石斛的茎叶粗大,这也符合石斛为热带亚热带植物、喜高温高湿环境的特点。此外,石斛的所有表型性状与海拔均无显著相关性,说明海拔对石斛表型性状无明显的影响效应。
石斛为兰科(Orchidaceae)附生植物,可附生在岩石上,也可附生在树干上。贵州省赤水市的旺隆镇和复兴镇以及四川省合江县的五通镇、凤鸣镇和福宝镇的5个种源的石斛均生长在林下岩石上;而其他5个种源的石斛均附生在不同树种的树干上。从表型性状的变异系数看,附生在林下岩石上的石斛种源各表型性状的变异程度总体上小于附生在树干上的种源,表明不同的附生模式也是造成石斛表型性状变异的原因之一。
聚类分析结果表明:在欧氏距离6.5处,可将10个种源分为3组,总体上表现出地理位置和纬度相近的种源聚在一起的规律。组Ⅰ包含的7个种源,纬度均在N28°范围内,均位于四川南部和与四川南部交界的贵州西北部;组Ⅱ包含的2个种源,纬度均在N29°范围内,均位于四川西南部;组Ⅲ仅包含1个种源,纬度在N21°范围内,位于云南最南端,该种源与其他9个种源的距离最远。表明石斛的表型变异与其产地的地理位置有关,表现出按地理位置而聚类的特征[36-38]。在欧氏距离3.5处,可将组Ⅰ再进一步分为4个亚组,亚组内种源聚类主要与经度有关,其中,贵州省赤水市复兴镇和四川省合江县五通镇的2个种源先聚在一起,并与贵州省赤水市旺隆镇和四川省合江县凤鸣镇的2个种源聚在一起,这4个种源的来源地经度均在E106°范围内;而经度在E104°范围内的四川省宜宾市的思坡镇和宋家镇的2个种源各自独立成组。亚组的聚类结果说明在地理因子相近的条件下,遗传因子在表型性状变异中具有主要作用,杜会聪等[39]的研究结果也佐证了这一观点,即在所选研究对象的分布范围较小时,遗传因子是导致表型性状出现差异的主要原因。另外,根据亚组Ⅰ1的聚类结果,推测贵州省赤水市复兴镇与四川省合江县五通镇的种源的原产地可能相同,而与同一个市的旺隆镇的种源的原产地可能不同。
综上所述,不同石斛种源的营养器官和花器官性状的多样性丰富,种源间变异是其表型性状变异的主要来源,不同石斛种源的表型性状呈现以经纬度主导的地理变异模式。在本研究中,并没有对石斛的表型质量性状(植株茎、叶和花的形态和颜色)以及种实性状(种子质量和果实质量等)等进行考察,缺乏表型性状变异与气候因子的相关性分析,且对不同石斛种源的表型性状变异与其遗传变异的一致性也缺乏深度研究,因此,后续将进一步对这些科学问题进行研究和探讨。
猜你喜欢
特产研究(2022年6期)2023-01-17
辽宁林业科技(2021年1期)2021-03-16
广西植物(2020年13期)2020-09-12
广西植物(2020年13期)2020-09-12
武夷学院学报(2017年9期)2017-11-29
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17
江苏农业科学(2017年1期)2017-02-27
广西林业科学(2016年3期)2016-03-16
花卉(2015年11期)2015-09-14
湖北农业科学(2014年16期)2014-10-20
